Педобионты трансформированных экосистем вблизи населенных пунктов Иволгино-Удинской котловины Селенгинского среднегорья

Автор: Эрдынеева Ирина Андреевна, Доржиева Оюна Дымбрыловна, Аюрзанаева Марьяна Васильевна
Журнал: Вестник Бурятского государственного университета. Биология, география @vestnik-bsu-biology-geography
Статья в выпуске: 2, 2017 года.
Бесплатный доступ
В работе рассматривается видовой состав педобионтов трансформированных экосистем вблизи населенных пунктов Иволгино - Удинской котловины Селенгинского среднегорья (в районах с. Нижняя Иволга, Сотниково, Тальцы и 102-го микрорайона. На состав населения педобионтов влияют абиотические и антропогенные факторы, а также рельеф местности, который формируют свою микростацию.
Насекомые, почвенные насекомые, трансформированная экосистема, биоиндикатор, численность
Короткий адрес: https://sciup.org/148317837
IDR: 148317837 | УДК: 595.762
Pedobionts of transformed ecosystems near settlements Ivolgino - Udinsky basin of the Selenga midland
The work deals with the species composition of pedobionts, transformed ecosystems near settlements Ivolgino - Udinsky basin (in areas Nizhnyaya Ivolga, Sotnikovo, Talcy and the 102nd district) of Selenginskyi midland. The composition of pedobionts influence of abiotic and anthropogenic factors, as well as the terrain that form its microstate.
Текст научной статьи Педобионты трансформированных экосистем вблизи населенных пунктов Иволгино-Удинской котловины Селенгинского среднегорья
В работе Р.О. Бутовского отмечено, что для оценки состояния наземных экосистем рекомендуется использовать различные группы беспозвоночных животных, преимущественно обитателей почвы и подстилки. Сообщества почвенных беспозвоночных реагируют на антропогенные воздействия более отчетливо и раньше, чем это можно обнаружить на основе химических анализов почвы. Это объясняется тем, что подавляющая доля поступающих в окружающую среду веществ удерживаются верхними слоями почвы, из которой они вовлекаются в биогенный круговорот [3].
В работе Бабенко А.С.[2] указано, что встречаемость тех или иных видов может быть показателем влажности или сухости местообитания. Однако, характеристики гидротермического режима по встречаемости тех или иных видов имеют относительный, географически обусловленный характер, и популяции одного вида в разных частях широкого ареала характеризуются часто разным диапазоном требований в отношении гидротермических условий (согласно правилу сме- ны стаций). Представители почвенной фауны могут быть индикаторами и солевого режима почв. Например, в солончаках наблюдается специфический состав беспозвоночных-галофилов (стафилиниды рода Bledius, чернотелки), которые, как и растения — галофиты, могут быть индикаторами процессов засоления. Некоторые беспозвоночные, являющиеся в большей или меньшей степени калько-филами (моллюски, диплоподы, мокрицы), могут быть индикаторами богатства почвы кальцием. Наиболее удобны для биоиндикационных работ крупные почвенные животные (представители мезо- и макрофауны). Среди них достаточно много истинных педобионтов, что обусловливает тесную и ярко выраженную связь этих животных с изменениями, происходящими в почве. Важно, что среди представителей мезофауны много видов-полифагов, мало связанных с определенным типом пищи.
В целом, выбор определенной группы или вида беспозвоночных в качестве индикатора почвенных условий должен основываться на его доминировании в естественных (или эталонных) местообитаниях.
Материалы и методы исследования
Исследования проводились в трансформированной экосистеме вблизи населенных пунктов Нижняя Иволга, Сотниково, Тальцы и 102 — микрорайон Ивол-гино — Удинской котловины Селенгинского среднегорья.
Материал данной работы собран с июня по август 2013 -2014 гг.. Для сбора материала использован стандартный метод послойной выборки почвенных проб размером 50x50 (0,25 м²) до глубины 20 см [6,7]. В результате собраны 127 видов беспозвоночных из 8 отрядов, 19 семейств, 32 родов.
Результаты исследования
Трансформированная экосистема вблизи населенного пункта Нижняя Иволга в период исследования представлена 10 видами (таб.1).
Таблица 1
Видовой состав педобионтов трансформированных экосистем вблизи населенного пункта Нижняя-Иволга Иволгино-Удинской котловины
|
Вид |
Коэфф. встречаемости (%) |
Коэфф. доминирования (%) |
|
|
1. |
Губоногие многоножки ( Chilopoda ) |
0,17 |
17 |
|
2. |
Личинка корнееда ( Eodorcardion sp. ) |
0,11 |
11 |
|
3. |
Эноплопс сибирский ( Enoplops sibiricus ) |
0,05 |
5 |
|
4. |
Хрущ собачий ( Lasiopsis caninus Zoubk .) |
0,17 |
17 |
|
5. |
Личинка чернотелки ( Blaps sp.) |
0,05 |
5 |
|
6. |
Личинка щелкуна ( Corymbites sp.) |
0,11 |
11 |
|
7. |
Личинка комара ( Rhymosia sp. ) |
0,05 |
5 |
|
8. |
Жужелица ( Harpalius sp.) |
0,05 |
5 |
|
9. |
Личинка Пилюльщика ( Byrrhus sp.) |
0,05 |
5 |
|
10. |
Личинка коконопряда ( Gastropacha sp. ) |
0,17 |
17 |
По-видимому, на такое небольшое разнообразие влияют уплотнение почвы в связи выпасом скота и близость автотрассы, а увеличение численности многоножек ( Chilopoda ) связано с ростом степени урбанизации [4].
Видовой состав в трансформированной экосистеме вблизи поселка Сотниково представлен 22 видами (табл. 2).
Таблица 2
Видовой состав педобионтов трансформированных экосистем вблизи населенного пункта Сотниково Иволгино-Удинской котловины
|
Вид |
Коэфф. встречаемости (%) |
Коэфф. доминирования (%) |
|
|
1. |
Клоп — солдатик ( Pyrrhocoris apterus Linnaeus) |
0,06 |
6 |
|
2. |
Личинка скакуна ( Cicindela sp. ) |
0,02 |
2 |
|
3. |
Жужелица (Amara amplicolis Baliani) |
0,06 |
6 |
|
4. |
Жужелица (Amara aenea De Geer) |
0,07 |
7 |
|
5. |
Личинка жужелицы ( Amara sp .) |
0,03 |
3 |
|
6. |
Жужелица (Amara sp.) |
0,02 |
2 |
|
7. |
Личинка жужелица золотистоямчатая ( Carabus clathratus L.) |
0,07 |
7 |
|
8. |
Жужелица (Harpalus lumbaris Mannerhein) |
0,02 |
2 |
|
9. |
Личинка жужелицы ( Harpalius lumbaris Man-nerhein) |
0,02 |
2 |
|
10. |
Жужелица (Harpalius affinis Schrank) |
0,02 |
2 |
|
11. |
Личинка жужелицы ( Harpalius affinis Schrank) |
0,13 |
13 |
|
12. |
Жужелица Curtonotus fodinae ( Mannerheim ) |
0,02 |
2 |
|
13. |
Кожеед сибирский ( Dermestes sibiricus Er.) |
0,06 |
6 |
|
14. |
Чернотелка Anatolica sp. |
0,09 |
9 |
|
15. |
Личинка чернотелки ( Anatolica sp.) |
0,02 |
2 |
|
16. |
Личинка чернотелки ( Crypticus sp.) |
0,02 |
2 |
|
17. |
Личинка чернотелки ( Prosodes ps .) |
0,06 |
6 |
|
18. |
Личинка слоника скосаря ( Otiorhynchus sp. ) |
0,02 |
2 |
|
19. |
Личинка комара ( Rhimozia sp.) |
0,09 |
9 |
|
20. |
Личинка стафилина Кведиус ( Quedius sp. ) |
0,02 |
2 |
|
21. |
Личинка мухи ( Coenomyia sp.) |
0,02 |
2 |
Такое разнообразие связано с тем, что на этих участках в данный период приходится пик активности жизненного цикла некоторых видов насекомых.
На пробной площадке вблизи 102 — микрорайона максимальный коэффициент доминирования составляет от 8–15%, (табл. 4).
Таблица 4
Видовой состав педобионтов трансформированных экосистем вблизи населенного пункта 102-й микрорайон г. Улан-Удэ Иволгино-Удинской котловины
|
Виды |
Доля вида (%) |
Проценты вида (%) |
|
|
1. |
Круглые черви ( Enhifridae ) |
0,08 |
8 |
|
2. |
Многоножка Костянка обыкновенная ( Linthobius forticatus Linnaeus) |
0,02 |
2 |
|
3. |
Губоногие многоножки ( Chilopoda sp. ) |
0,15 |
15 |
|
4. |
Клоп-солдатик ( Pyrrhocoris apterus Linnaeus) |
0,02 |
2 |
|
5. |
Личинка Стафилины ( Philothus sp. ) |
0,02 |
2 |
|
6. |
Личинка жужелицы ( Carabus arvensis Herbst) |
0,06 |
6 |
|
7. |
Личинка Жужелица ( Carabus sp.) |
0,06 |
6 |
|
8. |
Жужелица (Poecilius gebleri Dejean) |
0,02 |
2 |
|
9. |
Личинка Платиума ( Pterosticus sp. ) |
0,02 |
2 |
|
10. |
Личинка Тускляка ( Amara sp.1 ) |
0,11 |
8 |
|
11. |
Личинка Тускляка ( Amara sp.) |
0,02 |
2 |
|
12. |
Личинка Бегун золотистый (Harpalius affinis Schrank) |
0,04 |
4 |
|
13. |
Личинка Бегун 1 ( Harpalius sp .1) |
0,04 |
4 |
|
14. |
Личинка Бегун 2 ( Harpalius sp2 ) |
0,02 |
2 |
|
15. |
Личинка Бегун (Harpalius sp.3) |
0,02 |
2 |
|
16. |
Личинка бронзовки золотистой (Cetonia aurata Linnaeus) |
0,08 |
8 |
|
17. |
Личинка бронзовки медная ( Protaetia metallica Hbst) |
0,06 |
6 |
|
18. |
Личинка хруща ( Melolontha sp). |
0,02 |
2 |
|
19. |
Личинка щелкун блестящий (Selatosomus aeneus Linnaeus) |
0,02 |
2 |
|
20. |
Личинка медляк малый (Gonocephalum pusillum Fabr.) |
0,04 |
4 |
|
21. |
Личинка цикадки ( Draeculacephala sp .) |
0,02 |
2 |
|
22. |
Личинка комар — долгоножка ( Tipula sp. ) |
0,02 |
2 |
Таким образом, в лесных трансформированных экосистемах 102-го микрорайона видовой состав педобионтов представлен 22 видами. Доминируют губо-ногие многоножки ( Chilopoda ) — 15%, личинки Тускляка ( Amara sp.1 ), личинки бронзовки (Cetonia) и круглые черви (Enhitridae ) — 8%, остальные виды представлены от 6 до 2%. Такое многообразие видового состава педобионтов в почве связано с рельефом местности, так как в отличие от станции Тальцы здесь имеется мезорельеф, который формируют свою микростацию.
В трансформированных лесных участках вблизи станции Тальцы максимальный коэффициент встречаемости беспозвоночных животных составляет 0,5– 0,14%, а доминирование от 50–14% (табл. 3).
Таблица 3
Видовой состав педобионтов трансформированных экосистем вблизи населенного пункта Тальцы Иволгино-Удинской котловины
|
Вид |
Коэфф. встречаемости (%) |
Коэфф. доминирования (%) |
|
|
1. |
Дождевые черви |
0,5 |
50 |
|
2. |
Личинка камора ( Tipula sp.) |
0,09 |
9 |
|
3. |
Poecilius gebleri |
0,09 |
9 |
|
4. |
Жужелица ( Carabus sp.) |
0,14 |
14 |
|
5. |
Личинка щелкуна (Salatosomus sp. ) |
0,09 |
9 |
|
6. |
Личинка слоника скосаря ( Otiorhynchus sp. ) |
0,09 |
9 |
В лесных трансформированных экосистемах вблизи станции Тальцы основную долю составляют круглые черви ( Enhitridae ) — 50%, жужелицы ( Carabus sp .) — 14%, личинки щелкуна (Salatosomus sp. ), комара ( Tipulа ), слоника скосаря ( Otiorhynchus sp. ) составляют 9%. Такое низкое многообразие, возможно, связано с тем, что в почвах имеется высокое содержание фульвокислот. Фульвокислоты обладают резкокислой реакцией и хорошей растворимостью в воде, поэтому они энергично разрушают минеральную часть почвы, играя существенную роль в развитии подзолообразовательных процессов. При взаимодействии с катионами щелочных и щелочноземельных металлов фульвокислоты образуют соли, растворимые в воде при любой реакции раствора и способны вымываться нисходящими токами воды из верхней части почвы вниз. [5]. И такая среда влияет на видовой состав педобионтов.
Таким образом, по индексу Чекановского — Серенсена трансформированные лесные экосистемы станции Тальцы и 102-го микрорайона отличаются от других точек исследования и сходны между собой на 81%, так как представляют лесные экосистемы. Станция Тальцы расположена на северо-восточной стороне от г. Улан-Удэ, поэтому, возможно, на состав населения педобионтов влияют абиотические и антропогенные факторы. Трансформированные экосистемы вблизи населенных пунктов Sotnicovo и Nichnia Ivolga представляют 53% сходства, так как они являются степными трансформированными экосистемами, которые формируют свою стацию, что наглядно демонстрирует дендограмма, где группируются два кластера (102 — cvartal и Taltsy) и ( Sotnicovo и Nichnia Ivolga) (рис. 2).
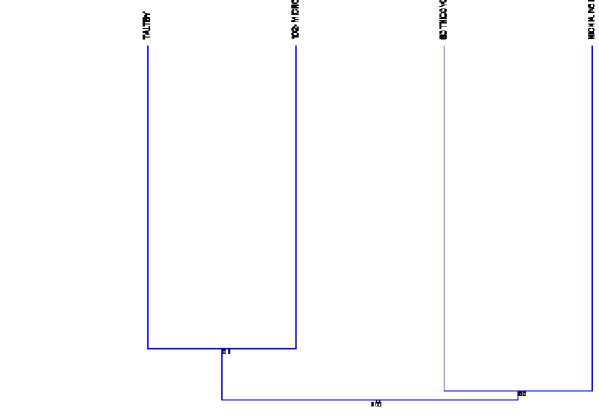
Рис .2. Дендрограмма кластерного анализа четырех объектов, построенная по индексу Чекановского — Серенсена.
Список литературы Педобионты трансформированных экосистем вблизи населенных пунктов Иволгино-Удинской котловины Селенгинского среднегорья
- Алексеева Е. Е. Почвенная мезофауна степей и лесов Западного Забайкалья. М.: Изд-во Моск. гос. пед. ун-та, 1974. 20 с.
- Бабенко А. C. Почвенные беспозвоночные как индикаторы состояния территории // Геохимия живого вещества: материалы Международной молодежной школы- семинара (Томск, 2-5 июня 2013 г.); Томский политехнический университет. Томск: Изд- во Томск. политехн. ун-та, 2013. C. 40-42.
- Бутовский Р. О. Экотоксикология почвенных беспозвоночных животных. Тула: Изд-во Тульск. пед. ун-та, 2009. 80 с.
- Вершинина С. Д. Структура почвенной мезофауны в градиенте урбанизации // Вестник Удмуртского университета. Ижевск, 2011. Вып. 2. С. 84-895.
- Гаркуша И. Ф. Почвоведение с основами геологии. М.; Л.: Сельхозиздат, 1963.
