Перспективный фотосенсибилизатор для антимикробной фотодинамической терапии

Автор: Страховская Марина Глебовна, Беленикина Наталья Серафимовна, Никитина Валентина Васильевна, Коваленко Светлана Юрьевна, Коваленко Илья Борисович, Аверьянов Александр Вячеславович, Рубин Андрей Борисович, Галочкина Татьяна Владимировна
Журнал: Клиническая практика @clinpractice
Рубрика: Оригинальные исследования
Статья в выпуске: 1 (13), 2013 года.
Бесплатный доступ
Показана высокая антимикробная активность и широкий спектр действия фотосенсибилизатора из класса поликатионных металлофталоцианинов. Установлено, что эффективная фотодинамическая инактивация бактерий в присутствии фотосенсибилизатора поликатионной природы определяется его электростатическим связыванием с отрицательно заряженными структурами клеточных стенок бактерий. Полученные результаты обсуждаются с точки зрения перспектив применения антимикробной фотодинамической терапии в медицинской практике.
Фотосенсибилизатор, активные формы кислорода, антимикробная фотодинамическая терапия
Короткий адрес: https://sciup.org/14338444
IDR: 14338444
Promising photosensitizer for antimicrobial photodynamic therapy
A photosensitizer from polycationic metallophthalocyanines class is shown to have a high antimicrobial activity and a wide antimicrobial spectrum. It is established that the effective pho- todynamic inactivation of bacteria in the presence of photosensitizer of polycationic nature is determined by its electrostatic binding to negatively charged structures of bacterial cell walls. The usage of antimicrobial photodynamic therapy in medical practice is discussed from the point of view of the obtained results.
Текст научной статьи Перспективный фотосенсибилизатор для антимикробной фотодинамической терапии
Фотодинамическая инактивация микроорганизмов, в основе которой лежат цитотоксические свойства активных форм кислорода (АФК), генерируемых красителями-фотосенсибилизаторами в фотовозбужденном состоянии, открыта более ста лет назад. Однако лишь в последние два десятилетия исследования в этой области получили активное развитие, что обусловлено ростом антибиотикорезистент-ности и необходимостью разработки альтерна- тивных способов борьбы с возбудителями инфекционных заболеваний. В отличие от антибиотиков, каждый из которых специфически воздействует на определенную мишень в микробной клетке: клеточную стенку, цитоплазматическую мембрану, репликацию ДНК, транскрипцию или трансляцию белков, АФК вызывают неспецифическое повреждение всех клеточных компонентов, потенциально подверженных окислительным реакциям [1]. Множе- ственный характер окислительной деструкции микробных клеток-мишеней затрудняет выработку устойчивости к последующим циклам фотодинамических воздействий.
Фундаментальное различие в чувствительности грамположительных и грамотрицатель-ных бактерий к фотосенсибилизации связывают с принципиальными различиями в строении клеточных стенок этих двух групп бактерий [2]. Общепринято, что важнейшей структурой, отвечающей за общую устойчивость грамотрица-тельных бактерий к различным внешним агентам (антибиотикам, детергентам, красителям), является наружная мембрана, входящая в состав клеточной стенки. В основе наружной мембраны лежит двухслойная асимметричная структура, внешний слой которой состоит преимущественно из липополисахаридов (ЛПС, 2-3,5 х 10 6 молекул/клетку), которые занимают около 75% площади поверхности клетки, и встроенных между ЛПС белковых комплексов [3]. Суммарный отрицательный заряд ЛПС связан с высоким содержанием отрицательно заряженных групп в центральной части этих макромолекул – остатками фосфорной кислоты в D-глюкозамине липида А и/или гептозах кора, карбоксильными группами остатков 3-де-зокси-D-манно-октулозоновой кислоты (КДО) и кислых сахаров кора (галактозы, глюкуроновой кислоты). У грамотрицательных бактерий существуют пути модификации ЛПС, приводящие к уменьшению плотности отрицательных зарядов на поверхности клеток [4]: присоединение положительно заряженных групп (фосфо-этаноламина, аминоарабинозы, глюкозамина) к липиду А и гептозам кора, а также дефосфорилирование этих компонентов ЛПС.
Повышение чувствительности грамотрица-тельных бактерий к фотосенсибилизации имеет место при их дополнительной обработке заряженными поликатионными соединениями [5]. Под воздействием поликатионов происходит высвобождение ЛПС, дезинтеграция наружной мембраны и увеличение ее проницаемости для красителей. Поликатион может быть также использован как наноноситель антибактериальных препаратов, повышающий избирательность их действия за счет электростатического взаимодействия с клеточной стенкой бактерий. Подобные подходы лежат в основе сенсибилизации бактерий с помощью полимиксина, а также использования конъюгатов анионных красителей с полилизином для повышения их фотобактерицидной активности. В то же время и сами молекулы красителей одновременно с ядром – генератором активных форм кислорода, могут нести положительно заряженные заместители, повышающие эффективность взаимодействия фотосенсибилизатора с бактериальной клеткой-мишенью. К таким фотосенсибилизаторам относятся поликатионные фталоцианины, с высокой эффективностью генерирующие синглетный кислород и показавшие высокую эффективность на бактериальной биолюминесцентной тест-системе [6, 7]. В настоящей работе мы приводим данные по антимикробной активности одного из наиболее перспективных соединений – октакатионного фталоцианина цинка.
Материалы и методы исследования
Объектами исследования служили: эталонные штаммы грамположительных бактерий Staphylococcus aureus ATCC 25923 и Streptococcus pyogenes 151 БГСА, а также грамот-рицательных бактерий Escherichia coli ATCC 25922, Pseudomonas aeruginosa ATCC 27853, Campylobacter jejuni NCTC 11635, Helicobacter pylori NCTC 11639, Salmonella enteritidis 1742; клинические изоляты бактерий, среди которых имелись возбудители инфекций кожи и слизистых, и раневых инфекций ( S. aureus, S. pyogenes, E. coli, P. aeruginosa ); виды, вызывающие поражения слизистых желудочно-кишечного тракта ( C. jejuni, H. pylori, S. enteritidis, E. сoli, Serratia marcescens, Proteus mirabilis ). Для культивирования бактерий применяли соответствующие среды: Columbia Agar с добавлением 5% (об/об) или 10% (об/об) бараньей крови, Columbia Agar с добавлением 7% (об/об) лизированной лошадиной крови и Trypcase Soya Agar ("bioMerieux", Франция). Бактерии выращивали в течение суток в термостате при 37°С. Микроаэрофильные штаммы C. jejuni и H. pylori выращивали в анаэ-ростатах в среде с 5% кислорода в течение 4-х суток при 37°С. Материал агаровых культур суспендировали в растворе для инфузий, содержащем 0,9% хлорида натрия, до показателя мутности 1,0 McF с использованием денситометра "Densimat" ("bioMerieux", Франция).
Облучение проводили с использованием источника холодного белого света ЭКОМП (50 мВт/см2) или светодиодного источника красного света с максимумом испускания 684 нм и интенсивностью излучения на уровне образца 20 мВт/см2.
Для определения количества красителя окта-кис(холинил)-фталоцианина цинка, связавшегося в процессе инкубации с бактериальными клетками, раствор красителя в 0,9% NaCl или суспензию бактерий (108 КОЕ/мл) в 0,9% NaCl с той же концентрацией красителя инкубировали при комнатной температуре и пропускали через фильтры с диаметром пор 0,2 мкм ("Sarstedt", Австрия). Количество связавшегося красителя рассчитывали по разнице поглощения фильтратов, учитывая, что для используемого красителя молярный коэффициент поглощения в максимуме ε 683=190000 М-1 см-1.
Поверхностный дзета-потенциал бактериальных клеток измеряли на приборе Zetasizer Nano Z (Malvern Instruments Ltd).
Каждый эксперимент проводили не менее трех-пяти раз. Данные представлены в виде средних значений исследуемых величин ± стандартное отклонение.
В работе использовали реактивы фирмы Sigma (Сент-Луис, США), а также фотосенсибилизатор октакис(холинил)-фталоцианин цинка, синтезированный в ФГУП ГНЦ «НИО-ПИК».
Результаты и обсуждение
При исследовании спектра антимикробной активности октакис(холинил)-фталоцианина цинка были определены минимальные подавляющие рост микроорганизмов концентрации препарата (МПК90) эталонных и клинических штаммов грамположительных и грамотрица-тельных бактерий. МПК90 для клинических видов грамположительных бактерий составляла 0,25-0,30 мкг/мл, а для эталонных штаммов – менее 0,25 мкг/мл при дозе облучения красным светом 5 Дж/см2 (табл.).
Октакис(холинил)-фталоцианин цинк проявлял высокую фотодинамическую активность также в отношении грамотрицательных бактерий, хотя и меньшую по сравнению с грампо-ложительными штаммами. МПК90 в зависимости от вида грамотрицательных бактерий колебалась в достаточно широких пределах – 0,4-2,5 мкг/мл (табл.). Наиболее низкая активность фотосенсибилизатора выявлена в отношении Klebsiella spp. и Proteus mirabilis (МПК90 2,1-2,5 мкг/мл).
Таким образом, антимикробная фотодинамическая активность октакис-(холинил)фта-лоцианина цинка проявляется в отношении различных видов бактерий, в том числе клини- ческих штаммов грамотрицательных бактерий и MRSA штамма золотистого стафилококка. Полученные данные позволяют считать, что данный фотосенсибилизатор имеет широкий спектр антимикробной активности и перспективен для АФДТ различных бактериальных заражений. Однако при этом обнаружена значительная гетерогенность бактериальных штаммов в отношении чувствительности к фотодинамической инактивации, которая наблюдалась как между штаммами одного вида (эталонный штамм АТСС 25922 и клинический изолят E. coli), так и между различными видами (табл., рис. 1).
Известно, что среди различных штаммов бактерий имеется значительная гетерогенность строения олигосахаридов липида А и
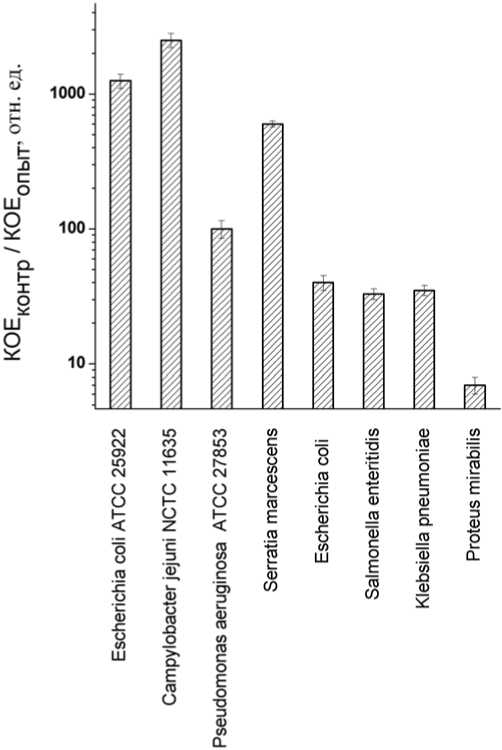
Рис. 1. Эффективность фотодинамической инактивации по тесту уменьшения колониеобразующей способности (КОЕконтр/КОЕопыт) различных штаммов бактерий после 10 мин инкубации суспензий с исходной плотностью108 КОЕ/мл в 0,9% NaCl с 1 мкМ октакис(холинил)-фталоцианина цинка и облучения белым светом в дозе 9 Дж/см2.
Таблица
Активность in vitro октакис(холинил)-фталоцианина цинка
(минимальная подавляющая рост концентрация МПК90, мкг/мл) в отношении различных штаммов бактерий при облучении дозой красного света 684 нм 5 Дж/см2
Повышенное сродство октакатионного фталоцианинового фотосенсибилизатора к клеткам S. marcescens является, очевидно, следстви-
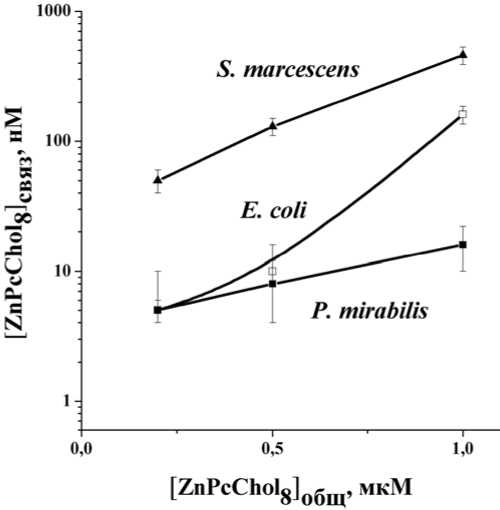
Рис. 2. Связывание октакис(холинил)-фталоцианина цинка клетками различных видов бактерий (108 КОЕ/мл в 0,9% NaCl). Количество связанного красителя [ZnPcChol8]связ рассчитано по разнице поглощения красителя (ε683 = 190000 М-1см-1) в фильтратах его бесклеточных растворов – [ZnPcChol8]общ и клеточных суспензий после 10 мин инкубации – [ZnPcChol8]своб ем отсутствия положительно заряженных заместителей в ЛПС этого вида. Напротив, показано, что липид А ЛПС S. marcescens содержит два глюкозаминовых остатка, каждый из которых фосфорилирован, т.е. несет отрицательный заряд.
Таким образом, катионные фотосенсибилизаторы более эффективно взаимодействуют с бактериальными клетками, ЛПС которых несут большее количество отрицательно заряженных групп. Из рис. 3 видно, что при связывании октакатионного фталоцианина с клетками E.coli происходит нейтрализация отрицательного заряда клеток. Таким образом, механизмом, обеспечивающим связывание катионных фотосенсибилизаторов микробными клетками, является электростатическое взаимодействие положительно заряженных заместителей в молекулах фотосенсибилизатора с отрицательно заряженными центрами связывания на клеточных стенках.
Заключение
Ключевыми параметрами, определяющими, какие из клеточных компонентов или метаболических процессов будут повреждены или инактивированы в процессе АФДТ, а также общую эффективность поражения бактериальных клеток, являются сродство фотосенсибилизатора к клеткам-мишеням и кинетика его захвата с прокрашиванием клеточного объема. Это, в свою очередь, зависит от заряда молекул фотосенсибилизатора. Условием эффективной прямой фотодинамической инактивации является тесная ассоциация фотосенсибилизатора с биологической мишенью, что следует из малого (10-50 нм) диффузионного радиуса синглетного кислорода в биологической среде [1, 5]. Цитоплазматическая мембрана бактерий отделена от внешней среды клеточной стенкой толщиной от 10 до 80 нм в зависимости от видовой принадлежности. Отрицательный заряд клеточных стенок бактерий обусловливает электростатическое связывание с ними катионных соединений. Фотобактерицидная активность октакис(холинил)-фталоцианина цинка определяется наличием восьми катионных заместителей на периферии молекулы.
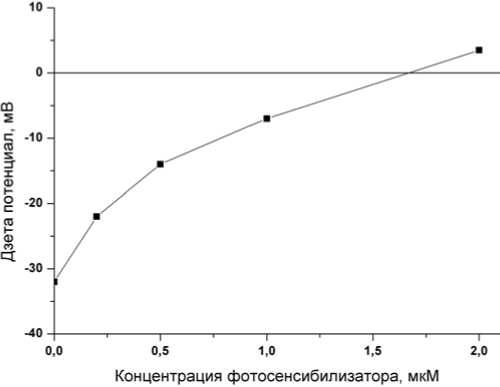
Рис. 3. Нейтрализация поверхностного заряда (дзета-потенциала) клеток Escherichia coli возрастающими концентрациями фотосенсибилизатора октакис(холинил)-фталоцианина цинка.
В заключение отметим, что фотодинамическая инактивация микроорганизмов может являться перспективным методом терапии инфекций, вызванных полирезистентными бактериальными штаммами, а также применяться в целях фотообеззараживания различных предметов и сред медицинского назначения.
Список литературы Перспективный фотосенсибилизатор для антимикробной фотодинамической терапии
- Jori G., Brown S.B. Photosensitized inactivation of microorganisms. Photochem Photobiol Sci 2004; 3: 403-405.
- Nitzan Y., Gutterman M., Malik Z., et al. Inactivation of gram-negative bacteria by photosensitized porphyrins. Photochem Photobiol 1992; 55: 89-96.
- Raetz C.R., Ulevitch R.J., Wright S.D., et al. Gram-negative endotoxin: an extraordinary lipid with profound effects on eukaryotic signal transduction. FASEB J 1991; 5: 2652-60.
- Raetz C.R., Reynolds C.M., Trent M.S. et al. Lipid A modification systems in gram-negative bacteria. Annu Rev Biochem 2007; 76: 295-329.
- Hamblin M.R., Hasan T. Photodynamic therapy: a new antimicrobial approach to infectious disease? Photochem Photobiol Sci 2004; 3: 436-450.
- Макаров Д.А., Кузнецова Н.А., Страховская М.Г. и др. Поликатионные фталоцианины цинка и алюминия: синтез, влияние степени замещения на физико-химические свойства и фотодинамическую активность в водной среде. Журнал физической химии 2009; 6: 1183-1190.
- Kuznetsova N.A., Yuzhakova O.A., Strakhovskaya M.G., et al. New heterogeneous photosensitizers with phthalocyanine molecules covalently linked to aminopropyl silica gel. Journal of Porphyrins and Phthalocyanines 2011; 15: 718-726.
