Пещерные археологические памятники как объект палинологических реконструкций

Автор: Алексейцева В.В., Рудая Н.А.
Журнал: Проблемы археологии, этнографии, антропологии Сибири и сопредельных территорий @paeas
Рубрика: Археология каменного века палеоэкология
Статья в выпуске: т.XXVII, 2021 года.
Бесплатный доступ
Пещерные памятники зачастую становятся объектами палинологических реконструкций, однако, вопрос о репрезентативности пещерных отложений для палинологических исследований в связи с особенностями распространения и захоронения пыльцевых зерен в пещерах на настоящий момент является актуальным и недостаточно изученным. В настоящей статье представлены результаты палинологического изучения поверхностных образцов, отобранных на трех памятниках: пещеры Страшная, Чагырская и Бийка-1 (Алтай). Сопоставление результатов, полученных на основе исследования образцов внутри и снаружи пещер, показало их сходность, что подтверждает предположение о возможности использования подобных данных для построения климатических реконструкций. Однако выделяется ряд аспектов, которые необходимо учитывать при интерпретации палинологических данных, полученных из отложений пещерных памятников, что также подтверждается другими исследователями. К таким аспектам относятся: расположение места отбора образца: отмечается отрицательная корреляция между расстоянием от входа в пещеру и концентрацией пыльцы, а также содержанием анемофильной пыльцы в образце; деятельность животных внутри пещеры: построение животными логов или насекомыми гнезд внутри пещеры может являться причиной искажения палиноспектров; морфология пещеры, размеры и количество входов в пещеру: отмечается, что более показательными являются пещеры с одним достаточно широким входом, т.к. он может обеспечить достаточную циркуляцию воздуха в пещере; ориентация входов в пещеру: палиноспектры внутри пещеры могут отражать локальную растительность территории, на которую направлен вход в нее; особенности распространения пыльцевых зерен растений различных семейств: отмечено накопление пыльцы энтомофильных растений ближе к задней стенке пещеры; различная степень стойкости оболочек пыльцевых зерен различных семейств: пыльцевые зерна с более стойкой к постдепозиционным процессам оболочкой могут присутствовать в палиноспектрах в большей концентрации в силу разрушения зерен с менее стойкой оболочкой.
Пещерные археологические памятники, палинологический анализ, алтай
Короткий адрес: https://sciup.org/145146126
IDR: 145146126 | УДК: 902/904 | DOI: 10.17746/2658-6193.2021.27.0012-0019
Cave archaeological sites as an object of palynological reconstructions
Cave sites often become objects of palynological reconstructions; however, the question of the representativeness of cave deposits for palynological researches in connection with the specifics of the distribution and burial of pollen grains in the caves is currently relevant and insufficiently studied. This article provides the results of a palynological study of surface samples from three sites: Strashnaya, Chagyrskaya, and Biyka-1 caves (Altai Territory). The comparison of the results based on the study of samples inside and outside the caves showed their similarity, which confirms the assumption that such data can be used for climatic reconstructions. However, there are aspects necessary to be taken into consideration when interpreting palynological data obtained from the deposits of cave sites, which is also confirmed by other researchers. Such aspects include the following: location of the sampling site: there is a negative correlation between the distance from the cave entrance and the concentration of pollen, as well as the content of anemophilicplants pollen in a sample; animal activity inside the cave: animal roosting and denning behavior or insect nesting inside the cave may cause distortion of palinospectra; cave morphology, size and number of cave entrances: it was noted that caves with one sufficiently wide entrance are more indicative, because it can provide sufficient air circulation in the cave; orientation of the cave entrances: palinospectra inside the cave may reflect the local vegetation of the territory to which the entrance is directed; specificity of the distribution of pollen grains of plants of various families: it was noted that the accumulation of pollen of entomophilic plants closer to the back of the cave is higher than anemophilic ones; different degree of resistance of the shells of pollen grains of different families: pollen grains with a shell more resistant to post-deposition processes may be present in palinospectra in greater concentration due to the destruction of grains with a less resistant shell.
Текст научной статьи Пещерные археологические памятники как объект палинологических реконструкций
Пещерные археологические памятники являются уникальными объектами для изучения активности древнего человека и для реконструкции природных условий, в которых он жил [Day, 2013]. В областях с засушливым и полузасушли-вым климатом пещеры могут быть единственным источником палинологических данных. В то же время пещеры отличаются сложной стратиграфией и особенностями осадконакопления внутри них. Благодаря этому подход к палинологическому изучению пещер имеет свою специфику. Тафономические процессы внутри пещер, а также механизмы заноса пыльцы в пещеру и ее захоронения на настоящий момент еще недостаточно изучены. В связи с этим вопрос о степени достоверности палинологических записей, полученных на основе материалов пещерных памятников, остается актуальным и дискуссионным. С целью выявления достоверности палинологических исследований пещерных памятников нами были сопоставлены данные поверхностных сборов на трех памятниках, расположенных на Алтае: пещеры Страшная, Чагырская и Бийка-1.
Методические вопросы достоверности реконструкций растительности по пещерным палиноспектрам
Множе ством исследователей, изучавших пещерные объекты, не учитывался ряд факторов, способных искажать конечный результат, что могло приводить к ошибочным выводам. Остается дискуссионным вывод Р. Солецки, в 1960 г. обнаружившего в пещере Шанидар захоронения девяти неандертальцев возрастом 60 тыс. л. н. [Solecki, 1963]. Для памятника был проведен палинологический анализ, который показал высокое содержание цветочной пыльцы для одного из захоронений
(Шанидар-4), чего не наблюдалась в остальных погребениях и в других частях пещеры. Автор исследования высказывает предположение о захоронении этого неандертальца на подстилке из цветов и о возможном медицинском или экономическом значении подобного способа захоронения [Solecki, 1975].
В 2014 г. исследования пещеры Шанидар были возобновлены археологами М. Фиаккони и К. Хантом для проверки старых данных и оценки перспективности палинологических исследований для палеоэкологических реконструкций Ближнего Востока. Для этого исследователями были изучены поверхностные образцы из пещеры и с территории вокруг нее [Fiacconi, Hunt, 2015].
Авторами отмечены различные тенденции изменения процентного соотношения разных таксонов в зависимости от удаленности места отбора образца от входа в пещеру. Подобное явление они объясняют отличающимися способами переноса пыльцы: некоторые из растений являются зоофильными (опыляемыми животными), некоторые – анемофильными (опыляемыми ветром). Таким образом, пыльца растений, переносимая ветром, более представлена в передней части пещеры и менее у задней стенки, в то время как переносимая животными пыльца, напротив, более представлена сзади и практически отсутствует у входа в пещеру. При изучении пещерных памятников этот фактор следует рассматривать в совокупности с параметрами входа в пещеру и ее морфологией, от которых напрямую зависит, насколько далеко ветер может проникать вглубь конкретной пещеры.
В своем исследовании М. Фиаккони и К. Хант приходят к выводу, что в целом данные изнутри пещер репрезентативны, но отмечается, что пещера Шанидар имеет один вход до статочно большого размера, что обеспечивает занос пыльцы в пещеру из окружающей среды в большом объеме. Все се- мейства, обнаруженные Р. Солецки в захоронении неандертальца Шанидар-4, присутствуют внутри пещеры и в настоящее время, что может говорить об ошибочности выводов исследователя о намеренном захоронении особи в цветах и наличии у неандертальцев подобных практик.
Вопросом о степени достоверности полученных в пещере данных задавались и другие исследователи [Coles et al., 1989; Edwards et al., 2015; Hunt, Fiacconi, 2017]. Д. и Л. Берни изучили три пещеры на территории штата Нью-Йорк [Burney D.A., Burney L.P., 1993]. Большинство с емейств растений, опыляемых ветром, оказались представлены внутри и снаружи пещер как с большим размером входа, так и с малым, приблизительно в одинаковом процентном соотношении. Исключение для одной из пещер составил вяз ( Ulmus ) – его процентное содержание в одном из образцов изнутри пещеры было крайне высоко в отличие от других таксонов. Авторы предполагают, что вяз в большом количестве мог быть принесен животными, следы обитания которых были замечены в данной пещере.
Деятельность животных и человека может являться причиной чрезмерной представленности тех или иных таксонов в палинокомплексе. Нередки случаи построения гнезд пчелами внутри пещер. Было выявлено, что пчелиные гнезда являются накопителем энтомофильной пыльцы, в частности, семейства сложноцветных. В связи с этим особо важным представляется отмечать присутствие следов обитания животных и насекомых в изучаемой пещере. Отмечено, что даже наличие бывшего пчелиного гнезда на месте исследования может приводить к искажениям конечного результата [Fiacconi, Hunt, 2015, p. 91].
В 2002 г. Наварро и др. были изучены поверхностные образцы снаружи и внутри двух пещер, располагающихся на территории Испании [Navarro et al., 2002]. Авторы отмечают зависимость результата исследования от морфологии пещер. Куэва де ла Плата – узкая длинная пещера с небольшим входом, показала более низкую концентрацию пыльцы, в отличие от Куэва де Жозе, обладающей широким входом. В обеих пещерах преобладают зо-офильные виды растений, но в более высокой концентрации они присутствуют в пещере Куэва де ла Плата, вероятнее всего по причине того, что занос пыльцы с помощью ветра в эту пещеру более проблематичен.
Образцы, отобранные из влажных отложений, показали более высокое содержание пыльцы подсемейства цикориевых, в то время как концентрация всей пыльцы и разнообразие представленных видов в этих образцах были заметно ниже. Это мо-14
жет объясняться тем, что пыльца из влажных отложений более подвержена постдепозиционным процессам и соответственно обладает худшей сохранностью [Burney D.A., Burney L.P., 1993, p. 523]; пыльцевые зерна цикориевых в свою очередь, вероятно, более стойкие к этим процессам и сохраняются в подобных отложениях лучше пыльцы других семейств. Было доказано, что сильное разрушающее воздействие на пыльцевые зерна оказывают часто сменяющиеся циклы влажных и сухих условий [Campbell, 1991].
Израильской исследовательницей М. Вайн-штайн-Эврон было также отмечено, что пещерные спектры могут скорее отражать локальную растительность, чем региональную, и подчеркнута необходимость повышенного внимания при попытках реконструкции региональной растительно сти по пещерным данным [Weinstein-Evron, 1994].
Методы и материалы
Для выявления степени достоверности пещерных палинологических данных для палеоэкологических реконструкций нами проанализированы поверхностные образцы, отобранные изнутри и снаружи трех пещерных памятников, располагающихся на территории Алтая: Чагырская, Страшная и Бийка-1.
Для памятника Бийка-1 в 2020 г. С.В. Шнайдер на палинологический анализ было отобрано семь образцов из отложений внутри пещеры, 10 образцов – из разреза снаружи. 20 поверхностных образцов было отобрано с пяти зон, по четыре образца с каждой: образцы ID 1.1-1.4 – внутри пещеры Бийка-1, ID 2.1-2.4 – снаружи у входа в пещеру Бий-ка-1, ID 3.1-3.4 – у входа в грот Бийка-2, ID 4.1-4.4 – около дороги в окрестностях пещерного комплекса и ID 5.1-5.2 – около реки Бийка, протекающей неподалеку от пещерного комплекса. Анализ был проведен для шести образцов (два образца из пещеры и по одному из остальных зон).
Пробоподготовка выполнялась по методике, разработанной Фаегри-Иверсеном в 1989 г. [Рудая, 2011]. Химическая обработка, применяемая в лаборатории PaleoData ИАЭТ СО РАН, включает в себя следующие этапы: 1) обработка 10-процентным раствором соляной кислоты HCl; 2) нагревание на водяной бане до температуры 90° в течение 10 минут с 10-процентным раствором гидроксида калия KOH; 3) ситование образца на сите с ячейкой 200-250 мкм; 4) нагревание с 40-процентным раствором плавиковой кислоты HF на водяной бане до температуры 90° в течение 2 часов; 5) нагревание на водяной бане до температуры 90° в течение 10 минут с 10-процентным раствором соля- ной кислоты HCl; 6) ситование на сите с ячейкой 7 мкм; 7) центрифугирование в течение 20 мин и декантирование; 8) добавление глицерина 85 %. После этапов 1, 2 и 5 образцы необходимо центрифугировать в течение 4 минут, декантировать, затем двукратно центрифугировать с дистиллированной водой. Для подсчета концентрации пыльцы и спор в образце на этапе пробоподготовки добавляется одна таблетка, содержащая ацетолизированные споры Lycopodium.
Микроскопирование осуществлялось с помощью светового микроскопа Zeiss AxioImager D2 с увеличением ×400. Таксономическая принадлежность пыльцевых зерен определялась путем сравнения с компаративной коллекцией ИАЭТ СО РАН, а также с помощью определителей и атласов. Результаты отражены в палинологических диаграммах, построенных в программе Tilia.
Поверхностные образцы на памятниках Страшная и Чагырская пещеры были отобраны в 2016 г. Отбор производился из разных точек внутри пещеры и в ее окрестностях для того, чтобы понять, адекватно ли пещерные палинокомплексы отражают современный им растительный покров. Про-боподготовка образцов была проведена в 2019 г. в палинологической лаборатории ИАЭТ СО РАН, однако, образцы тогда не были проанализированы. Для этих образцов в рамках настоящего исследования нами построены палинологические диаграммы с помощью программы Tilia и произведен анализ результатов.
Результаты и их интерпретация
Для пещеры Чагырская проанализированы данные по двенадцати поверхностным образцам: двум из пещеры, отобранным на расстоянии 4 (ID1) и 2 (ID2) метра от входа, и десяти (ID3-12) – с разных участков в ее окрестностях. В пещере выявлено преобладание березы и полыни . Высокое содержание показала сосна обыкновенная . Травянистые представлены астровыми и злаковыми (ок. 10 %), осоковыми, крапивными, маревыми, цикориевыми, яснотковыми (менее 5 %) (рис. 1).
В спектрах снаружи пещеры абсолютное преобладание показали хвойные, представленные соснами обыкновенной и сибирской, елью, пихтой, лиственницей. В составе травянистых, помимо таксонов, присутствующих в пещерных спектрах, выявлены розовые, бобовые, лютиковые (менее 5 %).
Можно заключить, что в целом палиноспек-тры внутри и снаружи пещеры Чагырская сходны между собой: выявлены одни и те же семейства, представленные в сходных соотношениях. Раз-
dOU3 и АЬяини з шпицоиие eieLMujaiapui dds euBN aeqivves

Рис.1. Палинологическая диаграмма поверхностных сборов на памятнике Чагырская пещера.
ееерюиоцэо.
eeepiojeisy.
еревья и кустарники Травы Споры
9B9dBqBiI_ 9dAq) 9B936|ndunuBy-L 9В9ЭВ|ПЭиПиба_Е.
9B93B3ISSBjgj_
9B93B0d^
9B95BWB"lL anaipiadA ^J 9B936ipOdOU0l|9_L 9B9dB||AqdoAjB3_L
у - о
-
=5 я
S
J9iseu|v_L ai -г
doua и 1ЯРяшяи вииАэ-
Х9ШПу_Е. 9B93Biqnyl.
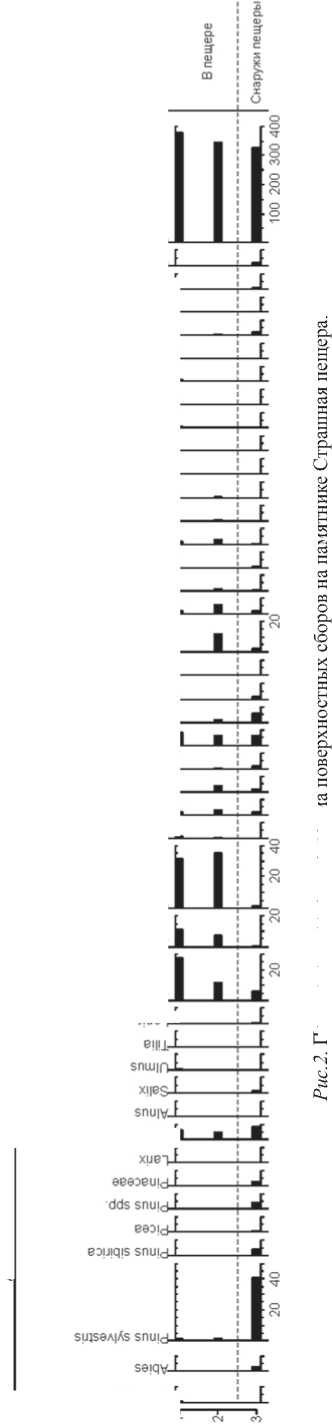
9B93Bj6BUO-L шnЛBJ9Л uiniAdofis j I 9B93BU06A|0 9B93BldV-L зпшАщ p± 9B93BS0a_L J919PUI 96qj9H-L Bisinieijy. BJ93IU0T-E dds einjeai нятся, однако, показатели древесной растительности в составе: преобладающая внутри пещеры береза практически не представлена снаружи, обратную ситуацию можно наблюдать относительно хвойных. Полынь и астровые также более представлены внутри пещеры. Относительно полыни и березы можно отметить корреляцию между процентным соотношением пыльцы в составе и расстоянием от входа в пещеру: образец ID1, отобранный дальше от входа, показывает более высокое содержание этих таксонов, чем ID2. Сосна, напротив, больше представлена в образце, отобранном ближе к входу в пещеру. Именно те таксоны, которые практически не представлены снаружи, обнаруживают большую концентрацию в образце, отобранном ближе к задней стенке пещеры, чем у входа в нее. Для пещеры Страшная палиноспектры показали более значительные отличия, чем для Ча-гырской. Проанализированы данные по трем поверхностным образцам: двум изнутри пещеры, отобранным у задней стенки в левом (ID1) и правом (ID2) углах, и одному снаружи, на расстоянии 3 метра от входа в пещеру (ID3) (рис. 2). Внутри пещеры выявлено абсолютное преобладание цикориевых – 32 %, и полыни – 28 %. В меньшей степени представлены злаки и астровые (ок. 8–10 %), а также бобовые для одного из двух образцов. Содержание остальных таксонов в составе не превышает 1–2 %. Спектры снаружи показали абсолютное преобладание сосны обыкновенной (40 %), в количестве 5–10 % присутствуют береза, полынь, злаки, капустные, другие таксоны представлены в количестве менее 5 %. Состав таксонов в палиноспектрах внутри и снаружи пещеры приблизительно сходен. Разную картину можно наблюдать относительно процентного соотношения сосны обыкновенной и цикориевых: в то время как снаружи сосна преобладает, внутри она практически не представлена, обратная тенденция отмечена относительно цикориевых. Полынь внутри пещеры также представлена в большей концентрации, чем снаружи. Другие таксоны присутствуют в сходных соотношениях. Вероятно, подобная ситуация может быть связана с морфологическими особенностями пещеры: узкая вытянутая форма пещеры не способствует глубокому проникновению воздушных потоков, что может приводить к меньшей концентрации у задней стенки пыльцы ветроопыляемых растений, к каковым относится сосна, и, соответственно, большей концентрации насекомоопыляемых, в частности, семейства сложноцветных. Для пещеры Бийка-1 получены данные по шести поверхностным образцам: двум внутри пеще- ры (ID1.1-1.2), одному у входа в пещеру (ID2.1), одному у входа в грот Бийка-2 (ID3.1), одному у реки Бийка, протекающей в окрестностях Бий-кинского пещерного комплекса (ID4.1) и одному у дороги неподалеку от пещерного комплекса (ID5.1) (рис. 3). Для образцов ID1.1-1.2 и ID2.1 были выявлены сходные палиноспектры. Преобладают цикориевые, лютиковые, розовые, в меньшей концентрации представлены полынь, ива, бобовые, норичниковые, астровые. Образцы из зон 3–5 заметно отличаются. Преобладают семейства осоковых и подорожниковых, снижается роль лютиковых и розоцветных. Возрастает роль древесных, представленных сосной, березой, появляются вяз, пихта, лиственница, отсутствовавшие в предыдущих спектрах. Цикориевые, абсолютно преобладавшие в образце внутри пещеры и у входа, в остальных образцах практически не представлены. Ива и норичниковые, присутствовавшие в зонах 1 и 2, в зонах 3–5 также практически отсутствуют. Остальные таксоны присутствуют в сходных соотношениях. Таким образом, палино спектры пещеры Бий-ка-1 недостаточно отражают древесную растительно сть и, напротив, показывают чрезмерную представленность подсемейства цикориевых. Подобно пещере Страшной, это может объясняться разницей в способах опыления растений различных семейств. Внутри пещеры и у входа в нее преобладает энтомофильная растительность, к каковой относятся цикориевые, лютиковые, розоцветные. В то же время снаружи более представлена анемофильные (осоковые, подорожниковые) и древесные таксоны. На основе полученных результатов можно сделать вывод, что в пещерных палиноспектрах в приблизительно сходном процентном соотношении присутствуют те же таксоны, что и снаружи, что может говорить о том, что материалы пещерных памятников можно использовать для построения реконструкций. Однако некоторые таксоны показали разные значения. Для пещеры Страшной изнутри выявлено абсолютное преобладание цикориевых, в то время как снаружи они практически не присутствовали, и, напротив, отсутствие сосны, преобладающей снаружи. Схожая ситуация отмечена относительно Бийки-1: внутри преобладают цикориевые, которых снаружи практически нет, но почти отсутствует пыльца древесных растений, обнаруженная снаружи. Судя по всему, для пещерных палиноспектров характерна большая представленность насекомоопыляемых таксонов, к каким относятся цикориевые, лютиковые, розоцветные, и меньшая представленность ветроопыляемых, таких как сосновые, осоковые, подорожниковые. aeaoesoy. eeaoejedAo- эвэов1пэипив; эвээв|пшис^ эвэовио6А|о^-к эвээвоа-к aBaoBui6B|UB|d-L-aBaoBpiqojQ-L авэовип-L авээв!|!3-к авээв1шв>1-aeaoeqe-pL ЭВЭЭВ|ПЛ|ОЛиОЭ-Ь douo и i<*iqui4u bwwAq. Рис.3. Палинологическая диаграмма поверхностных сборов на памятнике Бийка-1. |эрш aBqjdH-8B80Buaqja/vL эвээвивиа|ВлЛ-aBaoBoiyn-L ajnjpqBqi-L хившв1-1 eeeoeyBinjojos-L—a^. aeapiouoqoio. aeaoeipodouaqo-h-aeaoe||AqdoAjeo-L эвээвшо|пд_Ь aBdOBOissBjg-L aeapiojatsv-k aBdOBidv-L dBaoBinv-L dBaoBiBiusiiV-L В181ШЭр^-к эваэвэиэ-L Bjpaqdg-L sn|ndod-L snojano-L ds е|щэ! Для пещеры Чагырской были отмечены незначительные различия в процентных соотношениях в спектрах, однако в целом спектры были достаточно сходны друг с другом. Это может говорить о том, что, по-видимому, Чагырская пещера является достаточно широкой, чтобы обеспечивать лучшую циркуляцию воздуха, и может лучше отражать внешнюю растительность, чем Страшная и Бийка-1. Можно отметить, что места сбора поверхностных образцов из Чагырской пещеры находились до статочно близко от входа в пещеру, в то время как в Бийке-1 и Страшной образцы отбирались ближе к задней стенке, что также может способствовать более высокой репрезентативности результатов. Выводы В целом пещерные палиноспектры достаточно достоверно отражают внешнюю растительность, однако, существует ряд факторов, которые необходимо учитывать при интерпретации таких спектров. Выделяются следующие факторы. 1. Параметры и количе ство входов в пещеру и морфология пещеры; 2. Расстояние от места отбора образца до входа в пещеру; 3. Особенности распространения пыльцы различных семейств; 4. Постдепозиционные процессы, по-разному влияющие на пыльцу различных семейств; 5. Деятельность животных внутри пещеры, например, гнездование насекомых или обустройство животными логова, либо деятельность человека. Палинологическое исследование трех объектов, располагающихся на территории Алтая (Бийка-1, Страшная, Чагырская) показало, что построение палеоэкологических реконструкций на основе пещерных данных представляется возможным, однако, полученные результаты требуют особо внимательной интерпретации. Разница в спектрах для пещер Страшная и Бий-ка-1 относительно таких таксонов, как цикориевые, сосна и в меньшей степени лютиковые, осоковые, розоцветные, по-видимому, может быть объяснена морфологическими особенностями пещер и местом обора образцов на анализ, а также особенностями распространения пыльцы этих семейств, часть из которых является анемофильной, другая – энтомофильной. Отмечается, что пыльца энтомофильных растений имеет тенденцию к преобладанию ближе к задней стенке пещеры. Сходные палиноспектры внутри и снаружи пещеры Чагырской, достаточной широкой по своей морфологии, образцы из которой отбирались бли-18 же к входу, могут говорить о том, что этот объект является более репрезентативным для палеоэкологических реконструкций, чем пещеры Страшная и Бийка-1, а также о том, что лучше внешнюю растительность отражают образцы, отобранные ближе к входу в пещеру.

Список литературы Пещерные археологические памятники как объект палинологических реконструкций
- Рудая Н. А. Палинологический анализ // Методические подходы к использованию биологических индикаторов в палеоэкологии / научн. ред. Л.Б. Назарова. - Казань: Казан. ун-т, 2011. - 280 с.
- Burney D. A., Burney L. P. Modern pollen deposition in cave sites: experimental results from New York State // New Phytologist. - 1993. - Vol. 124. - P. 523-535.
- Campbell I. Experimental mechanical destruction of pollen grains // Palynology. - 1991., - no 15. - P. 29-33. https://doi.org/10.1080/01916122.1991.9989387.
- Coles G.M., Gilbertson D., Hunt C., Jenkinson R.D.S. Taphonomy and the palynology of cave deposits // Cave Science - Transactions British Cave Research Association. - 1989, - no 16. - P. 83-89.
- Day J. Botany meets archaeology: people and plants in the past // Journal of Experimental Botany. - 2013. - Vol. 64, no 18. - P. 5805-5816. https://doi.org/10.1093/jxb/ert068
- Edwards K., Fyfe R., Hunt C., Schofield J. Moving forwards? Palynology and the human dimension // Journal of Archaeological Science. - 2015, - no 56. - P. 117-132. https://doi.org/10.1016/j.jas.2015.02.010
- Fiacconi M., Hunt C. O. Pollen taphonomy at Shanidar Cave (Kurdish Iraq): An Initial evaluation // Review of Palaeobotany and Palynology. - 2015. -Vol. 223. -P. 87-93.
- Hunt C.O., Fiacconi M. Pollen taphonomy of cave sediments: What does the pollen record in caves tell us about external environments and how do we assess its reliability? // Quaternary International. - 2017. - Vol. 485. - P. 68-75.
- Navarro C., Carriyn J. S., Prieto A. R., Munuera M. Modern Cave pollen in an arid environment and its application to describe palaeorecords // Complutum. -2002. - Vol. 13. - P. 7-18.
- Solecki R. S. Prehistory in Shanidar Valley, Northern Iraq // Science. - 1963. - Vol. 139. - P. 179-193.
- Solecki R. S. Shanidar IV, a Neanderthal Flower Burial in Northern Iraq // Science. - 1975. - Vol. 190. - P. 880-881.
- Weinstein-Evron M. Biases in archaeological pollen assemblages: Case studies from Israel // Contributions to Microbiology. - 1994, - no 29. - P. 193-205.
