Плейотропное влияние мутаций высокопальмитиновости и высокоолеиновости масла семян на морфологические признаки растения подсолнечника

Автор: Демурин Я.Н., Борисенко О.М., Бочкарев Н.И.
Статья в выпуске: 1 (142-143), 2010 года.
Бесплатный доступ
Плейотропный эффект мутаций высокопальмитиновости (р) и высокоолеиновости (Ol) наблюдался в течение трѐхлетнего исследования серии аналогов линии ВК580. Мутация р приводит к уменьшению высоты растений на 41 см, числа листьев на 4 штуки и периода всходы-цветение на 2 дня. Мутация Ol достоверно увеличила высоту растений на 4 см. Изучение варьирования высоты растений в F2 и содержания пальмитино-вой кислоты в семенах F3 позволило установить факт ассоциативного наследования этих признаков при скре-щивании линий ВK850(р)×ВK508. Средняя высота гомозиготных высокопальмитиновых растений в F2 равнялась 109 см и была достоверно ниже высоты остальных растений 138 см.
Аналоги линии, подсолнечник, мутация, жирная кислота
Короткий адрес: https://sciup.org/142150924
IDR: 142150924
Pleiotropic effect of high palmitic and high oleic mutation in seed oil on morphological traits in sunflower
Pleiotropic effect of a high palmitic (p) and high oleic (Ol) mutations in sunflower seed oil was investigated on the genetic base of VK580 near-isogenic lines in the field conditions. It was found out that p mutation decreased plant height by 41 cm, quantity of leaves by 4 pieces and the period of emergence-flowering by 2 days. Ol mutation increased significantly plant height by 4 cm. Moreover, the fact of associated inheritance of high palmitic mutation with plant height in the case of selfing of F2 plants in the cross of VK850(p)×VK508 was observed. Average plant height of the high palmitic homozygotes in the segregated F2 population was lower and estimated as 109 cm with comparing to 138 cm for other plants. Thus, negative effect of the high palmitic mutation on plant height was certainly proved.
Текст научной статьи Плейотропное влияние мутаций высокопальмитиновости и высокоолеиновости масла семян на морфологические признаки растения подсолнечника
Введение. Влияние мутаций жирно-кислотного состава масла семян подсолнечника на другие селекционные признаки привлекало внимание учёных в ряде исследовательских программ.
Наиболее значимая работа была посвящена сравнительной оценке обычных и высокоолеи-новых гибридов по комплексу хозяйственно ценных признаков в различных экологических условиях в Испании [5]. Изогенные компоненты четырёх селекционных линий RHA271, RHA274, HA89 и HAJ152 не показали воздействия мутации высокоолеиновости Ol , полученной К.И. Солдатовым во ВНИИМК [3], на высоту растений, число листьев, диаметр корзинки, период всходы–цветение, автофертильность и масличность семянок [5]. С другой стороны, мутация Ol обладала позитивным эффектом на урожай семянок, их масличность и надземную биомассу, а также негативно воздействовала на период всходы–цветение, автофертильность и всхожесть у четырёх пар полученных изогенных гибридов. Кроме того, наблюдалось достоверное взаимодействие признак × генетическая среда [5].
Мутация высокопальмитиновости масла семян подсолнечника впервые получена П. Ивановым в Болгарии [6] после использования физического мутагенеза при облучении гамма-лучами семянок сорта Заря селекции ВНИИМК. Самоопылённая линия получила название 275 HP. Опубликованных данных о влиянии этой мутации на другие признаки растения нам не известно.
В работах испанских учёных сообщается о создании высокопальмитиновой линии CAS-5 после облучения семянок обычной линии рентгеновскими лучами [7, 8]. Плейотропный эффект мутации также не описан. С другой стороны, другой высокопальмитиновый мутант P-20, полученный после химического мутагенеза при обработке семянок обычной линии этилметансульфонатом (EMS), потерял всхожесть в М3, показывая наличие летального фактора в гомозиготе [7]. Кроме того, ещё одна высокопальмитиновая мутация была индуцирована нитрозоэтилмочевиной (НЭМ) в рамках программы химического мутагенеза для улучшения селекционных линий подсолнечника в Украине [2].
В ходе гибридологического анализа установлено, что наследование мутации высокопаль-митиновости р у линии CAS-5 контролируется одним или тремя генами в зависимости от комбинации скрещивания [9].
В ходе генетических исследований во ВНИИМК три независимо обнаруженные мутации высокопальтиновости были утеряны при размножении растений из-за пониженной жизнеспособности. Это произошло в M 3 после обработки семянок сорта Юбилейный 60 нитрозоэтилмочевиной, в I 2 при самоопылении растений образца K2222 и, наконец, в I 1 образца №429934 мировой коллекции ВИР. Тем не менее, высокопальмитиновая линия ЛГ30 была получена во ВНИИМК в результате скрининга ЦСМ-линий из Украины с последующим скрещиванием, индивидуальным отбором и самоопылением [1, 4].
Данная работа была направлена на изучение влияния мутаций высокоолеиновости и высо-копальмитиновости масла семян на некоторые морфометрические признаки растений с использованием линий генетической коллекции подсолнечника ВНИИМК.
Материал и методы. В исследовании использовали серию аналогов селекционной линии ВК580 по мутациям Ol и р . ВК580 является нормой, ВК508 обладает мутацией высокоолеиновости (87 %), ВК850 – мутацией высокопальмитиновости (24 %), а ВК805 содержит обе мутации высоко-олеиновости (6 %) и высокопальмитиновости (20 %) в дигомозиготе. Мутация высокопальмитино-вости происходит из линии ЛГ30 после ВС3 на линию ВК580.
Растения выращивали летом в полевых условиях на центральной экспериментальной базе ВНИИМК в 2007-2009 гг. Посев проводили по схеме 70×35 см по две семянки в гнездо. Полевая всхожесть семян высокопальмитиновых аналогов ВК850 и ВК805 была существенно ниже по сравнению с нормой ВК580. На стадии 3-х пар настоящих листьев проводили рендомизированную прорывку до одного растения в гнезде. Для скрещивания применяли ручную кастрацию материнской формы.
Жирно-кислотный состав масла определяли с помощью газо-жидкостной хроматографии метиловых эфиров на приборе Хром 5. Обработка экспериментальных данных проводилась с помощью программы Statistica 6.0.
Результаты и обсуждение . Плейотропный эффект мутаций высокопальмитиновости и вы-сокоолеиновости наблюдался в течение трёхлетнего исследования серии аналогов линии ВК580 (табл. 1). Достоверно установлено, что мутация р приводит к уменьшению высоты растений на 41 см (32 %), числа листьев на 4 штук (14 %) и периода всходы-цветение на 2 дня (4 %). Мутация Ol достоверно увеличила высоту растений на 4 см (3 %).
Доля влияния мутаций р и Ol (фактор генотипа) в общем варьировании высоты растений составила 46 % (табл. 2), что является значительной величиной с учётом различных экологических условий трёхлетнего исследования. При этом влияние всех факторов, включая генотип, год, а также их взаимодействие, было статистически достоверным для трёх изученных признаков. Фактор года оказывал основное влияние на период всходы–цветение и число листьев на 83 и 73 % соответственно.
Таблица 1 – Среднее значение и отклонение от стандарта морфометрических признаков различных генотипов подсолнечника
ВНИИМК, г. Краснодар, 2007-2009 гг.
|
Генотип |
Высота растений, см |
Период всходы-цветение, дни |
Число листьев, шт. |
|||
|
X |
± st |
X |
± st |
X |
± st |
|
|
ВК580 (st) |
127 |
0 |
54 |
0 |
28 |
0 |
|
ВК508 Ol |
131 |
+4* |
54 |
0 |
29 |
+1 |
|
ВК850 p |
86 |
-41* |
56 |
+2* |
24 |
-4* |
|
ВК805 Ol, p |
89 |
-38* |
55 |
+1* |
26 |
-2* |
|
* - различия достоверны, p< 0,05 |
||||||
Таблица 2 – Доля влияния факторов в общем варьировании признаков у аналогов линии ВК580
ВНИИМК, г. Краснодар, 2007-2009 гг.
|
Фактор |
Высота растения |
Период всходы-цветение |
Число листьев |
|
генотип |
0,46 |
0,07 |
0,11 |
|
год |
0,43 |
0,83 |
0,73 |
|
генотип×год |
0,03 |
0,02 |
0,06 |
|
остаточная |
0,08 |
0,08 |
0,10 |
С целью верификации существенного негативного эффекта мутации высокопальмитиново-сти на высоту растений, изначально обнаруженного у серии аналогов, был проведён тест на сопряжённое наследование этих двух признаков (рис. 1).
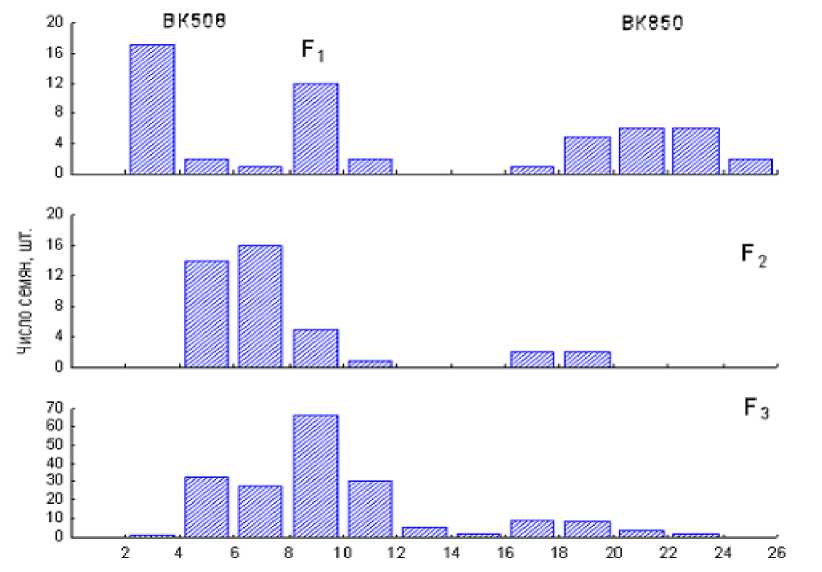
Пальмитиновая кислота, %
Рисунок 1 – Частота распределения значений содержания пальмитиновой кислоты в отдельных семенах родительских линий ВК508 и ВК850, F1, F2 и средних проб семян F3 с самоопылённых корзинок F 2 ВНИИМК, г. Краснодар, 2007-2009 гг.
Изучение семян F 1 от скрещивания аналогов ВK850( р )×ВK508 показало, что низкое содержание пальмитиновой кислоты является неполно доминирующим признаком со степенью доминирования d = -0,4. При этом среднее значение содержания пальмитиновой кислоты у родительских форм было 21 и 3 % соответственно, а у гибридных семян F 1 – 9 %.
Отдельные семена F 2 показали расщепление по содержанию пальмитиновой кислоты на два дискретных фенотипических класса: нормальный и промежуточный (4-10%) и высокопальмитиновый (17-19 %). Соответствующее эмпирическое расщепление 36:4 согласуется с модельным отношением 15:1 для двух рецессивных несцепленных генов (χ2 15:1 = 0,96, p>0,05). Другая часть семян этой популяции F2 была использована для посева, измерения высоты растений и их самоопыления.
Средние пробы семян F3 с 188 самоопылённых корзинок F2, как и следовало ожидать, показали расщепление по содержанию пальмитиновой кислоты на два фенотипических класса: нормальный и промежуточный (4-17 %) и высокопальмитиновый (18-23 %). Это эмпирическое расщепление 173:15 также достоверно соответствует схеме 15:1 с одинаковым значением χ215:1 = 0,96, p>0,05.
Изучение варьирования высоты 188 растений в F 2 и содержания пальмитиновой кислоты в семенах F 3 позволило установить факт ассоциативного наследования этих признаков при скрещивании линий ВK850×ВK508 (рис. 2). При этом коэффициент корреляции составил r = –0,54, а уравнение линейной регрессии имеет вид y = 157,8 – 2,3x.
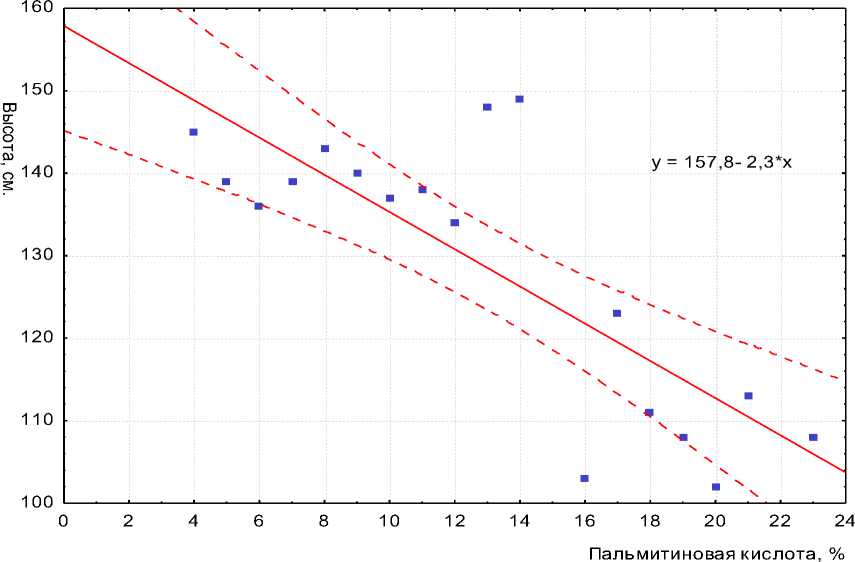
Рисунок 2 – Регрессия высоты растений F2 по содержанию пальмитиновой кислоты в масле семян F 3 , полученных при самоопылении. ВНИИМК, г. Краснодар, 2009 г.
Средняя высота 15 гомозиготных высокопальмитиновых растений в F2 равнялась 109 см и была достоверно ниже высоты остальных 173 растений, обладающих высотой 138 см (НСР05= 3 см). Очевидно, что это различие на 29 см по высоте между двумя фенотипическими классами в F 2 соответствует различию между родительскими формами ВК850 и ВК508.
Выводы. Существенное отрицательное влияние мутации высокопальмитиновости масла семян на высоту растений было установлено на основе как сравнительного изучения серии аналогов линии ВК580, так и в ходе гибридологического анализа. Генетический контроль мутации высо-копальмитиновости в скрещивании ВK850×ВK508 осуществлялся двумя независимо наследуемыми рецессивными генами. Основной вопрос заключается в поиске объяснения негативного
МАСЛИЧНЫЕ КУЛЬТУРЫ. Научно-технический бюллетень Всероссийского научно-исследовательского института масличных культур. Вып. 1 (142-143), 2010
воздействия мутации высокопальмитиновости на высоту растений как сцеплением генов, так и истинной плейотропией мутации.
