Поиск антагонистов возбудителя фомоза рапса озимого: первичный скрининг бактериальных штаммов из рабочей коллекции лаборатории биометода ФГБНУ ФНЦ ВНИИМК

Автор: Маслиенко Л.В., Заверюха Е.А., Дейнега Л.А., Кузнецова А.В.
Рубрика: Агрохимия, агропочвоведение, защита и карантин растений
Статья в выпуске: 3 (199), 2024 года.
Бесплатный доступ
В 2024 г. в лаборатории биометода агротехнологического отдела ФГБНУ ФНЦ ВНИИМК с целью разработки микробиологического метода снижения вредоносности наиболее опасной болезни озимого рапса - фомоза (Leptosphaeria maculans (Desm.) Ces. et de Not, анаморфная стадия Phoma lingam (Tode) Desm.) проводили поиск перспективных штаммов-продуцентов микробиопрепаратов из рабочей коллекции лаборатории. В результате первичного скрининга бактериальных штаммов антагонистов выделено 29 перспективных штаммов с разными типами антагонистической активности против возбудителя фомоза: 21 штамм бактерий рода Bacillus: Fa 4-2, D-10, 11-2, D 1-1, 11-3, 5-3, 3-3, 3-2, Far 8, D 1-3, Fz 9, D 7-3, K1-2, 01kopf Bacillus sp., Б (2-1), Б-5, Б-12 B. licheniformis, 5Б-1, D 7-1, ВВ(С) B. subtilis, Б-4 B. circulans и восемь штаммов рода Pseudomonas: 12-2, 15-1, 13-2, Sgc-1, 16-2, Oif 2-1, 14-4 Pseudomonas sp. и 14-3 P. chlororaphis, для проведения вторичного скрининга на фоне искусственного заражения патогеном.
Фомоз рапса озимого, первичный скрининг, бактериальные штаммы антагонисты
Короткий адрес: https://sciup.org/142243085
IDR: 142243085 | УДК: 633.853.494:632.937 | DOI: 10.25230/2412-608X-2024-3-199-46-54
Search of antagonists of phoma rot pathogen on winter rapeseed: initial screening of bacterial strains from the working collection of the biometod laboratory of VNIIMK
In 2024, in the laboratory of biomethod of the V.S. Pustovoit All-Russian Research Institute of Oil Crops, a search of promising strains-producers of microbiopreparations from the working collection of the laboratory was conducted. The purpose of the research was to develop a microbiological method to decrease the harmfulness of the most dangerous disease on winter rapeseed - Phoma rot (Leptosphaeria maculans (Desm.) Ces. et de Not, anamorph stage Phoma lingam (Tode) Desm.). As a result of the initial screening of bacterial antagonist strains, 29 promising strains with the different types of an antagonist activity against Phoma pathogen were selected: 21 bacterial strains from a genus Bacillus: Fa 4-2, D-10, 11-2, D 1-1, 11-3, 5-3, 3-3, 3-2, Far 8, D 1-3, Fz 9, D 7-3, K 1-2, 01kopf Bacillus sp., Б (2-1), Б-5, Б-12 B. licheniformis, 5Б-1, D 7-1, ВВ(С) B. subtilis, Б-4 B. circulans, and eight strains from a genus Pseudomonas: 12-2 , 15-1, 13-2, Sgc-1, 16-2, Oif 2-1, 14-4 Pseudomonas sp. and 14-3 P. chlororaphis, to conduct the secondary screening on a pathogen artificial inoculation background.
Текст научной статьи Поиск антагонистов возбудителя фомоза рапса озимого: первичный скрининг бактериальных штаммов из рабочей коллекции лаборатории биометода ФГБНУ ФНЦ ВНИИМК
Введение. Одной из наиболее вредоносных болезней рапса озимого ( Brassica napus L.) является фомоз. Возбудителем в центральной зоне Краснодарского края установлен Leptosphaeria maculans (Desm.) Ces. et de Not, анаморфная стадия Phoma lingam (Tode) Desm. [1].
Поражение посевов рапса фомозом приводит к их изреживанию, уменьшению ассимиляционной поверхности растений, снижению массы 1000 семян, их посевных качеств, технологических свойств, мас-личности, ухудшению жирно-кислотного состава и к значительным потерям урожая (до 40 %). При инфицировании растений рапса озимого фомозом в первые фазы развития возбудитель может вызывать их преждевременную гибель [2; 3; 4]. Также все чаще отмечается эпифитотийное развитие болезни на рапсе в южных областях Нечерноземья. Это объясняется резким увеличением площади посевов, минимальным сроком возврата культуры на прежнее место, значительным движением сортового материала как внутри страны, так и за ее пределами, недостаточной генетической и сортовой устойчивостью [5–7]. Основным способом защиты рапса озимого от фомоза является применение фунгицидов [8; 9]. Зарегистрированных биопрепаратов от фомоза на рапсе в России нет. Поэтому разработка микробиологического метода снижения вредоносности одной из наиболее распространенных и опасных болезней рапса – фомоза – является актуальной задачей.
В лаборатории биометода агротехноло-гического отдела ФГБНУ ФНЦ ВНИИМК многие годы ведутся исследования по разработке микробиологических средств защиты масличных культур от болезней. В основе разработанной в лаборатории концепции целенаправленного создания микробиопрепаратов для защиты подсолнечника и других сельскохозяйственных культур от болезней лежит поиск штаммов антагонистов. безопасных для человека, не-фитотоксичных, проявляющих высокую активность в широко варьируемых условиях против комплекса патогенов, обладающих полифункциональным типом действия [10]. Концепция включает следующие основные этапы:
-
- изыскание в естественных условиях штаммов антагонистов возбудителей болезней;
-
- ступенчатый скрининг штаммов в лабораторных условиях in vitro и на фоне искусственного заражения во влажной камере и в грунте;
-
- селекционное улучшение перспективных штаммов, безопасных для человека и нефитотоксичных для растений;
-
- разработку препаративных форм, регламентов производства и хранения микробиопрепаратов;
-
- обоснование стратегии применения микробиопрепаратов в системе защитных мероприятий.
В результате многолетних исследований в лаборатории создана рабочая коллекция перспективных штаммов грибов и бактерий антагонистов широкого круга возбудителей болезней масличных и других сельскохозяйственных культур.
Одним из основных этапов концепции целенаправленного создания микробиопрепаратов является ступенчатый скрининг штаммов в лабораторных условиях in vitro и на фоне искусственного заражения во влажной камере и в грунте. Настоящая работа посвящена первому этапу ступенчатого скрининга – первичному скринингу коллекционных бактериальных штаммов к агрессивному изоляту возбудителя фомоза озимого рапса Phoma lingam in vitro.
Материалы и методы. Исследования проводили в лаборатории биометода агро-технологического отдела ФГБНУ ФНЦ ВНИИМК в 2024 г. Объектом исследований служили: конидиальная стадия агрессивного изолята возбудителя фомоза рапса озимого Phoma lingam , выделенного в центральной зоне Краснодарского края; коллекционные штаммы бактериальных антагонистов возбудителей болезней масличных культур – 27 штаммов рода Bacillus (01корf Bacillus sp ., D-10 Bacillus sp., D 1-1 Bacillus sp., Fa 4-1 B. subtilis , Fa 4-2 Bacillus sp., Far 8 Bacillus sp., 5-3 Bacillus sp., D 7-1 B. subtilis , D 7-3 Bacillus sp., D 1-3 Bacillus sp., Fz 9 Bacillus sp., 3-1 Bacillus sp., Б-2 B. circulans , Б-4 B. circulans , Б (2-1) B. lichen-iformis , Б-5 B. licheniformis , Б-12 B. licheni-formis , 11-1 Bacillus sp., 11-3 Bacillus sp., 32 Bacillus sp., 3-3 Bacillus sp., 1а B. poly-myxa , Р-9 B. polymyxa , Р-8 B. polymyxa , К11 B. subtilis , ВВ(С) B. subtilis , 5Б-1 B. subtilis ), а также девять штаммов рода Pseudomonas (Sgc-1 Pseudomonas sp., 12-2
Pseudomonas sp., 13-2 Pseudomonas sp., 151 Pseudomonas sp., Sgrc-1 P. fluorescens , 143 P. chlororaphis , 14-4 P. chlororaphis , 16-2 Pseudomonas sp., Oif 2-1 Pseudomonas sp.) и семена рапса озимого сорта Лорис.
Оценку антагонистической активности штаммов in vitro к агрессивному изоляту возбудителя фомоза рапса озимого проводили методом двойных или встречных культур [11]. Метод позволяет определить антагонистическую активность выделенных или коллекционных штаммов при совместном культивировании с возбудителем болезни (агрессивным изолятом) в одной чашке Петри.
Культуры бактериальных антагонистов и возбудителя болезни выращивали отдельно в течение пяти – семи суток на ага-ризированной питательной среде. Стерильным сверлом размером 0,7 см вырезали блоки с мицелием антагониста и патогена и помещали в одну чашку Петри на расстоянии 6,0 см. Контролем служили культуры антагонистов и патогена, посеянные порознь.
Антагонистов и патоген выращивали на картофельно-сахарозном агаре (КСА) [12] и на специализированных средах: для бактерий из рода Bacillus – Тайлона-3 [13], из рода Pseudomonas – Кинга В [14].
Учёты взаимодействия патогенов с антагонистами проводили на 20-е сутки культивирования, отмечали:
-
- рост патогена и антагониста в % от площади чашки Петри (конкуренция за площадь питания);
-
- наличие или отсутствие зон задержки роста патогена в результате синтеза антагонистами гидролитических ферментов или веществ антибиотической природы (стерильная зона);
-
- нарастание антагониста на колонию патогена (гиперпаразитическая зона).
Результаты и обсуждение. Установлена антагонистическая активность всех 36 испытанных коллекционных бактериальных штаммов in vitro к агрессивному изоляту возбудителя фомоза озимого рапса Phoma lingam на всех испытанных питательных средах. При этом выявлены три типа антагонистической активности: двойной (конкуренция за площадь питания и антибиоз), только конкуренция за площадь питания и только антибиоз. Нарастания бактериальных антагонистов на колонию патогена (гиперпаразитическая зона) не отмечено (табл. 1–4).
На среде КСА сформировался двойной тип антагонистической активности бактерий из рода Bacillus у восьми штаммов, из них у пяти (Fa 4-2, D-10, 11-2 Bacillus sp . , Б (2-1) B. licheniformis , D 7-1 B. subtilis ), при средней конкуренции за площадь питания (19,4–32,9 %) установлена более высокая антибиотическая активность при стерильной зоне 14,5–23,5 мм. Тогда как у трех штаммов (Б-5, Б-12 B. licheniformis и D 1-1 Bacillus sp.) выявлена высокая конкуренция за площадь питания (61,9–82,2 %) при небольшой стерильной зоне (4,0– 13,0 мм) (табл. 1, рис. 1).
У остальных 19 бактериальных штаммов из рода Bacillus установлена антибиотическая активность. При этом максимальная стерильная зона (26,0–37,5 мм) отмечена у восьми штаммов (11-3, 5-3, 3-3, Far 8, D 1-3, Fz 9, Bacillus sp . , 5Б-1 B. subtilis , и Б-4 B. circulans ).
На среде Тайлона-3 у бациллярных штаммов установлены три типа антибиотической активности: двойной (конкуренция за площадь питания и антибиоз), только конкуренция за площадь питания и только антибиоз (табл. 2, рис. 2).
При двойном типе антагонистической активности максимальный антибиоз (23,5–25,0 мм) отмечен у четырех бациллярных штаммов (K1-2 Bacillus sp., 01kopf Bacillus sp., 5Б-1 B. subtilis и Б-12 B. lichen-iformis ). Максимальная конкуренция за площадь питания (55,5–76,5 %) установлена у пяти штаммов (11-3, 5-3, Fa 4-2, 112 Bacillus sp. и Б-5 B. licheniformis ) . Один штамм (ВВ(С) B. subtilis ) проявил только конкуренцию за площадь питания (72,6 %). Антибиоз сформировался у шести штаммов, при этом максимальная стерильная зона отмечена у двух бациллярных штаммов (D 7-3 и 3-2 Bacillus sp.).
Таблица 1
Антагонистическая активность штаммов бактерий из рода Bacillus к возбудителю фо-моза рапса озимого Phoma lingam при температуре 25 °С на 20-е сутки культивирования на среде КСА
ФГБНУ ФНЦ ВНИИМК, 2024 г.

|
Штамм антагонист |
Площадь зарастания поверхности питательной среды, |
Размер стерильной зоны, мм |
|||
|
антагонистом |
патогеном |
||||
|
% |
см2 |
% |
см2 |
||
|
Phoma lingam |
- |
- |
37,2 |
23,8 |
- |
|
конкуренция за площадь питания + антибиоз |
|||||
|
Fa 4-2 Bacillus sp. |
23,7 |
15,1 ± 0,3 |
7,4 |
4,8 ± 0,9 |
23,5 |
|
Б (2-1) B. licheniformis |
32,9 |
20,9 ± 1,0 |
5,7 |
3,7 ± 1,5 |
23,0 |
|
D-10 Bacillus sp. |
19,4 |
12,4 ± 6,0 |
15,2 |
9,7 ± 3,5 |
19,5 |
|
D 7-1 B. subtilis |
28,1 |
18,0 ± 5,4 |
15,6 |
10,0 ± 0,6 |
15,0 |
|
11-2 Bacillus sp. |
30,0 |
19,2 ± 3,2 |
10,3 |
6,6 ± 2,2 |
14,5 |
|
Б-5 B. licheniformis |
61,9 |
39,5 ± 8,6 |
2,5 |
1,7 ± 1,3 |
13,0 |
|
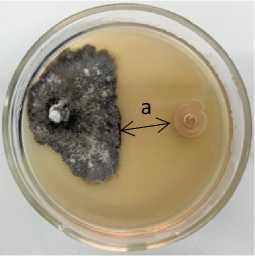
Б-12 B. licheniformis |
82,2 |
52,3 ± 3,8 |
1,2 |
0,8 ± 0,1 |
7,5 |
|
D 1-1 Bacillus sp. |
70,7 |
45,0 ± 7,2 |
5,4 |
3,5 ± 2,1 |
4,0 |
|
антибиоз |
|||||
|
11-3 Bacillus sp . |
3,2 |
2,1 ± 0,4 |
5,3 |
3,4 ± 0,8 |
37,5 |
|
5-3 Bacillus sp. |
4,4 |
2,8 ± 0,1 |
21,0 |
14,6 ± 1,8 |
32,0 |
|
3-3 Bacillus sp. |
6,7 |
4,3 ± 0,3 |
23,0 |
14,7 ± 1,0 |
30,5 |
|
Far 8 Bacillus sp. |
5,9 |
3,8 ± 2,9 |
20,7 |
13,3 ± 2,9 |
30,0 |
|
5Б-1 B. subtilis |
4,9 |
3,1 ± 0,3 |
14,6 |
9,3 ± 0,1 |
29,5 |
|
D 1-3 Bacillus sp. |
4,8 |
3,1 ± 0,1 |
22,2 |
14,2 ± 4,2 |
29,5 |
|
Fz 9 Bacillus sp. |
4,1 |
2,6 ± 0,3 |
16,8 |
10,8 ± 0,8 |
27,5 |
|
Б-4 B. circulans |
11,1 |
7,1 ± 1,2 |
10,4 |
6,7 ± 0,9 |
26,0 |
|
3-2 Bacillus sp. |
4,3 |
2,7 ± 1,9 |
29,8 |
19,0 ± 5,0 |
25,5 |
|
1а B. polymyxa |
5,6 |
3,6 ± 0,6 |
14,8 |
9,5 ± 1,5 |
24,5 |
|
P-9 B. polymyxa |
8,2 |
5,3 ± 2,1 |
26,4 |
16,9 ± 5,1 |
23,5 |
|
01kopf Bacillus sp. |
13,3 |
8,5 ± 3,7 |
20,5 |
13,0 ± 0,4 |
23,0 |
|
11-1 Bacillus sp. |
15,5 |
10,0 ± 3,0 |
6,7 |
4,3 ± 0,1 |
23,5 |
|
K1-2 Bacillus sp. |
17,5 |
11,2 ± 0,9 |
21,9 |
14,0 ± 2,0 |
21,5 |
|
Fa 4-1 B. subtilis |
13,9 |
8,9 ± 2,7 |
18,5 |
11,8 ± 0,8 |
21,0 |
|
D 7-3 Bacillus sp. |
2,8 |
1,8 ± 0 |
33,9 |
21,6 ± 0 |
21,0 |
|
K 1-1 B. subtilis |
9,9 |
6,4 ± 0,5 |
35,7 |
22,8 ± 2,8 |
19,5 |
|
ВВ(С) B. subtilis |
19,4 |
12,4 ± 5,0 |
19,8 |
12,6 ± 1,0 |
15,0 |
|
Б-2 B. circulans |
17,0 |
10,8 ± 5,5 |
38,3 |
24,4 ± 1,4 |
9,5 |
Phoma lingam + Б-12 B. licheniformis Phoma lingam + Fz 9 Bacillus sp.
Рисунок 1 – Антагонистическая активность бактериальных штаммов из рода Bacillu s к возбудителю фомоза рапса озимого Phoma lingam через 20 суток совместного культивирования на среде КСА при температуре 25 °С:
а – стерильная зона; 1 – конкуренция за площадь питания и антибиоз;
2 – антибиоз (ориг.) (ФГБНУ ФНЦ ВНИИМК, 2024 г.)
Таблица 2
Антагонистическая активность штаммов бактерий из рода Bacillus к возбудителю фо-моза рапса озимого Phoma lingam при температуре 25 °С на 20-е сутки культивирования на среде Тайлона-3
ФГБНУ ФНЦ ВНИИМК, 2024 г.
|
Штамм антагонист |
Площадь зарастания поверхности питательной среды |
Размер стерильной зоны, мм |
|||
|
антагонистом |
патогеном |
||||
|
% |
см2 |
% |
см2 |
||
|
Phoma lingam |
- |
- |
16,3 |
10,4 |
- |
|
конкуренция за площадь питания + антибиоз |
|||||
|
K1-2 Bacillus sp. |
41,3 |
26,2 ± 8,6 |
8,9 |
5,8 ± 3,1 |
25,0 |
|
01kopf Bacillus sp. |
35,7 |
22,7 ± 5,4 |
9,7 |
6,2 ± 2,1 |
25,0 |
|
5Б-1 B. subtilis |
21,9 |
13,9 ± 0,4 |
17,1 |
10,9 ± 2,8 |
24,0 |
|
Б-12 B. licheniformis |
41,4 |
26,3 ± 3,7 |
17,1 |
10,9 ± 2,6 |
23,5 |
|
Б-4 B. circulans |
21,0 |
13,3 ± 0,7 |
15,1 |
9,6 ± 1,0 |
21,5 |
|
Far 8 Bacillus sp. |
33,9 |
21,6 ± 4,2 |
23,1 |
14,7 ± 2,3 |
18,5 |
|
Fz 9 Bacillus sp. |
21,9 |
13,9 ± 6,3 |
16,4 |
10,5 ± 5,5 |
18,5 |
|
3-3 Bacillus sp. |
41,0 |
26,1 ± 8,0 |
11,0 |
7,0 ± 0,8 |
16,5 |
|
P-9 B. polymyxa |
36,2 |
23,0 ± 6,0 |
17,7 |
11,2 ± 1,8 |
16,5 |
|
K 1-1 B. subtilis |
35,3 |
22,4 ± 7,3 |
19,2 |
12,2 ± 3,5 |
16,5 |
|
D-10 Bacillus sp. |
36,8 |
23,5 ± 1,6 |
16,2 |
10,3 ± 3,5 |
14,5 |
|
11-1 Bacillus sp. |
27,5 |
17,5 ± 4,8 |
10,4 |
6,7 ± 1,3 |
14,5 |
|
D 1-1 Bacillus sp. |
46,3 |
29,5 ± 10,1 |
10,7 |
6,9 ± 0,2 |
13,5 |
|
D 1-3 Bacillus sp. |
47,1 |
29,9 ± 8,6 |
1,7 |
1,1 ± 0,3 |
8,5 |
|
Б (2-1) B. licheniformis |
47,6 |
30,2 ± 3,6 |
0,8 |
0,5 ± 0,1 |
8,0 |
|
11-3 Bacillus sp. |
55,6 |
35,4 ± 5,9 |
3,9 |
2,5 ± 2,1 |
11,5 |
|
5-3 Bacillus sp. |
56,5 |
36,0 ± 8,8 |
10,8 |
6,9 ± 3,5 |
16,5 |
|
Fa 4-2 Bacillus sp. |
64,4 |
41,1 ± 2,2 |
3,8 |
2,5 ± 0,9 |
6,5 |
|
11-2 Bacillus sp. |
71,7 |
45,7 ± 4,0 |
1,1 |
0,7 ± 0,5 |
8,0 |
|
Б-5 B. licheniformis |
76,5 |
48,6 ± 8,3 |
1,0 |
0,7 ± 0,2 |
8,5 |
|
конкуренция за площадь питания |
|||||
|
ВВ(С) B. subtilis |
72,6 |
46,2 ± 6,0 |
23,2 |
14,8 ± 3,6 |
0 |
|
антибиоз |
|||||
|
D 7-3 Bacillus sp. |
0,7 |
0,5 ± 0,1 |
29,1 |
18,6 ± 1,2 |
32,0 |
|
3-2 Bacillus sp. |
10,7 |
6,8 ± 3,9 |
35,9 |
22,8 ± 5,4 |
28,0 |
|
D 7-1 B. subtilis |
14,8 |
9,4 ± 0,1 |
36,3 |
23,1 ± 0,4 |
19,0 |
|
Fa 4-1 B. subtilis |
27,6 |
17,6 ± 2,1 |
31,6 |
20,1 ± 6,0 |
15,5 |
|
Б-2 B. circulans |
12,1 |
7,7 ± 0,4 |
28,2 |
17,9 ± 2,1 |
14,0 |
|
1а B. polymyxa |
24,6 |
15,6 ± 7,6 |
28,2 |
18,0 ± 7,4 |
1,0 |
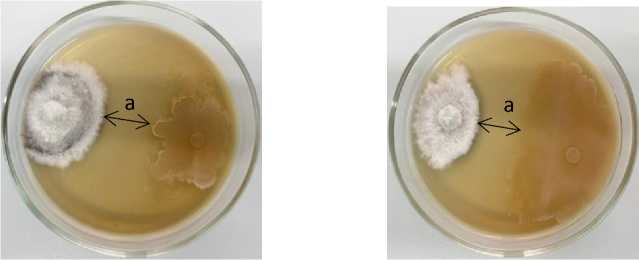
1 2
Phoma lingam + 5Б-1 B. subtilis Phoma lingam + 5-3 Bacillus sp.
Рисунок 2 – Антагонистическая активность бактериальных штаммов из рода Bacillu s к возбудителю фомоза рапса озимого Phoma lingam через 20 суток совместного культивирования на среде Тайлона-3 при температуре 25 °С: а – стерильная зона; 1, 2 – конкуренция за площадь питания и антибиоз (ориг.) (ФГБНУ ФНЦ ВНИИМК, 2024 г.)
Таблица 3
Антагонистическая активность штаммов бактерий антагонистов из рода Pseudomonas к возбудителю фомоза рапса озимого Phoma lingam при температуре 25 °С на 20-е сутки культивирования на среде КСА
ФГБНУ ФНЦ ВНИИМК, 2024 г.
|
Штамм антагонист |
Площадь зарастания поверхности питательной среды, % |
Размер стерильной зоны, мм |
|||
|
антагонистом |
патогеном |
||||
|
% |
см2 |
% |
см2 |
||
|
Phoma lingam |
- |
- |
37,2 |
23,8 |
- |
|
конкуренция за площадь питания + антибиоз |
|||||
|
12-2 Pseudomonas sp. |
26,3 |
16,8 ± 2,9 |
13,5 |
8,7 ± 6,1 |
12,5 |
|
15-1 Pseudomonas sp. |
83,7 |
53,3 ± 3,5 |
1,4 |
0,9 ± 0,1 |
6,0 |
|
антибиоз |
|||||
|
13-2 Pseudomonas sp. |
7,8 |
5,0 ± 0,4 |
25,4 |
16,2 ± 2,7 |
27,5 |
|
Sgc-1 Pseudomonas sp. |
6,3 |
4,1 ± 0,8 |
24,7 |
15,8 ± 0,6 |
23,5 |
|
16-2 Pseudomonas sp. |
23,6 |
15,1 ± 0,1 |
27,2 |
17,3 ± 6,6 |
15,5 |
|
Oif 2-1 Pseudomonas sp. |
9,1 |
5,8 ± 1,6 |
20,6 |
13,1 ± 2,2 |
15,0 |
|
14-4 Pseudomonas sp. |
15,8 |
10,1 ± 0,1 |
36,9 |
23,5 ± 11,9 |
11,5 |
|
14-3 P. chlororaphis |
13,7 |
8,7 ± 1,7 |
25,7 |
16,4 ± 6,4 |
10,5 |
|
Sgrc-1 P. fluorescens |
10,8 |
6,9 ± 1,5 |
35,9 |
22,9 ± 0,2 |
9,0 |
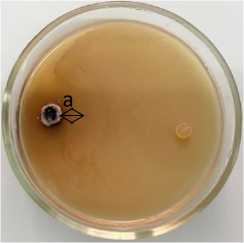
Phoma lingam + 15-1 Pseudomonas sp.
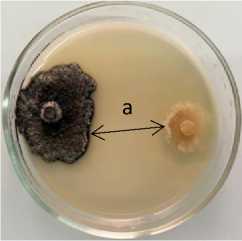
Phoma lingam + 13-2 Pseudomonas sp.
Рисунок 3 – Антагонистическая активность бактериальных штаммов из рода Pseudomonas к возбудителю фомоза рапса озимого Phoma lingam через 20 суток совместного культивирования на среде КСА при температуре 25 °С:
а – стерильная зона; 1 – конкуренция за площадь питания и антибиоз;
2 – антибиоз (ориг.) (ФГБНУ ФНЦ ВНИИМК, 2024 г.)
Таблица 4
Антагонистическая активность штаммов бактерий антагонистов из рода Pseudomonas к возбудителю фомоза рапса озимого Phoma lingam при температуре 25 °С на 20-е сутки культивирования на среде Кинга B
ФГБНУ ФНЦ ВНИИМК, 2024 г.
|
Штамм антагонист |
Площадь зарастания поверхности питательной среды, % |
Размер стерильной зоны, мм |
|||
|
антагонистом |
патогеном |
||||
|
% |
см2 |
% |
см2 |
||
|
Phoma lingam |
- |
- |
29,7 |
18,8 ± 0,8 |
- |
|
конкуренция за площадь питания + антибиоз |
|||||
|
13-2 Pseudomonas sp. |
26,5 |
16,9 ± 2,5 |
16,3 |
10,4 ± 6,9 |
16,5 |
|
12-2 Pseudomonas sp. |
53,9 |
34,3 ± 5,6 |
4,5 |
2,9 ± 1,7 |
13,0 |
|
антибиоз |
|||||
|
14-4 Pseudomonas sp. |
9,4 |
6,1 ± 1,1 |
27,6 |
17,5 ± 3,4 |
26,0 |
|
16-2 Pseudomonas sp. |
6,4 |
4,2 ± 0,2 |
22,5 |
14,4 ± 2,4 |
26,0 |
|
Oif 2-1 Pseudomonas sp. |
10,1 |
6,5 ± 1,2 |
26,6 |
17,0 ± 0,3 |
24,0 |
|
14-3 P. chlororaphis |
10,2 |
6,5 ± 1,6 |
30,4 |
19,5 ± 1,5 |
21,0 |
|
15-1 Pseudomonas sp. |
11,0 |
7,1 ± 1,2 |
28,8 |
18,4 ± 2,1 |
19,5 |
|
Sgc-1 Pseudomonas sp. |
18,0 |
11,5 ± 7,2 |
25,0 |
16,0 ± 1,1 |
15,0 |
|
Sgrc-1 P. fluorescens |
5,1 |
3,3 ± 1,3 |
36,1 |
24,6 ± 8,4 |
1,5 |
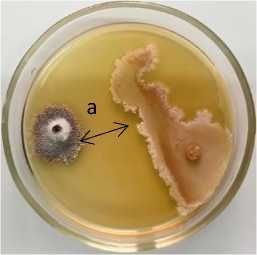
Phoma lingam + 12-2 Pseudomonas sp.
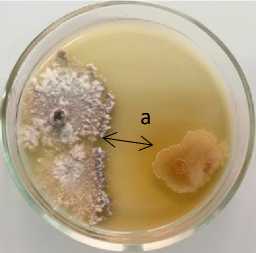
Phoma lingam + 14-3 P. chlororaphis
Рисунок 4 – Антагонистическая активность бактериальных штаммов из рода Pseudomonas к возбудителю фомоза рапса озимого Phoma lingam через 20 суток совместного культивирования на среде Кинга В при температуре 25 °С:
а – стерильная зона; 1 – конкуренция за площадь питания и антибиоз;
2 – антибиоз (ориг.) (ФГБНУ ФНЦ ВНИИМК, 2024 г.)
Таким образом, на двух питательных средах КСА и Тайлона-3 из 27 коллекционных штаммов бактерий из рода Bacillus выделен 21 перспективный штамм, показавший максимальную антагонистическую активность с разными типами действия, для проведения вторичного скрининга на фоне искусственного заражения в лабораторных условиях во влажной камере: Fa 4-2, D-10, 11-2, D 1-1, 11-3, 5-3, 33, 3-2, Far 8, D 1-3, Fz 9, D 7-3, K 1-2, 01kopf Bacillus sp., Б (2-1), Б-5, Б-12 B. li-cheniformis , 5Б-1, D 7-1, ВВ(С) B. subtilis и Б-4 B. circulans .
У бактерий из рода Pseudomonas на среде КСА установлен двойной тип антагонистической активности только у двух штаммов, при этом у штамма 12-2 Pseudomonas sp. отмечена средняя конкуренция за площадь питания (26,3 %) при более высокой антибиотической активности (стерильная зона составила 12,5 мм). Тогда как у штамма 15-1 Pseudomonas sp. установлена высокая конкуренция за площадь питания (83,7 %) при небольшой стерильной зоне (6,0 мм) (табл. 3, рис. 3).
У семи остальных штаммов выявлен тип антагонистической активности антибиоз, при этом максимальная стерильная зона (15,0–27,5 мм) отмечена у четырех штаммов (13-2, Sgc-1, 16-2, Oif 2-1 Pseudomonas sp.).
На специализированной среде Кинга В двойной тип антагонистической активности установлен так же, как и на среде КСА, у двух штаммов: 13-2 и 12-2 Pseudomonas sp. При этом у штамма 13-2 Pseudomonas sp. установлена средняя конкуренция за площадь питания (26,5 %) при более высокой антибиотической активности (стерильная зона – 16,5 мм). Тогда как у штамма 12-2 Pseudomonas sp. установлена более высокая конкуренция за площадь питания (53,9 %) при средней стерильной зоне (13,0 мм) (табл. 4, рис. 4).
У остальных семи штаммов установлен один тип антагонистической активности – антибиоз, с максимальной стерильной зоной (21,0–26,0 мм) у четырех штаммов (14-4, 16-2, Oif 2-1 и 14-3 Pseudomonas sp.).
Таким образом, на двух питательных средах КСА и Кинга В из девяти коллекционных штаммов бактерий рода Pseudomonas выделены восемь перспективных штаммов, показавших максимальную антагонистическую активность с разными типами действия против возбудителя фо-моза: 12-2, 15-1, 13-2, Sgc-1, 16-2, Oif 2-1, 14-4 Pseudomonas sp. и 14-3 P. chlorora-phis.
Заключение. В 2024 г. в результате первичного скрининга бактериальных штаммов антагонистов из рабочей коллекции лаборатории биометода агротехнологиче-ского отдела ФГБНУ ФНЦ ВНИИМК для проведения вторичного скрининга на фоне искусственного заражения возбудителем фомоза рапса озимого выделены перспективные штаммы бактерий с разными типами антагонистической активности: 21 штамм бактерий рода Bacillus : Fa 4-2, D-10, 11-2, D 1-1, 11-3, 5-3, 3-3, 3-2, Far 8, D 1-3, Fz 9, D 7-3, K1-2, 01kopf Bacillus sp., Б (2-1), Б-5, Б-12 B. licheniformis , 5Б-1, D 71, ВВ(С) B. subtilis, Б-4 B. circulans и восемь штаммов рода Pseudomonas : 12-2, 151, 13-2, Sgc-1, 16-2, Oif 2-1, 14-4 Pseudomonas sp. и 14-3 P. chlororaphis.
Список литературы Поиск антагонистов возбудителя фомоза рапса озимого: первичный скрининг бактериальных штаммов из рабочей коллекции лаборатории биометода ФГБНУ ФНЦ ВНИИМК
- Index Fungorum Data bases: [Электронный ресурс]. - Режим доступа: https://www.indexfun-gorum. org/names/NamesRecord. asp?RecordID=2301 54 (дата обращения: 01.10.2024).
- Пивень В.Т., Сердюк О.А. Фитосанитарный мониторинг болезней рапса // Масличные культуры. Науч.-тех. бюл. ВНИИМК. - 2011. - Вып. 2 (148-149). - С. 162-166.
- Бочкарев Н.И., Пивень В.Т., Тишков Н.М. [и др.]. Защита рапса // Защита и карантин растений. -2017. - № S1. - С. 37-76.
- Сердюк О.А., Трубина В.С., Горлова Л.А. Влияние внутренней инфекции на всхожесть и масличность семян масличных культур семейства капустные // Масличные культуры. - 2019. - Вып. 3 (179). -С. 119-123.
- Гасич Е.Д. Фомоз рапса (Обзор литературы) // Вестник защиты растений. - 2004. - № 1. - С. 1124.
- Gomzhina M.M., Gasich E.L. Plenodomus species infecting oilseed rape in Russia // Plant Protection News. - 2022. - Vol. 105 (3). - P. 135-147.
- Костин Н.К., Кузнецова А.А., Дудченко И.П. [и др.]. Видовой состав микромицетов, ассоциированных с растениями рапса озимого некоторых регионов России // Сб. мат-лов конф. «Генетические технологии в микробиологии и микробное разнообразие», Пущино, 06-08 декабря 2022 г. - М.: ГЕОС, 2022. - С. 56-59.
- Сердюк О.А. Сравнительная оценка эффективности препаратов из группы триазолов против склеротиниоза и фомоза на рапсе озимом // Защита и карантин растений. - 2012. - № 5. - С. 21-22.
- Девяткина Т.Ф., Чигорин С.С., СилаевА.И. [и др.]. Оценка эффективности фунгицидов в сдерживании альтернариоза и фомоза на яровом рапсе // Аграрный научный журнал. - 2024. - № 5. - С. 1927.
- Маслиенко Л.В. Лаборатория биологических средств защиты растений (вчера, сегодня, завтра) // История научных исследований во ВНИИМКе за 90 лет. - Краснодар, 2002. - С. 191-197.
- Егоров Н.С. Выделение микробов антагонистов и биологические методы учета их антибиотической активности. - М.: Изд-во Моск. ун-та, 1957. - 78 с.
- Лысак В.В., Желдакова Р.А., Фомина О.В. Микробиология. Практикум: пособие. - Минск: БГУ 2015. - 115 с.
- Егоров Н.С. Основы учения об антибиотиках. - М.: Наука, 2004. - 528 с.
- King E.D., Ward M.K. and Raney D.E. Two simple media for the demonstration of pyocyanin and fluorescin // Journal of laboratory and clinical medicine. - 1954. - V 44. - P. 301-307.
