Показатели роста и устойчивости к дефициту кислорода у колеоптилей Oryza sativa L. из коллекции Федерального научного центра риса

Автор: Богданова Е.М., Бертова А.Д., Кирпичникова А.А., Бикташева М.О., Кондратьева А.В., Шапиро А.С., Пузанский Р.К., Коротенко Т.Л., Мухина Ж.М., Емельянов В.В., Шишова М.Ф.
Журнал: Сельскохозяйственная биология @agrobiology
Рубрика: Зерновые культуры устойчивость и адаптация
Статья в выпуске: 3 т.58, 2023 года.
Бесплатный доступ
На этапе прорастания злаков первым развивается колеоптиль - ювенильный орган, выполняющий важную функцию защиты настоящего листа. Механизмы регуляции роста и развития колеоптиля во многом не исследованы. Особого внимания заслуживает его формирование у проростков риса, способных развиваться в условиях недостатка кислорода. В представленной работе впервые продемонстрировано значение скорости роста и жизнеспособности колеоптилей проростков риса при затоплении в определении устойчивости растений и их развития в последующем онтогенезе. Цель работы заключалась в оценке скорости роста и жизнеспособности колеоптилей в условиях затопления у образцов риса ( Oryza sativa L.) из коллекции Федерального научного центра риса (ФНЦ риса). В работе использовали семена 36 сортов и форм, в том числе 24 сорта отечественной селекции (Аметист, Анаит, Велес, Виола, Вита, Гамма, Жемчуг, Красноармейский 313, Кубань 3, Лидер, Наташа, Новатор, Олимп, Привольный 4, Рапан, Регул, Соната, Спринт, Титан, Уссур, Фонтан, Хазар, Шарм, Южная ночь), один сорт из Узбекистана (Девзра), три сорта из Китая (Xiannui, Zhongyon, Zhongyon 207) и восемь образцов из Международного института риса (International Rice Research Institute, Филиппины) (AA WAB 56-125, HHZ11 Y6-Y2-SUB1, HHZ8 SAL 14 SUB1, HHZ9 DT12 SUB1, IR 50, IR14 L 110, Kirkpinar, ПВ-1 IRBLSH). Первичное тестирование интенсивности роста растений проводили в полевом опыте в коллекционном питомнике (оросительная система ФНЦ риса). Для серии лабораторных исследований семена поверхностно стерилизовали и замачивали в горячей воде (55 °С) на 1 ч. Далее по 50 семян контрольных образцов помещали в эмалированный поддон на стеклянные мостки, покрытые марлей. В поддон наливали 4 % питательный раствор Кнопа, накрывали стеклом и проращивали в условиях нормального доступа воздуха. Семена опытных вариантов для создания гипоксии помещали в емкости объемом 750 мл и заливали тем же раствором под самый верх; емкости герметично закрывали крышкой. В обоих вариантах растения выращивали в течение 4 сут при 29 °С в темноте. Для измерения длины колеоптилей проростки раскладывали в чашки Петри, сканировали с помощью HP ScanJet G2710 («Hewlett-Packard», США) и оцифровывали изображения в программе ImageJ (версия 1.8.0_172; https://imagej.nih.gov/ij/download.html). После сканирования по 5 колеоптилей из контрольных и опытных вариантов использовали для определения жизнеспособности в тесте по восстановлению солей тетразолия. По результатам опытов к быстрорастущим были отнесены сортам Спринт и Кубань 3, длина колеоптилей у которых достигала 19-25 мм как при нормоксии, так и при гипоксии. В группу медленнорастущих вошли три сорта филиппинской селекции, несущие аллель SUB1A (HHZ11 Y6-Y2-SUB1, HHZ8 SAL 14 SUB1, HHZ9 DT12 SUB1), китайский сорт Xiannui и отечественные сорта Аметист, Жемчуг, Наташа, Рапан и Южная ночь. Для этой группы было характерно подавление роста колеоптилей под действием гипоксии в 2,5-3,0 раза. Нужно отметить наличие положительной корреляции между длиной колеоптиля при нормоксии и гипоксии (r = 0,70, p = 10-6), то есть формы, активно растущие в аэробной среде, быстро росли и при затоплении. Последующий детальный анализ роста колеоптилей в условиях гипоксии показал, что изменения роста соответствовали нескольким паттернам. Наиболее распространенной реакцией на гипоксию колеоптилей риса (I паттерн) было подавление роста с сохранением небольшой части растений, продолжавших рост. Именно в эту группу вошли все филиппинские сорта - носители аллеля SUB1A . В другой группе (II паттерн) длина колеоптилей под действием гипоксии превышала таковую при нормоксии. Примерно половина растений, использующих III паттерн, замедляла рост, а другая часть продолжала расти как при нормоксии. Растения, использующие IV паттерн, показали наименьшие изменения распределения. Только у сорта Спринт наблюдались черты стратегии избегания (LOES, low oxygen escape syndrome), связанные с усилением роста, хотя оно было несильным. Уникальным V паттерном обладал сорт Южная ночь, у которого колеоптили проростков медленно росли как в контроле, так и при затоплении. В условиях гипоксии у всех протестированных форм жизнеспособность значительно снижалась. У наиболее быстрорастущих сортов (Спринт, Кубань 3) интенсивность окрашивания при использовании солей тетразолия была выше как в контроле (в 6 раз), так и в опыте (в 2 раза) по сравнению с медленнорастущими формами (Аметист, Южная ночь, филиппинские SUB1A -сорта). Полученные данные позволяют заключить, что удлинение колеоптиля коррелировало с интенсивностью метаболизма и устойчивостью к затоплению, а следовательно, может быть использовано в качестве критерия для оценки реакции сортов и форм риса на недостаток кислорода.
Oryza sativa, рис, затопление, гипоксия, колеоптиль, рост, устойчивость
Короткий адрес: https://sciup.org/142238895
IDR: 142238895 | УДК: 633.18:581.1:57.04 | DOI: 10.15389/agrobiology.2023.3.538rus
Growth and viability of coleoptiles under oxygen deficiency in Oryza sativa L. from the collection of the Federal Rice Research Center
The distinctive ability of rice seedlings lies in the ability to germinate under conditions of oxygen lack. At the germination stage, the first to develop is the coleoptile, a juvenile organ that protects the true leaf in cereals. The mechanisms of regulation of growth and development of this organ have not been studied to a large extent. Special attention may be paid to a coleoptile in seedlings capable of germinating under oxygen conditions. In the presented study, for the first time, the importance of the growth rate and viability of coleoptiles of rice seedlings during flooding in determining survival and development was demonstrated. A total of 36 varieties and forms from the collection of the Federal Research Center for Rice, Krasnodar, were tested. Sprint and Kuban 3 were among the fastest growing varieties. Their coleoptiles reached 19-25 mm length both under normoxia and hypoxia. The slowest growing group included three Philippine varieties harbouring the SUB1A allele (HHZ11 Y6-Y2-SUB1, HHZ8 SAL 14 SUB1, HHZ9 DT12 SUB1), Chinese variety Xiannui and domestic varieties Amethyst, Zhemchug, Natasha, Rapan and Yuzhnaya noch. This group of varieties was characterized by inhibition of the growth of coleoptiles by 2.5-3 times under submergence. In main a positive correlation was estimated between the coleoptile length in normoxia and hypoxia (r = 0.70, p = 10-6), i.e. forms that actively grow in an aerobic environment also grow rapidly when flooded. Further detailed analysis of the growth of coleoptiles under hypoxic conditions showed that growth changes correspond to several patterns. The most common reaction to hypoxia in rice coleoptiles of the first group was growth suppression, but with the preservation of a small part of the plants that continue to grow. This group included all Philippine cultivars harbouring the SUB1A allele. In the second group, a significant proportion was plants which length under submergence exceeded that at normoxia. Approximately half of the plants of the third group slowed growth arrest, while the other part continued to grow as in normoxia. The fourth group included the variety Sprint, which was the only one to have traits of avoidance strategy (LOES - low oxygen escape syndrome) associated with growth enhancement, although this enhancement was not intensive. The variety Yuzhnaya noch had a unique growth pattern, the coleoptiles of which grew slowly both in normoxia and being submerged. In addition to growth, the viability of coleoptiles was analyzed in the work, which was assessed using a tetrazolium test. Under hypoxic conditions, the viability of all tested forms significantly decreased. In the fastest growing varieties (Sprint, Kuban 3), the color intensity of tetrazolium salts was higher both in the control (6-fold) and in the experiment (2-fold), compared to slow growing forms (Amethyst, Yuzhnaya noch, Philippine SUB1A varieties). In general, growth rate correlates with metabolic rate and submergence tolerance. Obtained results illustrate the tolerance to oxygen deficiency of the genotypes from the collection of the Federal Research Center of Rice, and show that coleoptile elongation can be used as a criterion for assessing the tolerance of rice varieties to the lack of oxygen.
Текст научной статьи Показатели роста и устойчивости к дефициту кислорода у колеоптилей Oryza sativa L. из коллекции Федерального научного центра риса
Рис (Oryza sativa L.) — древнейшая сельскохозяйственная культура, возделывание которой началось более 5 тыс. лет до н.э. Она составляет основу питания нескольких миллиардов человек. В 2022 году было собрано 525,6 млн т зерна риса, который выращивается на более чем 167 млн га . Достаточно интенсивно рис возделывается в России. Посевные площади под рис на июнь 2022 года в Российской Федерации составили 162,5 тыс. га . Урожай риса в главном рисосеющем регионе Российской Федерации — на Кубани (92,3 тыс. га), по данным Росстата, в 2022 году составил 582,6 тыс. т . Расшифровка механизмов адаптации растений риса к затоплению может быть использована для получения новых, устойчивых к длительному затоплению сортов для снижения количества пестицидов в рисовых чеках и экологизации отрасли рисоводства.
Отличительная особенность риса заключается в способности прорастать в условиях недостатка (гипоксия) или полного отсутствия кислорода (аноксия). Растения, произрастающие в водно-болотной среде, в том числе рис, используют две стратегии адаптации к дефициту кислорода. Первая стратегия направлена на активное избегание кислородной недостаточности (low oxygen escape syndrome, LOES), вторая — пассивная стратегия покоя (low oxygen quiescence syndrome, LOQS) (1).
При развитии LOES-стратегии наблюдаются быстрое удлинение побегов, гипонастический изгиб и изменение анатомии листьев для улучшения диффузии газов, формирование аэренхимы, увеличение числа поверхностных придаточных корней (2-6). Образование аэренхимы, необходимое для интенсификации газообмена между органами и тканями растения, регулируется фитогормонами, в первую очередь этиленом. Именно он рассматривается в качестве ключевого регулятора гормонального статуса растения в процессе развития LOES-стратегии: этилен инициирует понижение содержания абсцизовой кислоты и способствует повышению концентрации ауксина и гиббереллинов (3, 5, 7, 8). У риса LOES-стратегия регулируется генами SNORKEL1 ( SK1 ) и SNORKEL2 ( SK2 ), которые относятся к семейству ERF-VII (ethylene response factor). При повышении их экспрессии в междоузлиях не только увеличивается содержание гиббереллинов, но и повышается чувствительность к ним, что стимулирует активность вставочных меристем (9).
Важная адаптация к дефициту кислорода — образование газовой пленки на гидрофобной поверхности листьев и стеблей затопленных растений. Известно, что растения риса удерживают поверхностную газовую пленку даже при длительном (4-5 сут) затоплении (10). Она отвечает за доставку кислорода с надводной поверхности и важна для осуществления подводного фотосинтеза, обеспечивающего растения энергией (5, 6). Ускоренный рост происходит и за счет метаболизации сахаров, которая осуществляется при участии протеинкиназ. Например, CIPK15 (calcineurin B-like proteininteracting protein kinase15) инициирует SnRK1A-зависимый каскад анаэробного расщепления крахмала (11). В свою очередь, SnRK1A (sucrose-nonfermenting1-related protein kinase 1A), взаимодействуя с фактором транскрипции MYBS1, стимулирует экспрессию гена α-амилазы, особенно важной для расщепления крахмала при прорастании семян.
Торможение роста и обмена веществ — основные критерии альтернативной LOQS стратегии. Этой стратегии сопутствуют метаболические адаптации, препятствующие энергетическому голоданию, закислению цитоплазмы и токсичности анаэробных метаболитов. Энергия, выработанная в процессе метаболизма крахмала, гликолиза и брожений, преимущественно тратится на процессы транспорта, защиту клеток от активных форм кислорода (АФК) и предотвращение денатурации белков (5, 11, 12). Главным регулятором LOQS у риса, как и в случае со стратегией избегания, служит этилен. Белок, кодируемый геном SUB1A и относящийся к группе этилен-чувствительных факторов транскрипции (ERF-VII), тормозит синтез этилена, приводя к снижению количества гиббереллинов. При этом стимулируется накопление брассиностероидов — регуляторов роста, способствующих распаду биологически активных гиббереллинов и аккумуляции белка SLENDER RICE1 (SLR1), который подавляет трансдукцию гиббереллинового сигнала (13). Наряду с этим SUB1A тормозит метаболизм сахаров, снижая экспрессию генов амилазы и сахарозосинтазы (14). При осуществлении стратегии LOQS во время затопления затормаживается рост растений, а сэкономленные ресурсы направляются на пережидание периода недостатка кислорода и последующее отрастание после восстановления газообмена.
Колеоптиль — особый ювенильный орган проростков у злаков. Он прорастает сквозь почву, сохраняя интактными настоящие листья. Если свет попадает на колеоптиль, он прекращает рост и сквозь него прорастает настоящий лист. Колеоптиль имеет ограниченный период развития, когда все его клетки проходят этап роста растяжением. Считается, что экологически более длинные колеоптили во многих случаях обладают преимуществом, например обеспечивая защиту от высокой температуры и плотной среды (15, 16). Способность удлинять колеоптиль оказалась полезной для сельскохозяйственных культур в неблагоприятных условиях, поскольку глубокий посев важен для роста в более подходящих условиях температуры и влажности, снижает риск повреждения при нападении мышей или других животных, а также уменьшает вред от действия гербицидов до появления всходов (17-19).
Установлено значительное число факторов, регулирующих удлинение колеоптилей на транскрипционном и посттрансляционном уровнях (20, 21). Рост растяжением хорошо изучен на колеоптилях злаков, в частности овса, кукурузы и канареечника. Отметим, что у риса на этапе прорастания первым из зерновки развивается именно колеоптиль. Колеоптили этого растения заканчивают свой рост на 6-9-е сут. В условиях затопления рост колеоптилей сортов риса, придерживающихся LOES стратегии, значительно ускоряется, а у LOQS-форм, наоборот, затормаживается (22). В связи с этим целесообразно использовать длину колеоптиля как критерий при выборе генотипов, контрастных по способности к росту, а следовательно, по устойчивости.
В настоящее время в Федеральном научном центре риса (г. Краснодар) в Уникальной научной установке — УНУ Коллекция генетических ресурсов риса хранится 7,3 тыс. образцов. Генофонд риса в УНУ представлен сортами, сортообразцами, мутантами, дигаплоидами O. sativa двух подвидов — indica и japonica , включает 82 разновидности из 42 рисосеющих стран.
Сохраняемый генофонд всесторонне изучается для выявления источников и доноров ценных признаков (23, 24). Однако тестирование коллекционных образцов по устойчивости к различным неблагоприятным факторам окружающей среды (в том числе по отношению к недостатку кислорода), к сожалению, далеко от своего завершения. На наш взгляд, свойство растений риса менять стратегию адаптации в условиях затопления на ранних стадиях онтогенеза способно во многом определить конечную урожайность этой сельскохозяйственной культуры. Кроме того, не ясно, можно ли экстраполировать рост и устойчивость ювенильного органа на устойчивость и развитие взрослого растения.
В представленной работе впервые продемонстрировано значение скорости роста и жизнеспособности колеоптилей проростков риса при затоплении в определении устойчивости растений и их развития в последующем онтогенезе.
Цель работы заключалась в оценке скорости роста и жизнеспособности колеоптилей в условиях затопления у сортов и форм риса из коллекции Федерального научного центра риса.
Методика. Выращивание риса ( Oryza sativa L.) и фенотипирование растений по хозяйственно ценным признакам проводили в 2019-2022 годах в коллекционном питомнике (оросительная система ФНЦ риса, г. Краснодар, п/о Белозерное).
В работе использовали семена 36 сортов и форм из коллекции ФНЦ риса. В анализ были взяты 24 сорта риса отечественной селекции (Аметист, Анаит, Велес, Виола, Вита, Гамма, Жемчуг, Красноармейский 313, Кубань 3, Лидер, Наташа, Новатор, Олимп, Привольный 4, Рапан, Регул, Соната, Спринт, Титан, Уссур, Фонтан, Хазар, Шарм, Южная ночь), один сорт из Узбекистана (Девзра), три сорта из Китая (Xiannui, Zhongyon, Zhongyon 207) и восемь образцов из Международного института риса (International Rice Research Institute, IRRI, Филиппины) (AA WAB 56-125, HHZ11 Y6-Y2-SUB1, HHZ8 SAL 14 SUB1, HHZ9 DT12 SUB1, IR 50, IR14 L 110, Kirkpinar, ПВ-1 IRBLSH). Среди филиппинских образцов были три формы, несущие аллель SUB1A , характерный для наиболее устойчивых и медленнорастущих LOQS-сортов риса.
В рамках международного сотрудничества образцы риса мировой селекции поступали в коллекцию научного центра из разных унифицированных питомников сортоиспытания (IRRI, Филиппины), в которых применялись различные способы возделывания: IRLON — при укороченном затоплении, IRCTN — питомник холодостойких форм, IRTON — искусственно орошаемый рис умеренных широт, IRLYN-SUB — глубоководный, толерантный к затоплению рис, GAR-IRLL — рис орошаемых низменностей, питомник зеленых технологий; GSR-Rell — поливной, суперрис, питомник зеленых технологий, .
Скорость роста проростков определяли визуально по 9-балльной шкале, где 1 балл — низкая, 3-5 баллов — средняя, 7-9 баллов — высокая. К сортам с высокой скоростью роста (9 баллов) на начальной стадии прорастания относили генотипы с длиной проростка на 7-е сут более 2 см; 7 баллов — 1,5-2,0 см, 3-5 баллов — 1,0-1,5 см, 1 балл — менее 1 см. В результате сорта и сортоформы риса разделили на три группы по интенсивности роста (низкая, средняя и высокая) на ранней стадии онтогенеза. Далее оценивали развитие растений в условиях нормоксии: фиксировали общую высоту, длину метелки, длительность периода развития до цветения и совокупного вегетационного периода (длительность вегетационного периода сортообразцов оценивали с даты всходов до цветения, а затем до полной спелости зерна). Определяли элементы продуктивности, включая массу 1000 зерен, и общую продуктивность метелки. Для биометрического анализа на делянках отбирали модельные снопы из 10 растений.
Для серии лабораторных исследований семена поверхностно стерилизовали 50 % раствором гипохлорита натрия в течение 15 мин, 10 раз промывали стерильной водой и замачивали в горячей воде (55 ° С) на 1 ч. Далее по 50 семян контрольных образцов помещали в эмалированный поддон на стеклянные мостки, покрытые марлей. В поддон наливали 4 % питательный раствор Кнопа (25) до уровня стеклянных мостков, накрывали стеклом и проращивали в условиях нормального доступа воздуха. Семена опытных вариантов для создания гипоксии помещали в емкости объемом 750 мл, заливали тем же раствором под самый верх (высота водяного столба 12 см) и герметично закрывали крышкой. В обоих вариантах растения выращивали в течение 4 сут при 29 ° С в темноте. На 4-е сут содержание кислорода в гипоксическом растворе составляло в среднем 0,46 мг/л. Его измеряли с помощью анализатора растворенного кислорода Эксперт-009 («Эконикс-Эксперт», Россия). Посуду, марлю и растворы для работы с растениями предварительно стерилизовали.
Для измерения длины колеоптилей проростки раскладывали в чашки Петри, сканировали с помощью HP ScanJet G2710 («Hewlett-Packard», США) и оцифровывали изображения в программе ImageJ (версия 1.8.0_172) . Для анализа использовали все проросшие растения из 50 посеянных. После сканирования по 5 колеоптилей из контрольных и опытных вариантов использовали для определения жизнеспособности в тесте по восстановлению солей тетразо-лия. Колеоптили помещали в пробирки с 5 мл раствора хлорида 2,3,5-три-фенилтетразолия (8 г/л), приготовленного на 0,1 М натрий-фосфатном буфере (pH 6,9). Далее в течение 15 мин раствор с отрезками растительной ткани подвергали вакуум-инфильтрации (-70 Па), после чего пробирки оставляли на ночь в термостате (29 °С).
На следующий день проводили экстракцию трифенилформазана (восстановленная форма трифенилтетразолия, окрашенная в красный цвет) из растительных тканей. Для этого колеоптили, 3-кратно промытые дистиллированной водой, заливали 1,5 мл 96 % этилового спирта в микропробирках объемом 2 мл и нагревали в течение 10 мин при 85 ° С в твердотельном термостате TDB-120 («BioSan», Латвия). Оптическую плотность измеряли при X = 485 нм на спектрофотометре SPECTROstar Nano («BMG LABTECH GmbH», Германия). Жизнеспособность оценивали в единицах оптической плотности А 485 на 1 проросток. Эксперименты проводили в трех биологических повторностях.
Статистический анализ осуществляли в среде языка R (26). Определяли средние значения показателей ( M ) и стандартные ошибки средних (±SEM). Проверку на нормальность проводили, используя тест Шапиро-Уилка. Большая часть выборок показала распределение длин, отличное от нормального, поэтому для их сравнения применяли непараметрический тест Манна-Уитни-Уилкоксона. Для сравнения окрашивания тетразолием использовали t -тест. При множественных сравнениях p-значения корректировали методом FDR (false discovery rate). Проверку принадлежность значений длин при гипоксии и нормоксии к одному распределению проводили 542
с применением теста Колмогорова-Смирнова, рассчитывали несходство (D) и p-значение (p-value). Ядерную оценку плотности случайной величины анализировали с использованием функции Епанечникова. Для определения корреляции вычисляли коэффициент Спирмена ( р ).
Результаты. По данным фенотипирования, развитие растений O. sa-tiva из коллекции ФНЦ риса, вошедших в эксперимент, не зависело от страны и питомника происхождения (табл. 1). Скорость роста оценивали на начальных этапах развития проростков. Именно в этот период растениям приходится преодолеть 10-сантиметровый слой воды, образующийся в результате залива чека после посева («укороченное затопление»).
Сорта риса Южная ночь, Наташа, Велес и HHZ8 SAL 14 SUB1, отнесенные к группе низких (низкая интенсивность роста), имели ограниченную высоту стебля (см. табл. 1). К группе высоких принадлежали высокорослые растения сортов Девзра, Kirkpinar и Кубань 3. Интересно, что к группе высоких можно было отнести и сорта, достигавшие лишь половины высоты высокорослых, — IR 50, Zhongyon и Zhongyon 207. Однако отметим, что это преимущественно касалось сортов иностранной селекции, которые отбирали по иным критериям. Кроме того, при различных температурных условиях устойчивость растений к затоплению в значительной степени варьирует (27). То есть скорость роста и устойчивость сортов, выведенных на Филиппинах или в Китае, может существенно меняться при выращивании в условиях Краснодарского края. В 2022 году в период посева (I декада мая) среднесуточная температура не превышала 16 -17 °С . Возможно, именно это стало причиной отсутствия четкой корреляции между скоростью роста на начальном этапе и последующим развитием растений риса (см. табл. 1).
К наиболее медленнорастущим относились три сорта филиппинской селекции, несущие аллель SUB1A , китайский сорт Xiannui и отечественные сорта Аметист, Жемчуг, Наташа, Рапан и Южная ночь. Длина колеоптилей у них составляла около 10 мм в контроле и около 3 мм при гипоксии, что означает подавление роста в 2,5-3,0 раза. Следует заметить, что из представленных в эксперименте пяти форм, имеющих окрашенные антоцианом зерновки (Виола, Вита, ПВ-1 IRBLSH, Наташа и Южная ночь), две последние оказались медленнорастущими. Самым медленнорастущим был сорт Южная ночь, у которого в контроле колеоптили вырастали до 4,0 мм, а в опыте — до 2,5 мм. Тем самым впервые для перечисленных сортов было показано замедление роста колеоптилей при недостатке кислорода. Однако интенсивность подавления в условиях затопления была согласована с нативной интенсивностью роста проростков риса в контрольных условиях.
Для большей части сортов и форм было характерно снижение медианной (внизу от диагонали, соответствующей равенству) длины колеоптиля при кислородном голодании (см. рис. 1), только у сортов Виола и Красноармейский 313 различия между нормоксией и гипоксией оказались недостоверны (табл. 2). В наибольшей степени рост в условиях затопления подавлялся у сортов HHZ11 Y6-Y2-SUB1, HHZ8 SAL 14 SUB1, IR14 L 110, IR 50, Xiannui, Аметист, Жемчуг, Наташа и Южная ночь. Три сорта (Спринт, Велес и Привольный 4) показали статистически значимое (p < 0,05) увеличение длины при гипоксии (см. рис. 1, табл. 2). Также нужно отметить наличие положительной корреляции между длиной колеоптиля при нор-моксии и гипоксии ( р = 0,70; p = 10 - 6), то есть формы, активно растущие в аэробной среде, быстро росли и при затоплении.
-
1. Характеристика сортов и сортообразцов Oryza sativa L. разного происхождения из УНУ Коллекция генетических ресурсов риса Федерального научного центра риса (г. Краснодар) при выращивании в условиях Краснодарского края (n = 10, N = 3, M±SEM, 2019-2022 годы)
Сорт, сортообразец
Страна происхождения
Год репродукции семян
Скорость роста, питомник, цвет зерна
Период, сут
Высота растения, см
Длина метелки, см
Число колосков на метелке, шт.
Продуктивность метелки, г
Масса 1000 зерен, г
до цветения]
вегетации
AA WAB 56-125
Филиппины
2021
В, IRCTN
84
128±3
93,0±3,5
16,9±0,4
104±6
2,2±0,2
30,2±0,2
HHZ11 Y6-Y2-
Филиппины
2018
Н, GAR-IRLL,
102
140±2
73,0±2,8
15,4±0,6
56±10
1,0±0,1
21,7±0,3
SUB1
IRLYN-SUB
HHZ8 SAL 14 SUB1 Филиппины
2018
Н, IRLYN-SUB
113
146±2
71,0±2,6
22,1±1,1
92±8
1,2±0,2
22,7±0,3
HHZ9 DT12 SUB1
Филиппины
2018
Н, GSR-Rell
104
140±2
70,0±3,5
17,4±0,3
99±7
1,3±0,2
14,0±0,1
IR 50
Филиппины
2020
В, IRTON
80
118±3
66,0±2,8
17,8±0,6
82±12
1,3±0,3
18,7±1,2
IR14 L 110
Филиппины
2020
С, IRLON
95
134±1
76,0±3,8
19,6±0,7
115±17
1,6±0,3
22,3±0,3
Kirkpinar
Филиппины
2019
В, IRLON
84
125±2
113,0±4,1
16,2±0,5
94±6
2,7±0,4
35,2±0,5
Xiannui
Китай
2021
В, IRCTN
75
112±4
88,0±4,6
21,4±0,5
155±12
2,7±0,4
26,3±0,5
Zhongyon
Китай
2019
В, IRCTN
70
110±4
68,0±4,5
20,2±1,0
203±21
4,3±0,3
27,9±0,3
Zhongyon 207
Китай
2020
В, IRCTN
92
130±2
71,0±2,2
19,3±0,5
125±24
2,7±0,5
22,1±0,1
Аметист
Россия
2019
Н
86
120±2
94,0±4,3
13,8±0,5
98±7
2,8±0,5
30,6±0,5
Анаит
Россия
2021
Н
67
102±1
90,0±7,3
18,4±0,6
78±10
2,9±0,5
37,2±0,5
Велес
Россия
2021
Н
88
125±3
73,0±3,7
16,2±0,2
189±21
3,8±0,5
26,4±0,2
Виола
Россия
2021
С, фиолетовозерный
82
118±3
97,0±5,0
14,2±0,4
67±5
1,7±0,3
23,5±0,3
Вита
Россия
2021
В, фиолетовозерный
70
106±4
88,0±3,3
18,3±0,4
121±13
2,6±0,6
22,1±0,3
Гамма
Россия
2019
В
77
115±4
79,0±6,1
16,0±1,0
124±12
3,4±0,6
26,8±0,5
Девзра
Узбекистан
2020
В
70
105±2
130,0±5,4
20,5±0,7
82±7
2,6±0,4
30,7±0,5
Жемчуг
Россия
2019
В
67
102±2
90,0±4,8
13,9±0,3
88±12
2,0±0,2
24,0±0,2
Красноармейский
76±10
1,4±0,2
30,2±0,5
313
Россия
2020
В
61
98±3
105,0±2,2
15,4±0,2
Кубань 3
Россия
2021
В
63
100±4
114,0±3,5
16,0±0,4
67±15
1,6±0,2
28,7±0,5
Лидер
Россия
2021
В
78
116±5
101,0±5,1
16,3±0,7
127±12
2,9±0,5
26,2±0,3
Наташа
Россия
2019
Н
77
118±2
74,0±8,2
18,5±0,5
91±7
1,7±0,2
25,6±0,7
-
2. Значения критериев оценки достоверности различий при сравнении длины колеоптилей проростков у сортов риса ( Oryza sativa L.) разного происхождения из УНУ Коллекция генетических ресурсов риса Федерального научного центра риса (г. Краснодар) в условиях нормоксии, гипоксии и результаты тетразоли-евого теста ( n = 5, N = 3, M ±SEM, лабораторный тест)
Сорт, сортообразец
Значения критерия
Значение тетразолиевого теста, отн. ед.
Манна-Уитни-Уилкоксона
FDR
нормоксия
гипоксия
AA WAB 56-125
4,32E - 20
1,30E - 19
0,077±0,007
0,008±0,001
HHZ11 Y6-Y2-SUB1
2,61E - 08
3,91E - 08
0,013±0,002
0,004±0,001
HHZ8 SAL 14 SUB1
8,05E - 11
1,26E - 10
0,027±0,005
0,005±0,001
HHZ9 DT12 SUB1
1,66E - 13
3,41E - 13
0,012±0,001
0,004±0,001
IR 50
5,52E - 47
1,99E - 45
0,050±0,012
0,006±0,001
IR14 L 110
1,15E - 20
3,77E - 20
0,131±0,017
0,008±0,0001
Kirkpinar
5,90E - 31
1,06E - 29
0,502±0,014
0,007±0,001
Xiannui
1,25E - 17
2,99E - 17
0,046±0,007
0,004±0,001
Zhongyon
3,46E - 11
5,66E - 11
0,269±0,013
0,005±0,001
Zhongyon 207
1,77E - 14
3,99E - 14
0,087±0,010
0,005±0,001
Аметист
4,94E - 29
5,93E - 28
0,057±0,009
0,006±0,001
Анаит
6,64E - 22
2,39E - 21
0,083±0,011
0,006±0,001
Велес
2,69E - 05
3,34E - 05
0,090±0,005
0,008±0,001
Виола
0,193686*
0,193686*
0,094±0,019
0,010±0,001
Вита
1,56E - 18
4,02E - 18
0,160±0,019
0,010±0,001
Гамма
5,98E - 24
3,07E - 23
0,221±0,018
0,010±0,001
Девзра
4,14E - 07
5,74E - 07
0,550±0,009
0,006±0,001
Жемчуг
5,05E - 19
1,40E - 18
0,275±0,025
0,005±0,001
Красноармейский 313
0,134287*
0,138124*
0,148±0,007
0,015±0,002
Кубань 3
0,000117
0,00014
0,172±0,009
0,017±0,001
Лидер
4,98E - 06
6,64E - 06
0,232±0,010
0,010±0,001
Наташа
4,47E - 27
3,22E - 26
0,050±0,006
0,008±0,001
Новатор
8,66E - 08
1,25E - 07
0,069±0,009
0,010±0,001
Олимп
2,94E - 28
2,64E - 27
0,201±0,043
0,006±0,001
ПВ-1 IRBLSH
6,87E - 13
1,18E - 12
0,028±0,003
0,010±0,001
Привольный 4
0,000828
0,000932
0,334±0,015
0,011±0,001
Рапан
3,53E - 13
6,68E - 13
0,020±0,004
0,004±0,001
Регул
0,001238
0,00135
0,160±0,006
0,010±0,001
Соната
2,91E - 22
1,31E - 21
0,149±0,021
0,008±0,001
Спринт
1,58E - 05
2,03E - 05
0,129±0,024
0,011±0,001
Продолжение таблицы 1
|
Новатор |
Россия |
2021 |
В |
60 |
98±2 |
83,0±3,7 |
15,1±0,2 |
86±10 |
2,3±0,3 |
24,6±0,2 |
|
Олимп |
Россия |
2019 |
В |
85 |
122±4 |
89,0±3,4 |
15,3±0,4 |
130±15 |
3,0±0,7 |
23,5±0,5 |
|
ПВ-1 IRBLSH |
Филиппины |
2020 |
В, краснозерный |
100 |
140±1 |
101,0±4,5 |
19,1±0,4 |
117±8 |
2,6±0,8 |
24,2±0,2 |
|
Привольный 4 |
Россия |
2021 |
Н |
77 |
115±3 |
92,0±6,2 |
15,8±0,6 |
120±22 |
3,6±0,6 |
24,8±0,3 |
|
Рапан |
Россия |
2021 |
С |
78 |
116±2 |
85,0±2,4 |
15,3±0,6 |
121±10 |
3,2±0,3 |
26,3±0,3 |
|
Регул |
Россия |
2021 |
В |
82 |
118±2 |
92,0±2,7 |
16,5±0,5 |
105±15 |
2,5±0,3 |
30,4 ±0,5 |
|
Соната |
Россия |
2019 |
Н |
84 |
120±2 |
87,0±2,5 |
14,3±0,4 |
97±12 |
2,5±0,3 |
26,7±0,2 |
|
Спринт |
Россия |
2021 |
В |
74 |
106±4 |
87,0±7,1 |
18,1±1,1 |
135±24 |
3,8±0,5 |
27,7±0,9 |
|
Титан |
Россия |
2021 |
В |
74 |
115±5 |
86,0±4,3 |
14,6±0,8 |
104±6 |
3,1±0,3 |
35,0±0,4 |
|
Уссур |
Россия, Приморье |
2020 |
В |
63 |
98±1 |
86,0±6,0 |
19,7±0,6 |
122±10 |
3,2±0,3 |
29,2±0,4 |
|
Фонтан |
Россия |
2020 |
В |
64 |
102±3 |
100,0±5,1 |
22,1±1,1 |
102±7 |
2,2±0,4 |
26,4±0,5 |
|
Хазар |
Россия |
2021 |
Н |
74 |
110±4 |
89,0±3,2 |
17,1±0,6 |
103±13 |
2,7±0,4 |
26,6±0,3 |
|
Шарм |
Россия |
2019 |
В |
65 |
100±2 |
84,0±4,1 |
19,8±0,8 |
102±22 |
2,3±0,2 |
26,9±0,5 |
|
Южная ночь |
Россия |
2021 |
Н, фиолетовозерный |
80 |
120±2 |
68,0±3,3 |
16,5±0,5 |
96±9 |
2,3±0,5 |
21,3±0,3 |
П р и м е ч ани е. Н, С, В — сорта и сортоформы соответственно с низкой, средней и высокой скоростью роста на ранней стадии онтогенеза. Все сорта белозерные, за исключением тех, для которых указан иной цвет зерна. IRLON — укороченного затопления, IRCTN — питомник холодостойких, IRTON — искусственно орошаемый, рис умеренных широт, GAR-IRLL — орошаемых низменностей, питомник зеленых технологий; GSR-Rell — поливной, супер-рис, питомник зеленых технологий; IRLYN-SUB — глубоководный.
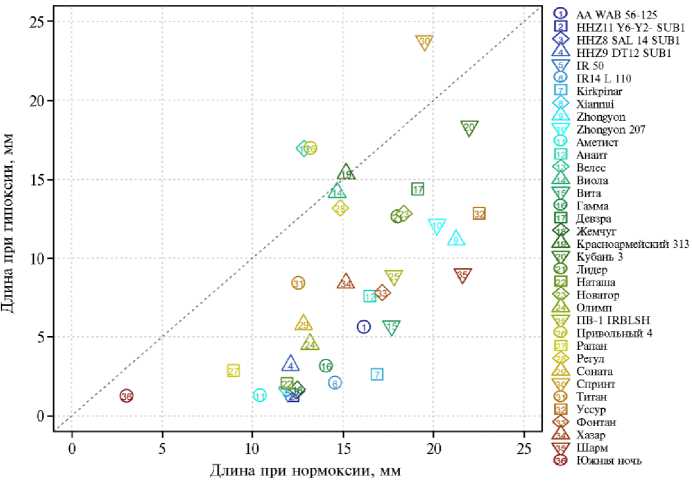
Рис. 1. Выборочные медианы длин колеоптилей проростков у сортов риса ( Oryza sativa L.) разного происхождения из УНУ Коллекция генетических ресурсов риса Федерального научного центра риса (г. Краснодар) после 4 сут проращивания при нормоксии и гипоксии (лабораторный тест) . Линия по диагонали соответствует равенству длины при аноксии и гипоксии.
Продолжение таблицы 2
|
Титан |
0,004084 |
0,004324 |
0,092±0,013 |
0,014±0,001 |
|
Уссур |
1,71E - 13 |
3,41E - 13 |
0,453±0,014 |
0,006±0,001 |
|
Фонтан |
2,34E - 24 |
1,40E - 23 |
0,107±0,022 |
0,010±0,001 |
|
Хазар |
5,18E - 13 |
9,33E - 13 |
0,268±0,011 |
0,006±0,001 |
|
Шарм |
5,55E - 22 |
2,22E - 21 |
0,234±0,005 |
0,011±0,001 |
|
Южная ночь |
0,000305 |
0,000354 |
0,061±0,013 |
0,009±0,002 |
Примечание. FDR — уровень ложноположительных результатов (false discovery rate). Для всех сортов варианты нормоксия и гипоксия в тетразоливом тесте имели статистически значимые различия ( t -тест, p ≤ 0,05).
* Различия в условиях нормоксии и гипоксии недостоверны.
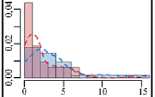
Частью адаптивной стратегии на уровне популяции может быть изменение распределений значений признаков в ответ на изменение условий среды. При построении гистограмм оценки плотности вероятности значений длин колеоптилей проростков риса в различных условиях аэрации можно видеть, что изменения роста соответствовали нескольким паттернам.
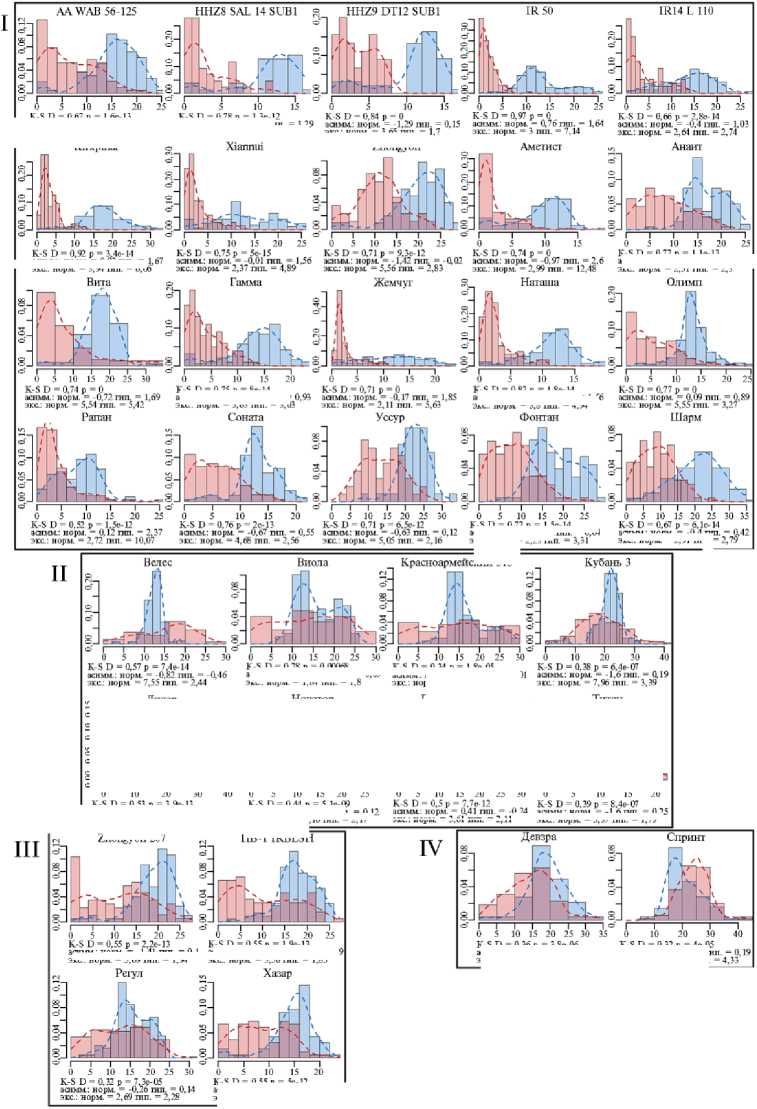
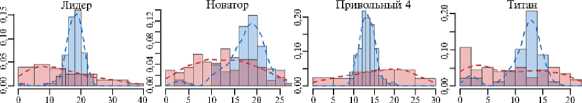
Самым распространенным ответом (рис. 2, I паттерн) оказался сдвиг распределения к малым значениям длин (рост коэффициента асимметрии). При этом для гистограмм при гипоксии было свойственно наличие высокого пика у малых, близких к нулю значений, в сочетании с протяженным «хвостом» значений справа. Это свидетельствует о том, что наиболее часто встречающейся реакцией на гипоксию колеоптилей риса в этой группе было подавление роста с сохранением небольшой части растений, продолжающих рост. Такая стратегия, по-видимому, связана с ожиданием более благоприятных условий и близка стратегии покоя LOQS. Более того, именно в эту группу входили все филиппинские сортоформы-носители аллеля SUB1A — наиболее яркие представители такой стратегии адаптации.
II паттерн (см. рис. 2) заключался в снижении остроконечности (эксцесса) распределения вероятности значений длин при гипоксии. Существенную долю при этом составляли настолько большие растения, которых не было или почти не было при нормоксии. То есть в ответ на стресс популяция распределяла риски равномерно по всем возможным вариантам, что может позволить выжить хоть некоторым особям в неопределенной ситуации.
III паттерн представлял комбинацию из двух предыдущих. Распределения при гипоксии становились сглаженными и сдвигались влево. При этом большая часть была сосредоточена в области небольших значений, а также в области значений, характерных для нормоксии. Примерно половина растений замедляла рост, а другая часть продолжала расти как при нормоксии. Следовательно, на уровне популяции может быть зафиксирована доля потерь, зависящих от выбора стратегии.
Растения, использующие IV паттерн, показали наименьшие изменения распределения. При этом у сорта Девзра отмечали небольшое снижение остроконечности распределения и сдвиг влево, что сходно с III паттерном. В свою очередь у сорта Спринт была выявлена уникальная реакция — явное смещение распределения длин вправо. То есть только у него наблюдались черты стратеги избегания (LOES), связанные с некоторым усилением роста (см. рис. 1, 2). Также уникальным паттерном роста (V паттерн) обладал сорт Южная ночь, колеоптили проростков которого медленно росли как в контроле, так и при затоплении (см. рис. 2).
Можно заключить, что выявленный нами ростовой ответ колеоптилей соответствовал началу адаптационных процессов уже на первых этапах онтогенеза при развитии проростков риса, а паттерны изменения ростового ответа отражали интенсивность проявления той или иной стратегии адаптации.
A.-» U — V,Jj Р — .>,№15 эсимм: норм. = -1,87 пш. = 0.61 экс.: норм. = 10,81 гип. = 2,56
к-М) = 0,7Оп=2е-ГЗ экс.: норм.' = 3,61 пт. = 2.11
К-1 D = 0,55 р = JJe-13 асимм.: норм. = -1,41 пт. = 0,1 асимм.: норм. = -0,53 пш. = 0,39
экс.: норм. = 3,65 тип. = 1.7 Zhongyon
Южная ночь
K-S D = Ц55 р = 5е-12 асимм.: норм = -1,3 пш. = 0,21 эка: норм. = 5,6 гип. = 2,18
К S D = 0,36 р = Зле-06 К S D = 0,32 р - 4^05 эсимм.: норм. = -0,26тин. = 0,08 асимм.: норм. = 0,86 ти экс.: норм = 4,07 гип. = 2,77 эка: норм. = 3,65 гаи.
Рис. 2. Гистограммы распределения длин колеоптилей проростков у сортов риса ( Oryza sativa L.) разного происхождения из УНУ Коллекция генетических ресурсов риса Федерального научного центра риса (г. Краснодар) после 4 сут проращивания в условиях нормоксии (синий цвет) и гипоксии (красный цвет): I-V — паттерны ответа. Абсцисса — длина, мм, ордината — плотность вероятности. Пунктирные линии — ядерная оценка плотности с функцией Епанечникова, K-S — тест Колмогрова-Смирнова, асимм. — коэффициент асимметрии, экс. — коэффициент эксцесса, норм. — нормоксия, гип. — гипоксия.
К-8 И = 0,67 р = 1,6е-13 асимм.: норм. = -1,08 пш. = 0,51 эка: норм = 4,52 пш. = 2,24
Kirkpinar
К-» U = 0,78 р = 1,Зе-12 эсимм.: норм. = 1.46 пш. = 1,29 эка: норм. = 4,3 тип, = 3,81
асимм: норм. = 0,02 гни. = 1 экс.: норм. — 3,94 пш. — 6,06
K-S D = 0,75 р = 8е-14 а(имм: норм. = -0,87 тип. = экс.: норм. = 3,63 тип. = 3,03
K-S D = 0,82 р = 1*86-14 асимм.: норм. = -0,7 тип. = 1,56 экс.: норм = 3.8 пш. = 4.94
К-8 U = 0,72 р = 1.5е-14 эсимм.: норм. = 0.16 пт. = 0,61 экс.: норм. = 2,23 гип
КМ1- I.1...-. р 1.1с- 1 ' асимм.: норм. = 0,05 пш. = ( экс.: норм = 2,51 гаи = 2,3
эсимм: норм, = -0.4 пт экс.: норм. = 2,91 пт. =
К-8 D = 0.28 р = U.UOUM затми: норм. = 0.31 гип. = -0,09 экс.: норм. = 1,84 тип. = 1,8
K S и = 0,34 р = 1,8е-05 асимм: норм. = 0.81 пш. = -0,01 — - 2,78 тип. = 1,86
К О и = 0,44 р = 5,16-09 аатмм: норм. = -1,1 тип. = 0,12 экс.: норм. = 5,16 тип. = 2,17
эсимм: норм._= -1.6 тип. = и, экс.: норм. = 5,37 тип. = 1,73
Zhongyon 207
ПВ-1 [RBLSH
экс.: норм. = 5,69 inn. = 1,91
экс.: норм = 3,56 тип. = 1,85
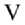
K-S D = 0,32 р = 0,00059 асимм: норм. = 1,6 тип. = 2.05 экс.: норм. =4,96 пт. = 8,38
Хорошо известно, что колеоптили увеличиваются в длину за счет клеточного роста растяжением (28). Такой рост, как и любой другой, тре- бует затрат энергии, однако именно энергетические ресурсы клетки лимитированы при недостатке кислорода. Разнообразие паттернов ростовой реакции, по-видимому, обусловлено изменениями, происходящими в метаболизме растительного организма при гипоксии. Кислородное голодание приводит к дефициту энергии (3, 6). Тормозится окислительное фосфорилирование, снижается продукция активных форм кислорода (АТФ). Единственным источником АТФ становится гликолиз, переходящий в брожения. Запускаются ацидоз цитоплазмы и накопление токсичных продуктов обмена, в частности, ацетальдегида и этанола (1, 3, 6). Помимо этого, гипоксия тормозит биосинтез белка и других полимеров (6), индуцирует генерацию активных форм кислорода и азота (6, 29, 30) которые вызывают окислительные повреждения липидов и белков (31, 32). Как обсуждалось ранее, скудные запасы энергии, вырабатываемой при метаболизации крахмала, проростки риса тратят в зависимости от выбранной стратегии устойчивости: растения, придерживающиеся стратегии избегания (LOES), — на стимуляцию роста, а те, которые тормозят рост (LOQS), — на поддержание клеточных структур (2-6, 11, 12).
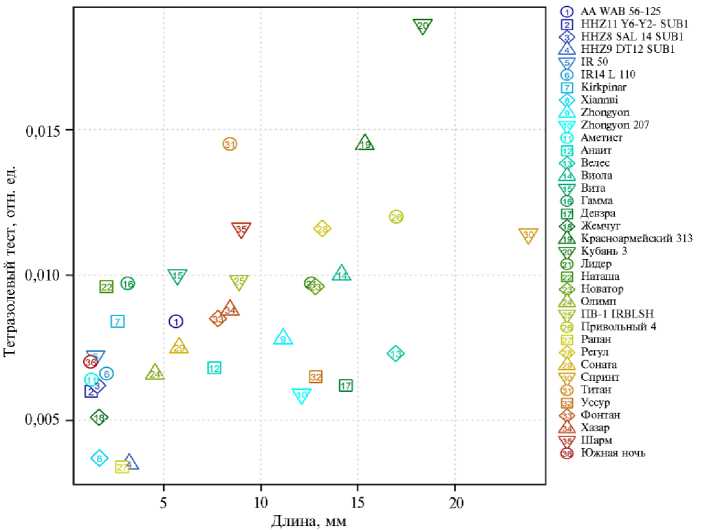
Рис. 3. Связь выборочных медиан длин колеоптилей проростков у сортов риса ( Oryza sativa L.) разного происхождения из УНУ Коллекция генетических ресурсов риса Федерального научного центра риса (г. Краснодар) после 4 сут проращивания при гипоксии с результатами тетразолиевого теста.
В связи с этим следующий этап нашей работы был сфокусирован на оценке жизнеспособности колеоптилей с помощью тетразолиевого теста, который оценивает интенсивность метаболизма. Известно, что чем насыщеннее окрашивание, которое развивается в тесте с солями тетразолия, тем выше жизнеспособность. У всех протестированных форм жизнеспособность по тетразолиевому тесту значительно снижалась в условиях гипоксии по сравнению с контролем (t-тест, p ≤ 0,05, см. табл. 2). Однако у медленнорастущих форм это снижение было не таким сильным, как у быстрорастущих, что может быть связано и с интенсивностью обмена веществ, и с выбранной стратегией адаптации. Тетразолиевый тест характеризует метабо- лическую активность тканей. У быстрорастущих форм метаболизм интенсивный, и в условиях затопления его торможение было существенно (в 1020 раз). У медленнорастущих форм он и так замедлен, и его снижение при гипоксии было не таким интенсивным, но во всех случаях статистически значимым (в 7 раз). Более того, между интенсивностью окраски тетразо-лием и длиной колеоптиля показана положительная зависимость (р = 0,69; p = 10-5), особенно при гипоксии (рис. 3). У наиболее быстрорастущих сортов (Спринт, Кубань 3, Красноармейский 313) интенсивность окраски солей тетразолия была выше как в контроле (в 6 раз), так и в опыте (в 2 раза), по сравнению с медленнорастущими формами (Аметист, Рапан, Южная ночь, Xiannui, филиппинские SUB1A-сорта).
Изменение интенсивности окрашивания тетразолием ранее было использовано для тестирования жизнеспособности проростков пшеницы и риса, различающихся по устойчивости к недостатку кислорода (33, 34). Показано, что тест применим для оценки устойчивости надземной части. У риса побеги меньше повреждаются в условиях кислородного голодания.
Таким образом, протестированные сорта и формы риса из коллекции Федерального научного центра риса значительно различались по интенсивности роста. На стадии прорастания в условиях полевого эксперимента было выявлено три группы сортов с высокой, средней и низкой скоростью роста. Именно в этот период развивающиеся проростки были затоплены. Однако интенсивность роста на ранней стадии онтогенеза не коррелировала с высотой взрослого растения, что можно объяснить отсутствием стандартизации этапа прорастания (7 сут), а также наложением дополнительного стрессового фактора — низкой температуры (14 °С). Последующий лабораторный этап анализа позволил протестировать рост колеоптилей на 4-е сут развития. Самыми быстрорастущими оказались сорта Спринт и Кубань 3, длина колеоптилей которых достигала 19-25 мм как в контроле, так и при затоплении. Отметим, что сорт отечественной селекции Кубань 3 достигал максимальной высоты в полевых исследованиях. Он был отнесен к паттерну II при анализе распределения длин колеоптилей у проростков риса разных сортов, в то время как сорт Спринт — к паттерну IV. Самыми медленнорастущими отечественными сортами были Аметист, Жемчуг, Наташа, Рапан и Южная ночь. В контроле их колеоптиль достигал 10 мм, а при гипоксии — около 3 мм. Все эти сорта отнесли к паттерну I, как и сорта филлипинской селекции, несущие аллель SUB1A, для которых характерна стратегия торможения роста (LOQS). Тем самым полученные нами результаты впервые указывают на вариативность адаптационных механизмов, отражающихся в интенсивности ростовых процессов уже на ранних этапах онтогенеза. На торможение метаболизма при недостатке кислорода указывает сравнительный анализ окраски тканей колеоптилей солями тетразолия. У быстрорастущих сортов, в том числе у сортов Кубань 3 и Спринт, она была в 6 раз более интенсивной, чем у медленно растущих — Аметист, Рапан, Южная ночь, Xiannui, филиппинские SUB1A-сорта. Полученные нами данные позволяют заключить, что интенсивность роста колеоптилей коррелирует с интенсивностью метаболизма и устойчивостью к затоплению. К сожалению, механизмы роста колеоптилей при недостатке кислорода еще окончательно не установлены. Требуется продолжение исследований для понимания того, к какой из стратегий устойчивости относятся протестированные сорта и каковы факторы, определяющие их возможное соотношение. Однако представленные результаты уже сейчас ха- рактеризуют устойчивость к недостатку кислорода генотипов, хранящихся в коллекции Федерального научного центра риса.
Список литературы Показатели роста и устойчивости к дефициту кислорода у колеоптилей Oryza sativa L. из коллекции Федерального научного центра риса
- Voesenek L.A.C.J., Bailey-Serres J. Flooding tolerance: O2 sensing and survival strategies. Current Opinion in Plant Biology, 2013, 16(5): 647-653 (doi: 10.1016/j.pbi.2013.06.008).
- Mommer L., Visser E.J.W. Underwater photosynthesis in flooded terrestrial plants: a matter of leaf plasticity. Annals of Botany, 2005, 96(4): 581-589 (doi: 10.1093/aob/mci212).
- Bailey-Serres J., Voesenek L.A.C.J. Flooding stress: acclimations and genetic diversity. Annual Review of Plant Biology, 2008, 59: 313-339 (doi: 10.1146/annurev.arplant.59.032607.092752).
- Polko J.K., Voesenek L.A.C.J., Peeters A.J.M., Pierik R. Petiole hyponasty: an ethylene-driven, adaptive response to changes in the environment. AoB PLANTS, 2011, 2011: plr031 (doi: 10.1093/aobpla/plr031).
- Voesenek L.A.C.J., Bailey-Serres J. Flood adaptive traits and processes: an overview. New Phy-tologist, 2015, 206(1): 57-73 (doi: 10.1111/nph.13209).
- Chirkova T., Yemelyanov V. The study of plant adaptation to oxygen deficiency in Saint Peters-burg University. Biological Communications, 2018, 63(1): 17-31 (doi: 10.21638/spbu03.2018.104).
- Yemelyanov V.V., Shishova M.F. The role of phytohormones in the control of plant adaptation to oxygen depletion. In: Phytohormones and abiotic stress tolerance in plants /N. Khan, R. Nazar, N. Iqbal, N. Anjum (eds.). Springer, Berlin, Heidelberg, 2012: 229-248 (doi: 10.1007/978-3-642-25829-9_10)
- Hartman S., Sasidharan R., Voesenek L.A.C.J. The role of ethylene in metabolic acclimations to low oxygen. New Phytologist, 2021, 229(1): 64-70 (doi: 10.1111/nph.16378).
- Hattori Y., Nagai K., Furukawa S., Song X.-J., Kawano R., Sakakibara H., Wu J., Matsumoto T., Yoshimura A., Kitano H., Matsuoka M., Mori H., Ashikari M. The ethylene response factors SNORKEL1 and SNORKEL2 allow rice to adapt to deep water. Nature, 2009, 460: 1026-1030 (doi: 10.1038/nature08258).
- Winkel A., Pedersen O., Ella E., Ismail A.M., Colmer T.D. Gas film retention and underwater photosynthesis during field submergence of four contrasting rice genotypes. Journal of Experi-mental Botany, 2014, 65(12): 3225-3233 (doi: 10.1093/jxb/eru166).
- Bailey-Serres J., Fukao T., Gibbs D.J., Holdsworth M.J., Lee S.C., Licausi F., Perata P., Voesenek L.A.C.J., van Dongen J.T. Making sense of low oxygen sensing. Trends in Plant Science, 2012, 17(3): 129-138 (doi: 10.1016/j.tplants.2011.12.004).
- Bailey-Serres J., Lee S.C., Brinton E. Waterproofing crops: effective flooding survival strategies. Plant Physiology, 2012, 160(4): 1698-1709 (doi: 10.1104/pp.112.208173).
- Schmitz A.J., Folsom J.J., Jikamaru Y., Ronald P., Walia H. SUB1A-mediated submergence tolerance response in rice involves differential regulation of the brassinosteroid pathway. New Phytologist, 2013, 198(4): 1060-1070 (doi: 10.1111/nph.12202).
- Fukao T., Xu K., Ronald P.C., Bailey-Serres J. A variable cluster of ethylene response factor-like genes regulates metabolic and developmental acclimation responses to submergence in rice. The Plant Cell, 2006, 18(8): 2021-2034 (doi: 10.1105/tpc.106.043000).
- Takahashi H., Sato K., Takeda K. Mapping genes for deep-seeding tolerance in barley. Euphytica, 2001, 122: 37-43 (doi: 10.1023/A:1012608721291).
- Rebetzke G.J., Zheng B., Chapman S.C. Do wheat breeders have suitable genetic variation to overcome short coleoptiles and poor establishment in the warmer soils of future climates? Func-tional Plant Biology, 2016, 43(10): 961-972 (doi: 10.1071/FP15362).
- Brown P.R., Singleton G.R., Tann C.R., Mock I. Increasing sowing depth to reduce mouse damage to winter crops. Crop Protection, 2003, 22(4): 653-660 (doi: 10.1016/S0261-2194(03)00006-1).
- O’Sullivan P.A., Weiss G.M., Friesen D. Tolerance of spring wheat (Triticum aestivum L.) to trifluralin deep-incorporated in the autumn or spring. Weed Research, 1985, 25(4): 275-280 (doi: 10.1111/j.1365-3180.1985.tb00645.x).
- Schillinger W.F., Donaldson E., Allan R.E., Jones S.S. Winter wheat seedling emergence from deep sowing depths. Agronomy Journal, 1998, 90(5): 582-586 (doi: 10.2134/agronj1998.00021962009000050002x).
- Bovill W.D., Hyles J., Zwart A.B., Ford B.A., Perera G., Phongkham T., Brooks B.J., Rebetzke G.J., Hayden M.J., Hunt J.R., Spielmeyer W. Increase in coleoptile length and establishment by Lcol-A1, a genetic locus with major effect in wheat. BMC Plant Biology, 2019, 19: 332 (doi: 10.1186/s12870-019-1919-3).
- Luo H., Hill C.B., Zhou G., Zhang X.-Q., Li C. Genome-wide association mapping reveals novel genes associated with coleoptile length in a worldwide collection of barley. BMC Plant Biology, 2020, 20: 346 (doi: 10.1186/s12870-020-02547-5).
- Ma М., Cen W., Li R., Wang S., Luo J. The molecular regulatory pathways and metabolic adaptation in the seed germination and early seedling growth of rice in response to low O2 stress. Plants, 2020, 9(10): 1363 (doi: 10.3390/plants9101363).
- Коротенко Т.Л., Садовская Л.Л. Результаты применения современных методов при изучении генетического разнообразия Oryza s. L. Труды Кубанского государственного аграрного университета, 2018, 72: 202-206 (doi: 10.21515/1999-1703-72-202-206).
- Коротенко Т.Л., Чижикова С.С., Пустовалов Р.А. Взаимосвязь биологических, биохимических и технологических признаков качества генплазмы коллекции риса. Бюллетень Государственного Никитского ботанического сада, 2019, 133: 174-181 (doi: 10.36305/0513-1634-2019-133-174-181).
- Yemelyanov V.V., Lastochkin V.V., Chirkova T.V., Lindberg S.M., Shishova M.F. Indoleacetic acid levels in wheat and rice seedlings under oxygen deficiency and subsequent reoxygenation. Biomolecules, 2020, 10(2): 276 (doi: 10.3390/biom10020276).
- R Core Team. R: The R Project for Statistical Computing. Доступ по ссылке: https://www.r-project.org/. Дата обращения: 26.02.2023.
- Huang S., Shingaki-Wells R.N., Petereit, J., Alexova R., Millar A.H. Temperature-dependent metabolic adaptation of Triticum aestivum seedlings to anoxia. Scientific Reports, 2018, 8: 6151 (doi: 10.1038/s41598-018-24419-7).
- Pucciariello C. Molecular mechanisms supporting rice germination and coleoptile elongation un-der low oxygen. Plants, 2020, 9(8): 1037 (doi: 10.3390/plants9081037).
- Blokhina O., Fagerstedt K.V. Oxidative metabolism, ROS and NO under oxygen deprivation. Plant Physiology and Biochemistry, 2010, 48(5): 359-373 (doi: 10.1016/j.plaphy.2010.01.007).
- Yemelyanov V.V., Lastochkin V.V., Prikaziuk E.G., Chirkova T.V. Activities of catalase and peroxidase in wheat and rice plants under conditions of anoxia and post-anoxic aeration. Russian Journal of Plant Physiology, 2022, 69(6): 117 (doi: 10.1134/S1021443722060036).
- Chirkova T.V., Novitskaya L.O., Blokhina O.B. Lipid peroxidation and antioxidant systems under anoxia in plants differing in their tolerance to oxygen deficiency. Russian Journal of Plant Physi-ology, 1998, 45(1): 55-62.
- Shikov A.E., Lastochkin V.V., Chirkova T.V., Mukhina Z.M., Yemelyanov V.V. Post-anoxic oxidative injury is more severe than oxidative stress induced by chemical agents in wheat and rice plants. Acta Physiologiae Plantarum, 2022, 44(9): 90 (doi: 10.1007/s11738-022-03429-z).
- Емельянов В.В., Ласточкин В.В., Чиркова Т.В., Слюсаренко А. Роль салициловой кислоты в регуляции механизмов адаптации растений к недостатку кислорода и последующему окислительному стрессу. Мат. докл. VII Съезда физиологов растений России «Физиология растений — фундаментальная основа экологии и инновационных биотехнологий». Нижний Новгород, 4-10 июля 2011. Часть I /Под ред. Вл.В. Кузнецова, А.П. Веселова, Г.А. Рома-нова. Нижний Новгород, 2011: 236-237.
- Yemelyanov V.V., Lastochkin V.V., Chirkova T.V. The role of polyamines in signaling and adap-tation to oxygen deprivation and subsequent re-aeration in plants. Proceedings of 4th International symposium «Plant Signaling & Behavior». St. Petersburg, 19-24 June 2016 /V. Demidchik, O. Voitsekhovskaja, E. Tyutereva, G. Pozhvanov (eds.). SINEL Co. Ltd., SPb, 2016: 107.
