Поливариантность морфогенеза побегов в клонах Salix triandra (Salicaceae) на фоне периодичности ливневых осадков
 на фоне периодичности ливневых осадков")
Автор: Афонин Алексей Алексеевич
Журнал: Бюллетень науки и практики @bulletennauki
Рубрика: Биологические науки
Статья в выпуске: 1 т.7, 2021 года.
Бесплатный доступ
В статье обоснована необходимость изучения развития однолетних черенковых саженцев ивы трехтычинковой ( Salix triandra ). Цель : выявление и анализ сезонной динамики суточного прироста побегов в клонах S. triandra на фоне избыточного атмосферного увлажнения в виде трех ливней в 2020 г. Материал и методы . Методы исследования: селекционные, структурно-морфологические, хронобиологические. Объект: модельная инбредно-клоновая популяция S. triandra первого года жизни. Число клонов 7, повторность шестикратная. Материал: растущие однолетние побеги на однолетних черенковых саженцах; каждый саженец формировался в один побег. Результаты . В условиях эксперимента укореняемость черенков составила 100%. На материнских черенках выкопанных саженцев сформировались диморфные корневые системы. Годичный прирост наиболее сильных побегов составил 175…215 см. Суточный прирост побегов достиг пиковых значений к 10 июня: средний прирост составил 3,6 см/сут, максимальный - 4,4 … 4,9 см/сут. После 10 июня суточный прирост неравномерно снижался до завершения вегетации. Часть побегов завершила рост до начала августа, часть продолжила рост до середины августа. Установлена цикличность сезонной динамики суточного прироста в виде трех пиков, разделенных двумя провалами. Третий пик формируется после третьего ливня. Обсуждение и выводы . Цикличность сезонной динамики суточного прироста побегов характеризуется высокой степенью синхронизации ростовых процессов. Вероятно, на все побеги действует некоторый мощный импульсный синхронизатор неизвестной природы. Не исключено, что периодические ливни могут оказывать модифицирующее действие на механизмы, регулирующие ростовые процессы. Временная дискретность морфогенеза побегов обусловлена полной или частичной реализацией сезонного цикла развития побегов. Продолжительность роста обусловлена межклоновыми и внутриклоновыми различиями. Вероятно, продолжительность морфогенеза контролируется как генетическими, так и эпигенетическими факторами.
Ива трехтычинковая, черенковые саженцы, ризогенез, однолетние побеги, суточный прирост, сезонная динамика, поливариантность онтогенеза, дискретность онтогенеза
Короткий адрес: https://sciup.org/14117925
IDR: 14117925 | УДК: 582.623.2:581.14:57.034 | DOI: 10.33619/2414-2948/62/02
Polyvariance of shoot morphogenesis in clones of Salix triandra (Salicaceae) against the background of the periodicity of rainstorms
The article substantiates the need to study the development of one-year saplings from cuttings of almond willow ( Salix triandra ). Purpose of research : identification and analysis of seasonal dynamics of daily growth of shoots in S. triandra clones against the background of excessive atmospheric moisture in the form of three showers in 2020. Material and methods . Research methods: selection, structural-morphological, chronobiological. Object of research: model inbred-clone one-year population of S. triandra . The number of clones is 7; the recurrence is 6-fold. Material: growing annual shoots on one-year saplings; each sapling was formed into a single shoot. Results . Under experimental conditions, the rooting capacity of cuttings was 100%. Dimorphic root systems were formed on the mother cuttings of the excavated seedlings. The annual growth of the strongest shoots was 175 ... 215 cm. The daily growth in shoots reached peak values by June 10: the average daily growth was 3.6 cm/day, the maximum - 4.4 ... 4.9 cm/day. After June 10, the daily growth rate decreased unevenly until the end of the growing season. Some shoots completed growth before the beginning of August, and some continued growth until the middle of August. The cyclical nature of the seasonal dynamics of daily growth in the form of three peaks separated by two dips is established. The third peak is formed after the third rainstorm. Discussion and conclusions . Cyclical seasonal dynamics of daily growth of shoots is characterized by a high degree of synchronization of growth processes. Probably, all the shoots are affected by some powerful pulse synchronizer of unknown nature. It is possible that periodic showers may have a modifying effect on the mechanisms that regulate growth processes. The temporal discreteness of shoot morphogenesis is due to the full or partial realization of the seasonal cycle of shoot development. The duration of growth is due to interclonal and intraclonal differences. Probably, the duration of morphogenesis is controlled by both genetic and epigenetic factors.
Текст научной статьи Поливариантность морфогенеза побегов в клонах Salix triandra (Salicaceae) на фоне периодичности ливневых осадков
Бюллетень науки и практики / Bulletin of Science and Practice
Keyword: almond willow, Salix triandra , saplings from cuttings, rhizogenesis, annual shoots, daily growth, seasonal dynamics, polyvariance of ontogenesis, discreteness of ontogenesis.
Ива трехтычинковая ( Salix triandra L., 1753) — широко известный представитель рода Salix L., 1753 (Salicaceae Mirb., 1815, Malpighiales Juss. ex Bercht. & J. Presl, 1820) [1–2]. Традиционно относится к секции Amygdalinae Koch, 1837 ( Triandrae Dumort., 1825) подрода Salix Nas., 1936 ( Amerina Dumort., 1862) (s. l.) [1]. Однако по результатам молекулярногенетических исследований таксон Triandrae заслуживает статуса самостоятельного подрода [3–4]. С цитогенетической точки зрения, S. triandra — диплоид (2 n = 2 x = 38) [5]. Не исключено, что в природных популяциях этого вида существует анеуплоидия [6]. Вследствие небольшого генома S. triandra используется как модельный объект для получения новой ценной информации [7].
С практической точки зрения, S. triandra представляет собой ценный растительный ресурс. Для создания энергетических плантаций на пахотных почвах малопригодна, поскольку ее продуктивность в этих условиях существенно ниже, чем у высокопродуктивных видов и гибридов секции Vimen [8]. Однако вследствие своей высокой неприхотливости рекомендована для выращивания на малоплодородных почвах [9]. Используется в защитном лесоразведении для укрепления берегов водоемов [9], а также для фиторемедиационной очистки почв, загрязненных углеводородами [10]. Благодаря продолжительному обильному цветению, является ценным медоносом [11]. Перспективный источник лекарственного сырья [12]. Главное достоинство S. triandra в том, что она дает высококачественный зеленый и белый прут для различных видов плетения [9, 13].
Культуры S. triandra, как правило, создают путем посадки одревесневших неукорененных черенков на постоянное место [9]. Правилам подготовки посадочных мест, заготовки и посадки черенков, а также дальнейшему уходу за черенковыми саженцами посвящен ряд современных исследований [14–15]. Эффективность ризогенеза — образования придаточных корней на черенках — в значительной мере определяет приживаемость черенков. Уже на второй год после посадки развивается мощная, глубоко идущая корневая система, которая обеспечивает засухоустойчивость растений [16]. Хорошо развитая корневая система позволяет максимально использовать водные ресурсы в виде атмосферных осадков, почвенной влаги и грунтовых вод [17]. Ризогенез на высаженных черенках происходит как на всем протяжении черенков (из корневых зачатков под чечевичками, преимущественно, вблизи узлов), так и на срезах [9]. Благодаря этому в ряде случаев формируются диморфные корневые системы, которые позволяют использовать как влагу весенне-летних дождей, так и грунтовые воды с относительно постоянным уровнем [18]. Наблюдаемые изменения климата приводят к непредсказуемости осадков в вегетационный период [19]. При избытке влаги снижается содержание кислорода в почве, и рост корней угнетается [20].
Одно из важнейших требований к культурам ивы — их высокая и стабильная продуктивность, которая зависит от множества разнообразных факторов [21]. Окружающая среда имеет тенденцию оказывать на количественные признаки высокопродуктивных видов Salix более сильное влияние, чем генотип [22]. В частности, факторы окружающей среды контролируют примерно 60% изменчивости урожая биомассы; примерно 10% приходится на генетические факторы; остальные 30% изменчивости обусловлены влиянием неконтролируемых факторов [23]. Для выявления влияния неконтролируемых факторов на рост и развитие ив необходимо вывить соотношение межклоновой и внутриклоновой изменчивости. Изменчивости морфофизиологических и хозяйственно-ценных признаков в клонах ив посвящен ряд исследований [21, 24–25].
Изменчивость морфогенеза побегов есть проявление поливариантности онтогенеза. Онтогенез характеризуется целостностью и в то же время дискретностью, обусловленной сменами этапов развития [26–27]. В традиционном представлении [28], поливариантность онтогенеза обусловлена разнообразием адаптивных реакций в генетически гетерогенных популяциях. Поливариантность онтогенеза определяется не только генетическими, но эпигенетическими механизмами, которые исследованы недостаточно [29]. Сведения о поливариантности морфогенеза нарастающих побегов в культивируемых клонах S. triandra в доступной нам литературе не обнаружены.
Цель данного исследования: выявить и проанализировать закономерности сезонной динамики суточного прироста побегов в клонах S. triandra на фоне избыточного атмосферного увлажнения в виде периодических ливневых осадков.
Материал и методы
Исследования проводились в салицетуме Брянского государственного университета (N53.273096, E34.353077). Тип лесорастительных условий (ТЛУ): D3. Тип растительности: опушка дубравы лещино-снытевой ( Quercetum coryloso-aegopodiosum ). Тип почв: серые лесные на лессовидном суглинке с меловыми подстилающими породами.
Объект исследований: модельная инбредно-клоновая популяция S. triandra , созданная весной 2020 г. Для создания модельной популяции использовалась разработанная нами схема популяционно-клоновой селекции [30].
В качестве исходного генетического материала использовались семена, собранные в 2000 г. с женской особи S. triandra в естественном ивняке зеленой зоны г. Брянска
(N53.201025, E34.507728; Alno-Salicetum cinereae (Kobenza 1950) Pass. 1956). Из этих семян была выращена семья F 0 . В 2010 г. в результате свободного внутрисемейного опыления одна из женских особей F 0 дала семена, из которых была выращена вторая семенная генерация F 1 . В 2017 г. в условиях беспыльцевого режима одна из женских особей F 1 дала семена, из которых была выращена третья семенная генерация F 2 . Подробно история создания данной семьи описана нами ранее [31].
Для создания модельной популяции в качестве родоначальников экспериментальных клонов E использовались сеянцы рассматриваемой семьи. За год до черенкования (весной 2019 г.) маточные особи были коротко обрезаны для получения достаточного количества порослевых побегов. Из нижних и средних частей этих побегов непосредственно перед посадкой нарезались черенки длиной 25 … 30 см при диаметре 0,8 … 1,0 см. Свежесрезанные черенки без какой-либо обработки были высажены в подготовленные с осени посадочные ямы размером 40×40×40 см. Схема размещения тригональная, с расстоянием между посадочными местами 1 м. Дата посадки 10.04.2020. Посадка вертикальная с заглублением черенков до одной почки на уровне почвы. В дальнейшем все черенковые саженцы формировались в один побег (по принципу «один саженец — один побег»). Количество заложенных клонов — 7, повторность — 6-кратная, общее число черенковых саженцев — 42.
В качестве основного контроля K использовался высокопродуктивный клон S. triandra , выделенный нами из естественного ивняка (N53.256240, E34.383236; Salicetum triandro-viminalis (Tx. 1931) Lohm. 1952) зеленой зоны г. Брянска в 2009 г. Черенки контрольного клона K нарезались и высаживались так же, как черенки экспериментальных клонов E . Повторность — 3-кратная. В качестве дополнительного контроля S использовалось 4 четырехлетних сеянца (ортета) той же семьи, что и родоначальники клонов. Весной 2020 г. эти сеянцы были обрезаны на пень, кусты формировались в два побега.
Вегетационный период 2020 г. охарактеризовался избыточным неравномерным атмосферным увлажнением [32]. В апреле количество осадков составило 13 мм (31% от нормы), в мае 138 мм (242% от нормы), в июне 141 мм (177% от нормы), в июле 76 мм (92% от нормы), в августе 44 мм (64% от нормы). Избыточное увлажнение было обусловлено тремя ливневыми дождями: 29.05 … 02.06 (95 мм), 19.06 … 23.06 (94 мм), 14.07 … 15.07 (56 мм). Временны́ е интервалы между днями с максимальным количеством осадков составили 22 и 24 сут. В этих промежутках между ливнями выпадало всего 22 и 8 мм осадков.
В весенне-летний период четыре саженца экспериментальных клонов выкопали для контроля за формированием корневых систем. После завершения вегетационного периода 14 саженцев трех клонов выкопали для окончательного описания корневых систем.
Наблюдения за развитием побегов проводили с 09.05 по 29.08. Каждые 4 сут замеряли длину побегов: L , см (выполнено 1217 измерений для 29 дней наблюдений). Все даты, в которые проводились наблюдения, пронумеровали в соответствии с днями вегетационного периода (1, 5, 9 … t i ). За дату начала вегетации ( t = 1) выбрали 01.05 (средняя многолетняя дата перехода среднесуточных температур через +10 °С). В итоге получили эмпирические ряды траекторий роста побегов — L(t) , см.
Для анализа сезонной динамики суточного прироста побегов использовали алгоритм, подробно описанный ранее [31]. На основании рядов L(t) для каждой даты вычислили ряды сезонной динамики суточного прироста побегов ΔL(t), см/сут, выровненные методом скользящего интервала продолжительностью 8 сут с шагом 4 сут (вычислено 1117 значений ΔL для 25 расчетных дней). На основании рядов ΔL(t) построили соответствующие графики, отражающие зависимость суточного прироста побегов ΔL, см/сут от порядкового дня t вегетационного периода. Группировку побегов производили на основании сравнительного анализа конфигурации графиков ΔL(t). Для каждой группы рассчитали и визуализировали средние (average) ряды динамики ΔL(t) av.
Результаты
Во всех исследованных клонах S. triandra укореняемость черенков составила 100%. На материнских черенках выкопанных саженцев образовалось два типа корней: тонкие и скелетные. Многочисленные тонкие корни (окончательной толщиной порядка 1 … 2 мм и длиной порядка 5 … 10 см) сформировались в первой половине мая. Эти горизонтально ориентированные корни достаточно равномерно распределены по всей подземной части черенков, выраженной приуроченности к узлам не выявлено. Немногочисленные скелетные корни (окончательной толщиной порядка 5 мм и длиной свыше 15 см) начали формироваться в первой декаде июня. Эти корни, направленные вниз и вбок, образовались на нижних срезах черенков (в отдельных случаях единичные скелетные корни образовались в средней части черенка). На 14 саженцах трех клонов, выкопанных после завершения вегетационного периода, различия в интенсивности развитости корневых систем не обнаружены.
Для исследования морфогенеза в экспериментальных клонах было использовано 38 саженцев. На четырех из них рост побегов завершился к 20.07, на остальных завершение роста побегов происходило в интервале с 24.07 по 25.08. По длине вызревших побегов выделено две группы саженцев: с побегами длиной 133 … 163 см и с побегами длиной 175 … 215 см. На трех саженцах длина побегов составила 125 см, а на одном — 288 см; эти саженцы были исключены из дальнейшего анализа. Между продолжительностью роста побегов и их длиной установлена достаточно высокая положительная корреляция: r = 0,774 ( P < 0,01). На однолетних черенковых саженцах контрольного клона K длина побегов составила 198 … 273 см, а на четырехлетних контрольных сеянцах — 278 … 330 см.
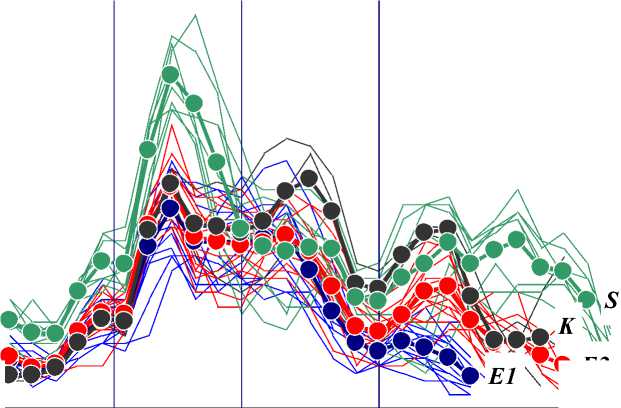
Сравнительный анализ рядов сезонной динамики суточного прироста побегов — Δ L(t) — в экспериментальных клонах показал, что саженцы, продолжившие рост после 20.07, образуют две группы: с побегами, закончившими рост до 01.08 (группа E 1), и с побегами, продолжившими свой рост после 01.08 (группа E 2). Саженцы двух клонов вошли только в первую группу, саженцы еще двух клонов — только во вторую группу, а саженцы трех клонов распределились по двум группам. Для каждой группы — Е 1 и Е 2 — были вычислены средние траектории динамики суточного прироста — Δ L(t) av . Средние траектории были вычислены также для саженцев контрольного клона K и контрольной группы сеянцев S . Визуализированные эмпирические и средние вычисленные ряды сезонной динамики суточного прироста всех побегов показаны на Рисунке 1.
Все ряды Δ L(t) сходны между собой по конфигурации, различаясь, в основном, лишь по размаху изменчивости Δ L . Рекордные суточные приросты — 6,1 … 6,8 см/сут — выявлены на побегах контрольной группы четырехлетних сеянцев. На побегах однолетних черенковых саженцев максимальный суточный прирост составил 2,4 … 4,3 см/сут (на отдельных побегах до 4,4 … 4,9 см/сут).
7,0 1
0,0
6,0 1
5,0 d
4,0 1
3,0
2,0 1
1,0 1
E2
10 20 30 40 50 60 70 80 90 100 110 120
Рисунок 1. Сезонная динамика суточного прироста всех исследованных побегов. По оси абсцисс — порядковые дни вегетационного периода, по оси ординат — суточный прирост побегов, см/сут. Группы побегов обозначены разными цветами: E 1 — синий, E 2 — красный, K — черный, S — зеленый. Средние ряды показаны увеличенными маркерами и подписаны. Вертикальными линиями показаны дни с максимумами ливневых осадков.
Визуализированные эмпирические и средние вычисленные ряды сезонной динамики суточного прироста побегов в экспериментальных клонах показаны на Рисунке 2.
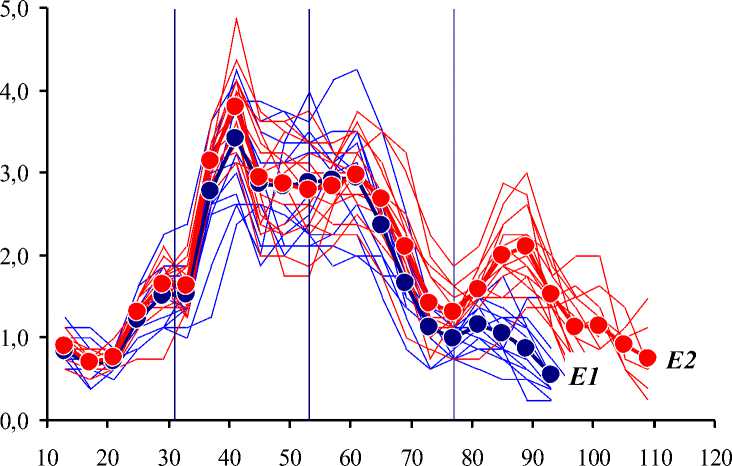
Рисунок 2. Сезонная динамика суточного прироста побегов в экспериментальных клонах. По оси абсцисс — порядковые дни вегетационного периода, по оси ординат — суточный прирост побегов, см/сут. Обозначения те же, что на Рисунке 1.
В начале вегетационного периода суточный прирост — ΔL, см/сут — всех побегов возрастал с некоторой задержкой прироста в конце мая – начале июня (что соответствует первому ливню) [32]. Максимальный прирост — 4,4 … 4,9 см/сут — зафиксирован 10.06. В этот момент средний прирост был несколько выше в группе E2. Далее ΔL неравномерно снижался. В период с 14.06 по 30.06 динамика ΔL дестабилизировалась при отсутствии межгрупповых различий; однозначная связь динамики ΔL с вторым ливнем не установлена. В конце июня – начале июля ΔL несколько увеличился. В первой половине июля ΔL снизился, причем ΔL снижался быстрее в группе E1. После 16.07 (что совпадает с датой третьего ливня) ΔL увеличился, но в группе E1 сразу же стал снижаться вплоть до окончания роста побегов в конце июля. В группе E2 ΔL возрастал до конца июля и только затем стал снижаться до окончания роста побегов в конце августа.
Визуализированные средние ряды сезонной динамики суточного прироста побегов — Δ L(t) av – в двух группах экспериментальных клонов Е 1 и Е 2, контрольного клона K и контрольной группы сеянцев S показаны на Рисунке 3.
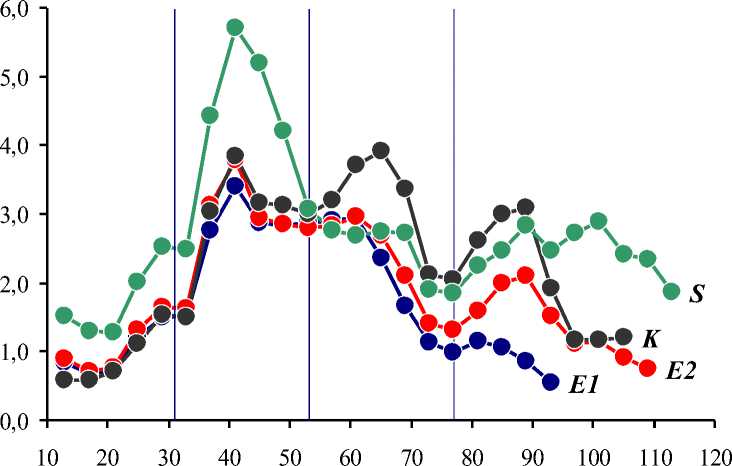
Рисунок 3. Средняя динамика суточного прироста побегов. По оси абсцисс — порядковые дни вегетационного периода, по оси ординат — суточный прирост побегов, см/сут. Обозначения те же, что на Рисунке 1 и Рисунке 2.
В целом сезонная динамика суточного прироста побегов характеризуется ярко выраженной цикличностью, которая проявляется в виде правильного чередования пиков и провалов Δ L . Все средние ряды Δ L(t) av синхронизированы между собой, различаясь лишь по размаху изменчивости Δ L . Наиболее отчетливо выражены главный пик (10.06) и главный провал (16.07). На побегах контрольной группы сеянцев S выявлено 4 пика Δ L (второй пик слабо выражен), на побегах контрольного клона K — 3 отчетливых пика Δ L , на побегах экспериментальной группы Е 2 также 3 пика Δ L (второй пик слабо выражен), на побегах экспериментальной группы Е 1 — 2 основных пика Δ L (второй пик слабо выражен) и очень слабый третий пик.
Временные интервалы между 1, 2 и 3 пиками ΔL составили 24 сут, что почти полностью совпадает с интервалами между ливнями (22 и 24 сут). Однозначная реакция суточного прироста на первый ливень не выявлена ни в одной из групп побегов. Возможное положительное («омолаживающее») действие третьего ливня выявлено для всех побегов. Такое же действие второго ливня выявлено только для побегов на черенковых саженцах контрольного клона K. На всех остальных побегах отчетливая реакция суточного прироста на второй ливень не выявлена.
Обсуждение и выводы
Родоначальники экспериментальных клонов S. triandra , как и сеянцы контрольной группы S , были получены путем регулярного инбридинга на протяжении трех семенных генераций, что позволило частично гомозиготизировать исходный генетический материал. Не исключено, что семена третьей генерации образовались путем нерегулярного гаметофитного апомиксиса в форме редуцированного партеногенеза. Восстановление диплоидности в этом случае возможно за счет пропуска второго мейотического деления или путем постзиготической дигаплоидизации. Такой механизм апомиксиса у диплоидов также позволяет гомозиготизировать популяцию [33–34]. Таким образом, в данном исследовании межклоновые различия обусловлены использованием выровненного генетического материала.
Исследованные однолетние черенковые саженцы сформировали диморфные корневые системы. Ранние тонкие (эфемерные) корни, распределенные по всей длине черенка, вероятно, образовались из уже имеющихся корневых зачатков. Очевидно, эти корни обеспечивают водоснабжение черенковых саженцев весной и в начале лета при достаточном увлажнении хорошо аэрируемых верхних слоев почвы. Поздние скелетные (толстые, грубые, глубокие) на нижних срезах черенков, вероятно, образовались через каллус de novo . Очевидно, эти корни обеспечивают водоснабжение черенковых саженцев в летние месяцы, поглощая влагу из глубоких слоев почвы. Они же служат основой для формирования многолетней корневой системы. Рост глубоких корней может ограничиваться возникновением субанаэробных условий — нехваткой кислорода при избыточном влагосодержании. Можно предположить, что описанный механизм формирования диморфных корневых систем обеспечивает устойчивость черенковых саженцев S. triandra к неравномерному увлажнению в течение первого года жизни.
Выявленная цикличность сезонной динамики суточного прироста побегов Δ L(t) , характеризуется высокой степенью синхронизации ростовых процессов (особенно синфазным усилением Δ L 10.06 и синфазным ослаблением Δ L 16.07, разделенными интервалом в 36 сут) на всех исследованных побегах. Вероятно, на все побеги действует некоторый мощный импульсный синхронизатор неизвестной природы.
Чередование пиков и провалов Δ L с периодичностью 24 сут позволяет предположить, что на ритмику развития побегов влияют внешние факторы в виде ливневых осадков с периодичностью 22 … 24 сут. Однако эта гипотеза подтверждается лишь частично. Можно допустить положительное («омолаживающее») действие третьего ливня (в середине июля), которое совпадает с увеличением суточного прироста всех исследованных побегов. «Ливневая» гипотеза не позволяет объяснить резкое снижение суточного прироста в промежутках между ливневыми дождями. Нельзя отрицать влияние ливневых осадков на динамику суточного прироста побегов. Однако, скорее всего, ливни оказывают модифицирующее действие на механизмы, регулирующие ростовые процессы.
Полный сезонный (точнее, субсезонный, или субаннуальный) цикл развития побегов в летние месяцы (июнь — первая половина августа) включает четыре многодневных (инфрадианных) цикла колебаний суточного прироста. В полной мере сезонный цикл реализован на побегах контрольной группы сеянцев S . На поздно завершивших рост побегах черенковых саженцев (контрольный клон K и группа E 2 экспериментальных клонов)
реализовано три инфрадианных цикла. На рано завершивших рост побегах черенковых саженцев (группа E 1 экспериментальных клонов) полностью реализовано лишь два инфрадианных цикла колебаний суточного прироста. Таким образом, установлена временна́я дискретность морфогенеза побегов, обусловленная полной или частичной реализацией сезонного цикла развития побегов.
Для полной реализации сезонного цикла развития побегов необходим достаточный объем структурно-энергетических ресурсов саженцев — взаимодействующих развивающихся систем: водоснабжающих корневых и фотосинтезирующих побеговых. Выявленная временна́я дискретность морфогенеза побегов может быть обусловлена нехваткой структурно-энергетических ресурсов, необходимых для выхода из провалов Δ L на восходящие ветви последующих инфрадианных циклов. Образно говоря, черенковым саженцам группы E 2 «не хватило сил» для выхода на третий цикл, а всем остальным черенковым саженцам «не хватило сил» для выхода на второй цикл. В пользу этой гипотезы свидетельствует тот факт, что в контрольной группе сеянцев S с 4-летней корневой системой полностью реализован сезонный цикл развития побегов, включающий четыре инфрадианных цикла.
На рано завершивших рост побегах экспериментальных клонов нехватка структурноэнергетических ресурсов может быть обусловлена как межклоновыми, так и внутриклоновыми различиями. Вероятно, в первом случае продолжительность морфогенеза контролируется генетическими факторами, а во втором случае — эпигенетическими, в частности, процессами ризогенеза.
Для верификации выдвинутых гипотез нами разработаны схемы двух популяционноклоновых экспериментов: по сравнению морфогенеза побегов S. triandra в разновозрастных посадках и по сравнению морфогенеза побегов на одновозрастных саженцах, выращенных из черенков, заготовленных из разных частей порослевых побегов. Выполнение экспериментов запланировано на 2021 г.
Список литературы Поливариантность морфогенеза побегов в клонах Salix triandra (Salicaceae) на фоне периодичности ливневых осадков
- Skvortsov A. K. Willows of Russia and adjacent countries. Taxonomical and geographical revision (English translation with additions). 1999.
- Chase M. W., Christenhusz M. J. M., Fay M. F., Byng J. W., Judd W. S., Soltis D. E.,.. Stevens P. F. An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG IV // Botanical Journal of the Linnean Society. 2016. V. 181. №1. P. 1-20. DOI: 10.1111/boj.12385
- Баркалов В. Ю., Козыренко М. М. Филогенетические отношения видов Salix L. subg. Salix (Salicaceae) по данным секвенирования межгенных спейсеров хлоропластного генома и ITS ядерной рибосомальной ДНК // Генетика. 2014. Т. 50. №8. С. 940-949. DOI: 10.7868/S0016675814070030
- Wu D., Wang Y., Zhang L., Dou L., Gao L. The complete chloroplast genome and phylogenetic analysis of Salix triandra from China // Mitochondrial DNA Part B. 2019. V. 4. №2. P. 3571-3572. DOI: 10.1080/23802359.2019.1674743
- Macalpine W. J., Shield I. F., Trybush S. O., Hayes C. M., Karp A. Overcoming barriers to crossing in willow (Salix spp.) breeding // Asp Appl Biol. 2008. V. 90. P. 173-180. http://hdl.handle.net/2160/10296
- Афонин А. А., Самошкин Е. Н. Зависимость размеров пыльцевых зерен ив трехтычинковой и пепельной от числа хромосом в клетках // Известия высших учебных заведений. Лесной журнал. 1999. №4. С. 35-40.
- Li W., Wu H., Li X., Chen Y., Yin T. Fine mapping of the sex locus in Salix triandra confirms a consistent sex determination mechanism in genus Salix // Horticulture Research. 2020. V. 7. №1. P. 1-12.
- DOI: 10.1038/s41438-020-0289-1
- Szczukowski S., Stolarski M., Tworkowski J., Przyborowski J., Klasa A. Productivity of willow coppice plants grown in short rotations // Plant Soil and Environment. 2005. V. 51. №9. P. 423.
- Морозов И. Р. Определитель ив и их культура. М.: Лесная промышленность, 1966. 254 с.
- Da Cunha A. C. B., Sabedot S., Sampaio C. H., Ramos C. G., da Silva A. R. Salix rubens and Salix triandra species as phytoremediators of soil contaminated with petroleum-derived hydrocarbons // Water, Air, & Soil Pollution. 2012. V. 223. №8. P. 4723-4731.
- DOI: 10.1007/s11270-012-1228-z
- Суханова Л. В. Размножение нектаропродуктивной ивы трехтычинковой черенками // Известия высших учебных заведений. Лесной журнал. 2004. №4. С. 16-22.
- Санникова Е. Г., Попова О. И., Компанцева Е. В. Ива трехтычинковая (Salix triandra L.) - перспективы и возможности использования в медицине и фармации // Фармация и фармакология. 2018. №6 (4). С. 318-339.
- DOI: 10.19163/2307-9266-2018-6-4-318-339
- Rather T. A., Qaisar K. N., Raja T. A., Khan M. A. Growth and productivity of wicker willow (Salix triandra L.) plantation in Kashmir // J. Res. Dev. 2009. V. 9. P. 30.
- Donnelly I., McDonnell K., Finnan J. Novel Approaches to Optimise Early Growth in Willow Crops // Agriculture. 2019. V. 9. №6. P. 116.
- DOI: 10.3390/agriculture9060116
- Епанчинцева О. В., Тишкина Е. А., Мищихина Ю. Д. Динамика прироста ив при использовании различных агротехнических приемов // Известия Оренбургского государственного аграрного университета. 2020. №4 (84). С. 97-103.
- DOI: 10.37670/2073-0853-2020-84-4-97-103
- Сукачев В. Н. О некоторых новых засухоустойчивых ивах // Доклады АН СССР. 1952. Т. 84. №2. С. 67-69.
- Zhao M., Wang W., Wang Z., Chen L., Ma Z., Wang Q. Water use of Salix in the variably unsaturated zone of a semiarid desert region based on in-situ observation // Journal of Hydrology. 2020. V. 591. P. 125579.
- DOI: 10.1016/j.jhydrol.2020.125579
- Zhu Y., Wang G., Li R. Seasonal dynamics of water use strategy of two Salix shrubs in alpine sandy land, Tibetan Plateau // Plos one. 2016. V. 11. №5. P. e0156586.
- DOI: 10.1371/journal.pone.0156586
- Fabio E. S., Leary C. J., Smart L. B. Tolerance of novel inter-specific shrub willow hybrids to water stress // Trees. 2019. V. 33. №4. P. 1015-1026.
- DOI: 10.1007/s00468-019-01835-4
- Pezeshki S. R., Anderson P. H., Shields F. D. Effects of soil moisture regimes on growth and survival of black willow (Salix nigra) posts (cuttings) // Wetlands. 1998. V. 18. №3. P. 460-470.
- DOI: 10.1007/BF03161538
- Verwijst T., Lundkvist A., Edelfeldt S., Forkman J., Nordh N. E. Effects of clone and cutting traits on shoot emergence and early growth of willow // Biomass and Bioenergy. 2012. V. 37. P. 257-264.
- DOI: 10.1016/j.biombioe.2011.12.004
- Fabio E. S., Volk T. A., Miller R. O., Serapiglia M. J., Kemanian A. R., Montes F.,.. Smart L. B. Contributions of environment and genotype to variation in shrub willow biomass composition // Industrial Crops and Products. 2017. V. 108. P. 149-161.
- DOI: 10.1016/j.indcrop.2017.06.030
- Miller R. O. Shrub Willow (Salix) Biomass Crop Performance on Five Sites Over Two Rotations in Michigan, USA and the Implications of Adequate Field Testing to Commercial Producers // BioEnergy Research. 2018. V. 11. №4. P. 803-815.
- DOI: 10.1007/s12155-018-9939-7
- Welc M., Lundkvist A., Verwijst T. Effects of Propagule Phenology (Non-Dormant Versus Dormant) and Planting System (Vertical Versus Horizontal) on Growth Performance of Willow Clones Grown Under Different Weeding Regimes // BioEnergy Research. 2018. V. 11. №3. P. 703-714.
- DOI: 10.1007/s12155-018-9929-9
- Rogers E. R., Zalesny R. S., Hallett R. A., Headlee W. L., Wiese A. H. Relationships among Root-Shoot Ratio, Early Growth, and Health of Hybrid Poplar and Willow Clones Grown in Different Landfill Soils // Forests. 2019. V. 10. №1. P. 49.
- DOI: 10.3390/f10010049
- Работнов Т. А. Жизненный цикл многолетних травянистых растений в луговых ценозах // Тр. БИН АН СССР. Сер. 3. Геоботаника. 1950. №6. С. 7-204.
- Комаров А. С., Зубкова Е. В., Фролов В П. Клеточно-автоматная модель динамики популяций и сообществ кустарничков // Сибирский лесной журнал. 2015. №3. С. 57-69.
- DOI: 10.15372/SJFS20150306
- Жукова Л. А., Комаров А. С. Поливариантность онтогенеза и динамика ценопопуляций растений // Журнал общей биологии. 1990. Т. 51. №4. С.450-461.
- Озернюк Н. Д. Разнообразие Онтогенезов: иерархия механизмов // Онтогенез. 2010. Т.41. №5. С. 323-324.
- Fuchylo Ya. D., Afonin A. A., Sbytna M. V. Selection bases of Developing new varieties of willow family (Salicaceae Mirb.) to create energy plantations // Plant Varieties Studying and Protection. 2016. №4 (33). P. 18-25. (in Ukrainian).
- DOI: 10.21498/2518-1017.4(33).2016.88607
- Афонин А.А. Структурный анализ ритмов развития однолетних побегов ивы трехтычинковой // Бюллетень науки и практики. 2019. Т. 5. №1. С. 22-32.
- Погода и климат. Климатический монитор. http://www.pogodaiklimat.ru/monitor
- Уразалиев К. Р. Гаплоидные технологии в селекции растений // Биотехнология. Теория и практика. 2015. №3. С. 33-44.
- DOI: 10.11134/btp.3.2015.4
- Петров Д. Ф. Генетически регулируемый апомиксис. Новосибирск: Наука, 1964. 189 с.
