Поражение заразихой (Orobanche сumana Wallr.) образцов подсолнечника коллекции ВИР и молекулярное дифференцирование контрастных по устойчивости растений
 образцов подсолнечника коллекции ВИР и молекулярное дифференцирование контрастных по устойчивости растений")
Автор: Челюстникова Т.А., Гучетль С.З., Арасланова Н.М., Антонова Т.С.
Рубрика: Защита растений и иммунология
Статья в выпуске: 2 (148-149), 2011 года.
Бесплатный доступ
Выполнена оценка устойчивости 500 образцов культурного подсолнечника из мировой коллекции ВИР к высоковирулентной популяции заразихи из Тацинского района Ростовской области. Полностью устойчивых образцов не обнаружено, однако они дифференцированы на группы с разной степенью пораже-ния. Выделены образцы с распределением тестированных растений на восприимчивые и полностью устой-чивые. ПЦР анализ ДНК показал генетическую неоднородность устойчивых и восприимчивых растений. Определены локусы, полиморфные для этих групп растений.
Подсолнечник, заразиха, устойчивость, полиморфизм
Короткий адрес: https://sciup.org/142151005
IDR: 142151005 | УДК: 633.854.78:631.52
Affection of sunflower samples of VIR collection by broomrape (Orobanche сumana Wallr.) and molecular differentiation of plants, having contrasts in resistance
An evaluation of stability of 500 samples of cultivated sunflower of world collection of VIR to high-virulent population of broomrape from Tatsinskiy area of Rostov region was made. Entirely resistant samples were not found, however, they were differentiated to groups with different degree of affection. The samples with distribution of trialed plants for susceptible and entirely resistant ones were defined. PCR-analysis of DNA had shown genetic in-homogeneity of resistant and susceptible plants. The loci, polymorphic for these groups of plants, were defined.
Текст научной статьи Поражение заразихой (Orobanche сumana Wallr.) образцов подсолнечника коллекции ВИР и молекулярное дифференцирование контрастных по устойчивости растений
Подсолнечник – один из значимых источников растительного масла в мире и основной – в Российской Федерации. В настоящее время мощным ограничивающим фактором в производстве подсолнечника в Ростовской и Волгоградской областях, а также в ряде районов Ставропольского и Краснодарского краев является заразиха ( Orobanche сumana Wallr.). Заразиха – паразитическое цветковое растение, не содержащее хлорофилла, использует воду, а также органические и минеральные вещества растения-хозяина, обладает высоким потенциалом воспроизводства. Поражение подсолнечника заразихой значительно снижает урожай, а многочисленные осыпающиеся семена засоряют почву, сохраняя всхожесть до 20 лет, ухудшая тем самым состояние посевов в последующие годы.
Основным способом борьбы с этим злостным сорняком является поиск источников генетической устойчивости к новым вирулентным расам паразита и вовлечение их в селекционные программы. Более высока вероятность обнаружения генов устойчивости в коллекциях дикорастущих видов, однако существует некоторая устойчивость к заразихе и у культурного подсолнечника [1; 2].
Использование молекулярных технологий позволяет избежать фенотипических ошибок при отборах, дает возможность сократить сроки для создания материала с генами устойчивости к новым расам заразихи и таким образом ускорить селекционный процесс. Молекулярные маркеры, сцепленные с генами устойчивости к заразихе, могут стать полезным инструментом для контроля переноса генов и быстрой идентификации индивидов с интересующим селекционеров признаком [1; 3; 4].
Tang и соавторы (2003) идентифицировали молекулярные маркеры, сцепленные с геном Or5 , определили положение этого локуса на молекулярной генетической карте подсолнечника [3]. Авторы предполагают, что эта область ответственна у подсолнечника за генетическую устойчивость к заразихе расы Е. Lu и соавторы (2000) идентифицировали пять SCAR (sequence characterized amplified region) маркеров в одной группе сцепления с геном Or5 . Предложенная группа маркеров может быть использована для проведения массового сегрегационного анализа, для насыщения зоны, приближенной к гену Or5, новыми молекулярными маркерами [4]. Солоденко с соавторами, (2003) выявили полиморфные фрагменты амплификации ДНК, дифференцирующие устойчивые и восприимчивые к заразихе растения, определили сцепление молекулярного маркера и локуса Or3 [5]. Используя опыт и результаты исследований, проведенных в мире, мы начали молекулярное дифференцирование устойчивых и восприимчивых растений подсолнечника к заразихе, собранной в регионах РФ.
Целью нашей работы являлась оценка устойчивости образцов подсолнечника коллекции ВИР к заразихе и поиск молекулярных различий между устойчивыми и восприимчивыми растениями.
Материалы и методы. Для выявления устойчивых к заразихе генотипов использованы 500 образцов мировой коллекции культурного подсолнечника ВИР и испанская линия Р 96, устойчивая к расам заразихи от А до F.
Семена подсолнечника сеяли в короба, заполненные смесью просеянной почвы с речным песком в соотношении 3:1. В почвенную смесь вносили семена заразихи, собранные в Тацинском районе Ростовской области, из расчета 200 мг семян на 1 кг почвы. По ранее опубликованным данным, в этой популяции заразихи высока концентрация высоковирулентных рас E, F,G [6].
Растения подсолнечника выращивали в теплице в течение 30 дней при температуре 22-25°С при 16 часовом фотопериоде. Через 30 дней после появления всходов проводили их оценку на устойчивость к заразихе. Восприимчивыми считались растения, на корнях которых были обнаружены клубеньки заразихи или сформировавшиеся побеги; устойчивыми – растения, на корнях которых не было обнаружено здоровых клубеньков и побегов, но были видны многочисленные некрозы клеток в области проникновения проростков заразихи и погибшие клубеньки. Учитывали процент пораженных растений и степень поражения. Степень поражения учитывали как среднее количество клубеньков и цветоносов паразита на одно поражённое растение. В качестве контроля был использован сорт ВНИИМК 8883, не обладающий устойчивостью к современным расам O. сumana .
Для молекулярного анализа отбирали по два настоящих листа с растения. Выделение ДНК проводили по модифицированному методу Saghai-Maroof et аl. [7]. Ткани листьев гомогенизировали с 0,9 мл СТАВ буфера, в течение часа инкубировали при температуре 65 оС. Затем добавляли половину объема смеси хлороформа и изоамилового спирта (24 : 1) и тщательно перемешивали с последующим центрифугированием. Очистку хлороформом проводили дважды. Осаждение ДНК проводили охлажденным до -20 оС изопропаноловым спиртом. Подсушенную ДНК растворяли в течение 12 часов в стерильной воде. Концентрацию ДНК в полученном препарате определяли по интенсивности свечения пробы объемом 10 мкл в ультрафиолетовом свете в 1 % -ном агарозном геле с содержанием бромистого этидия.
Для проведения полимеразной цепной реакции использовали 25 мкл реакционной смеси следующего состава: 67 мМ трис-HCl, рН 8,8; 16,6 мM сульфата аммония; 2,5 мM MgCl2; 0,01 % Tween 20; по 0,2 мM дезоксирибонуклеозид фосфатов; по 10 пМ прайме- ров; 10 нг матричной ДНК и 1 ед. рекомбинантной термостабильной ДНК полимеразы (Москва, ГосНИИгенетика). Амплификацию проводили в приборе Терцик (ДНК-технология, Россия). Для амплификации ДНК применяли нуклеотидные последовательности пяти пар SCAR-праймеров, сконструированных [4] (см. табл. 2). Термальный режим реакций подбирали для каждой пары праймеров с учетом их нуклеотидного состава. Для большинства проведенных реакций оптимальным оказался режим амплификации с начальной денатурацией при 96 оС в течение 2 мин, затем 30 циклов при соблюдении тем-пературно-времен-ного режима: отжиг при 60-65 оС в течение 40 сек, элонгация – 1 мин при 70 оС, денатурация при 94 оС – 30 сек, финальная элонгация – 2 мин.
Электрофоретическое разделение продуктов амплификации проводили в агарозном геле (2 % агароза, 1×ТАЕ-буфер) с использованием камеры для горизонтального электрофореза (SE.1, ДНК-технология, Россия) в течение 1-1,5 часов при силе тока 50 mA, напряжении 90-100 V. Документирование результатов электрофореза обеспечивалось при помощи гель-документирующей видеосистемы BIO-PRINT (Vilber Lourmat, Франция).
Результаты и обсуждение. Расовый состав популяций заразихи динамичен, находится в зависимости от преобладающего сортимента подсолнечника в регионе. Задачей селекционеров является поиск источников устойчивости к новым вирулентным расам и создание сортов и гибридов с привнесением в них генов устойчивости. В связи с этим и проведена оценка устойчивости образцов культурного подсолнечника мировой коллекции ВИР, различающихся по происхождению. Результаты приведены в таблице 1. Большая часть тестированного материала в разной степени, но стопроцентно поражалась заразихой. По степени поражения образцы коллекции ВИР условно можно разделить на три группы. Первая группа включает образцы со степенью поражения на уровне контроля. В нее вошли образцы: 2982 (из Испании), 2925 (из Франции), 2995 (из Португалии), 3020 (из Венгрии), 3074 и 3080 (из Мексики). Среднее количество клубеньков и побегов заразихи на одно поражённое растение у этих образцов было в пределах от 96 до 121. Вторая группа со средней степенью поражения от 58 до73 включает образцы: 2930 (из
Франции), 2975 (из Австралии), 2978 (из Испании), 2988 (из Болгарии), 3004 (из Югославии), 3015 (из Венгрии).
Таблица 1
Поражение некоторых образцов подсолнечника коллекции ВИР заразихой (O. сumana) при искусственном заражении в теплице
|
№ п/п |
№ каталога |
Происхождение |
Учетных расте-ний, шт. |
Пораженных растений, % |
Степень поражения |
|
|
1 |
2925 |
Франция |
30 |
100 |
110 |
|
|
2 |
2930 |
Франция |
30 |
100 |
61 |
|
|
3 |
2931 |
Франция |
31 |
100 |
20 |
|
|
4 |
2938 |
Аргентина |
34 |
100 |
12 |
|
|
5 |
2953 |
Аргентина |
30 |
89 |
11 |
|
|
6 |
2954 |
Аргентина |
30 |
82 |
12 |
|
|
7 |
2973 |
Австралия |
30 |
90 |
28 |
|
|
8 |
2975 |
Австралия |
30 |
100 |
73 |
|
|
9 |
2978 |
Испания |
30 |
100 |
62 |
|
|
10 |
2982 |
Испания |
31 |
100 |
121 |
|
|
11 |
2988 |
Болгария |
30 |
100 |
58 |
|
|
12 |
2995 |
Португалия |
30 |
100 |
96 |
|
|
13 |
2996 |
Португалия |
31 |
100 |
89 |
|
|
14 |
3004 |
Югославия |
30 |
100 |
68 |
|
|
15 |
3015 |
Венгрия |
29 |
100 |
65 |
|
|
16 |
3020 |
Венгрия |
30 |
100 |
98 |
|
|
17 |
3074 |
Мексика |
30 |
100 |
97 |
|
|
18 |
3080 |
Мексика |
30 |
100 |
115 |
|
|
19 |
Р 96 |
Испания |
30 |
78 |
12 |
|
|
19 |
ВНИИМК 8883, контроль |
30 |
100 |
115 |
||
Особенно интересна третья группа образцов, степень поражения которых была в пределах от 12 до 28 экземпляров заразихи на растение. Она включает образцы: 2931 (из Франции), 2973 (из Австралии), 2938, 2953 и 2954 (из Аргентины) и линию Р 96, созданную в Испании, не входящую в коллекцию ВИР и обладающую устойчивостью к расам от А до F. Среди тестированных образцов подсолнечника 2973, 2953, 2954 и Р 96 наблюдалось расщепление на непораженные и поражённые в слабой степени экземпляры.
Для выяснения причины различий по устойчивости к заразихе среди учетных растений внутри образцов, выполнено сравнение ДНК, выделенной отдельно из пяти пораженных и пяти непораженных растений образцов 2953, 2954 и линии Р 96. В нашей работе с популяцией заразихи, состоящей из смеси рас, учитывалось известное предположение [5], что гены устойчивости к ней расположены кластерно, подобно генам устойчивости к ложной мучнистой росе – Pl 1, Pl 2 и Pl 6, формирующим кластер и сцепленным с одними и теми же ДНК маркерами. Поэтому были использованы 5 локусов ДНК (табл. 2), предложенные Lu et al. (2000) [4], как характеризующие устойчивость к расе Е заразихи.
Таблица 2
Характеристика SCAR маркеров, использованных для дифференциации контрастных по устойчивости к заразихе растений подсолнечника
ВНИИМК 2011 г.
|
Назва ние локусов |
Последовательность фланкирующих праймеров 5’-3’ |
Полиморфизм в образцах |
||
|
Р 96 |
2953 |
2954 |
||
|
RTS 05 |
TGGTCGCAGATGGACGTGTGGGTG GTCGCAGAGAGTGAGAGAGAGTGT |
+ |
+ |
+ |
|
RTS28 |
AGTAGACGGGCAAAGCGAAAGGAT AGTAGACGGGTTGAATATGTTGAA |
- |
+ |
+ |
|
RTS29 |
GCTTCCCCTTAATGATCCGGAAGA GCTTCCCCTTGGCTAGAAGATGAA |
- |
- |
- |
|
RTS40 |
TCCACCGAGCTACCAGTTCCGGAG TCCACCGAGCGAGCATATTCCGAG |
- |
- |
- |
|
RTS43 |
CGGAGAGCGAGCATAGGGTCAGGT CGGAGAGCGACGACATCATTCCAA |
- |
- |
- |
|
( + ) - есть полиморфизм между пораженными и непораженными растениями; (- ) – нет полиморфизма между пораженными и непораженными растениями |
||||
При проведении амплификации ДНК по приведенным в таблице 2 пяти локусам получены следующие результаты. По локусам RTS29, RTS40 и RTS43 продукты амплификации ДНК у пораженных и непораженных растений не отличались. Отличия в ампликонах получены по локусам RTS05 и RTS28. По локусу RTS28 у пораженных и непораженных растений линии Р 96 амплифицированы одинаковые фрагменты ДНК. У непораженных растений образцов 2953 и 2954 обнаружены более интенсивные фракции и, возможно, при использовании других методов разделения продуктов реакции будут визуализированы различия по количеству фрагментов. Продукты амплификации ДНК по локусу RTS05 различали пораженные и непораженные растения у образцов 2953, 2954 и линии Р 96 (рисунок). У восприимчивых растений присутствовала фракция ДНК 650 п.н. Это указывает на генетическую неоднородность внутри указанных образцов, различающихся и по устойчивости к вирулентным расам заразихи. Поэтому локус RTS 05 можно в дальнейшем использовать как маркерный при анализе растений, устойчивых и восприимчивых к высоковирулентным расам заразихи.
Выводы . Среди 500 образцов культурного подсолнечника коллекции ВИР не выявлено полностью устойчивых к высоковирулентной популяции заразихи из Тацинского-района Ростовской области. Однако образцы различались по степени поражения. Обнару-
ISSN 0202-5493. МАСЛИЧНЫЕ КУЛЬТУРЫ. Научно-технический бюллетень Всероссийского научно-исследовательского института масличных культур. Вып.2 (148-149), 2011
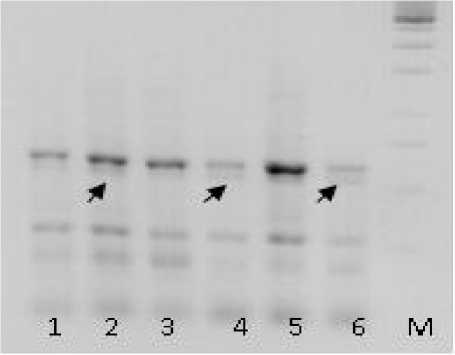
Рисунок – Электрофоретические спектры продуктов амплификации ДНК у устойчивых и восприимчивых растений подсолнечника по локусу RTS 05.
Дорожки: 1, 3, 5 – устойчивые растения; 2, 4, 6 – восприимчивые растения; образцы: 1, 2 – Р 96; 3, 4 – 2953; 5, 6 – 2954; М – маркер молекулярного веса 1 кb; стрелкой указана полиморфная зона 650 п.н.
-
3. Tang, S. Genetic mapping of the Or5 gene for resistance to Orobanche race E in sunflower / S. Tang, A. Heesacker, V.K. Kishore, A. Fernandez, E. Sadik, G. Cole, St. Knapp. // Crop. Sci. – 2003. –V.43. – P. 1021-1028.
-
4. Lu, Y.H. Development of SCAR markers linked to the gene Or5 conferring resistance to broomrape ( Orobanche cumana Wallr.) in sunflower / Y.H. Lu, J.M. Melero-Vara, J.A. Blanchard // TAG. – 2000. – 100. – P. 626-632.
-
5. Солоденко, А.Е. Маркирование гена устойчивости к заразихе Or3 у подсолнечника / А.Е. Солоденко, А. В. Саналатий, В.В. Толмачев, К.В. Ведмедева, Ю.М. Сиволап // Цитология и генетика. – 2005. – С. 9-12.
-
6. Антонова, Т.С. Вирулентность заразихи, поражающей подсолнечник в Волгоградской и Ростовской областях / Т.С. Антонова, Н.М. Арасланова, С.А. Рамазанова, С.З. Гучетль, Т.А. Челюстникова // Масличные культуры: Науч.-тех. бюл. ВНИИМК. – 2009. – Вып.1 (140). – С. 31-37.
-
7. Saghai-Maroof, M.A. Ribosomal DNA spacer-length polymorphisms in barley: Mende-lian inheritance, chromosomal location, and population dynamics / M.A. Saghai-Maroof, K.M. Soliman, R.A. Jorgensen, R.W. Allard // PNAS USA. – 1984. – 81. – P. 8014-8018.
жено четыре образца с разделением на устойчивые и восприимчивые растения. Методом ПЦР ДНК по двум SCAR локусам найдены генетические отличия у пораженных и непораженных растений внутри образцов. Локус RTS 05 пригоден для использования в качестве маркерного при анализе растений, устойчивых и восприимчивых к высоковирулентным расам.
Исследования выполнены при финансовой поддержке РФФИ, грант № 11-04-96502.
В.А. Гавриловой и к. с-х. н. В.Т. Рожковой за любезно предоставленный семенной материал подсолнечника из коллекции ВИР.
