Повышение яйценоскости у высокопродуктивных кур и проблема ее раннего прогнозирования

Автор: Штеле А.Л.
Журнал: Сельскохозяйственная биология @agrobiology
Рубрика: Обзоры, проблемы, итоги
Статья в выпуске: 6 т.49, 2014 года.
Бесплатный доступ
Процессы созревания первичной яйцеклетки в естественной взаимосвязи с желтком в яичнике, образование белка и скорлупы в яйцеводе, овуляция и цикл яйцекладки детерминированы геномом и регулируются на нейрогуморальном (нейрогормональном) уровне, реализуясь в системе генотип-среда, что в итоге и определяет яичную продуктивность птицы. Продуктивность лимитируется длительностью созревания яйцеклетки и образования желтка, а также снижением репродуктивного потенциала и качества яиц с возрастом кур и(или) вследствие влияния технологических факторов. Наблюдаемые фенотипические изменения при селекционном давлении в условиях современных технологий свидетельствуют о расширении физиологических границ проявления признаков продуктивности (яйценоскости, массы и качества яиц) при интенсивном метаболизме. У гибридных кур современных яичных кроссов в течение длительного периода продуктивного использования возможна ежесуточная овуляция яйцеклетки-желтка при синхронизации процессов образования и снесения полноценного яйца. Яйценоскость кур можно прогнозировать в постэмбриональном онтогенезе по числу крупных фолликулов, созревающих в яичнике, при условии достижения 85-90 % яйценоскости в начальные 5-6 нед продуктивного периода. Немаловажное значение имеет также устойчивый цикл яйцекладки с небольшим интервалом (1-2 сут). Быстро формирующиеся в яичнике 5-6 крупных фолликулов при совпадающем недельном периоде созревания яйцеклетки-желтка в овогенезе определяют биологический потенциал функционирования репродуктивных органов у высокопродуктивных кур. Использование этих показателей при оценке особи и ее потомства дает возможность на ранних этапах онтогенеза прогнозировать высокую яичную продуктивность, что, в свою очередь, позволит ускорить селекционные работы при совершенствовании и создании новых яичных кроссов.
Яичная продуктивность, геном кур, яйцеклетка, яичник, овогенез, яйцекладка, полноценность, концепция, прогнозирование
Короткий адрес: https://sciup.org/142133551
IDR: 142133551 | УДК: 636.5.034:636.018 | DOI: 10.15389/agrobiology.2014.6.26rus
Problem of egg productivity in hens and its early prediction
The generalized processes of natural maturation of oocyte with egg yolk in the ovary, production of egg white and eggshell in the oviduct, as well as ovulation and laying cycle are under the genotype control. Besides, they are affected by the environment and regulated by neurohumoral (neurohormonal) system. All these factors determine ultimately the egg productivity in poultry. Thus, it is limited by time, required for the oocyte maturation and yolk synthesis, and also by the decreasing reproductive potency of hens and egg quality because of aging and/or influence of technological factors. Whatever it was, the observed phenotypic variability due to selection pressure and the applied intensive technologies in poultry indicates the physiological limits of productivity (i.e. egg-laying rate, egg weight and quality) to become wider in hens with intensive metabolism. In the modern egg hybrid crosses a daily ovulation occurs synchronously with a mature egg formation and laying during long productive usage. The egg laying chickens can be forecasted in the postembryonic ontogenesis by the number of large follicles which are maturing in the ovary, if 85-90 % egg productivity is reached within 5-6 weeks after the laying begins. The steady cycle of egg laying with small interval (1-2 days) is also of importance. Thus, the rapid formation of 5-6 follicles in the ovary coinciding with weekly maturation of the yolk in ovogenesis determines the biological potency of reproductive function in high-yield laying hens. The use of these parameters for assessing egg chicks and their offspring gives an opportunity to predict the high-producers in the early ontogenesis. It will speed up breeding to improve modern egg crosses, create new ones, and also to optimize the intensive technologies in poultry.
Текст научной статьи Повышение яйценоскости у высокопродуктивных кур и проблема ее раннего прогнозирования
С 1950-х годов в результате длительной и интенсивной селекции и гибридизации годовая яйценоскость кроссбредных кур повысилась почти в 2 раза — до 320-330 шт., а масса яйца составила 60-65 г против 55-60 г. При этом в основном увеличилась масса белка и скорлупы, тогда как масса желтка в селекционных линиях яичных кроссов стабилизирована в пределах 16-17 г, у гибридных кур — в среднем 18 г (1). Следовательно, яичная продуктивность кур приблизилась к биологическому пределу.
Сложные процессы естественного созревания яйцеклетки с желтком в яичнике, образование белка и скорлупы в яйцеводе, овуляция и цикл яйцекладки детерминированы генетически, регулируются на нейрогуморальном (нейрогормональном) уровне и зависят от внешних условий. Поэтому яичная продуктивность лимитируется двумя основными факторами — длительностью созревания яйцеклетки и образования желтка и снижением репродуктивного потенциала и качества яиц с возрастом кур и(или) вследствие влияния технологических факторов.
В настоящее время активизируются исследования структуры и функциональной активности генома у высокопродуктивных кур, на основе чего разрабатываются современные методы селекции и оптимизируются технологии промышленного содержания поголовья. В связи с этим проблема раннего прогнозирования яичной продуктивности приобретает особую актуальность.
Генетические факторы продуктивности кур и селекция . Селекция и гибридизация с использованием достижений популяционной генетики составляют основу племенной работы с сельскохозяйственной птицей. Внутрилинейная селекции и строгий отбор по важнейшим хозяйственным признакам при скрещивании сочетающихся линий обеспечивают высокую продуктивность у гибридных кур яичных кроссов. Благодаря генетическому разнообразию оказалось возможным поддерживать изменчивость полигенных признаков и постоянно повышать продуктивность кур. Методы селекции дополняются ДНК-технологиями на основании использования генетического полиморфизма и многофункциональных молекулярных маркеров для раннего прогнозирования продуктивности (1).
Расшифровка гаплоидного генома кур выявила 24 тыс. генов и более 1 млрд пар нуклеотидов в полиморфных участках ДНК, отражающих генетическое разнообразие. SNPs (single nucleotide polymorphism) представляют собой основу при маркерной селекции (2). Этот подход особенно результативен в случае хозяйственно полезных признаков, изменчивость которых сцеплена с половыми хромосомами. Раннее тестирование и отбор петушков в селекционных (закрытых) линиях и родственных группах служит эффективным приемом в селекции и сокращает время отбора в поколениях.
Для генома кур описано около 3 млн SNPs, что дает возможность установить местонахождение маркеров для полигенных признаков продуктивности (3). Это позволяет идентифицировать гены структурных белков, регуляторов множественного действия, а также гены-маркеры, локализованные на участке ДНК, и использовать их при картировании хромосом и в селекционной работе. Описано создание чипов для 600 тыс. нуклеотидных последовательностей с идентификацией соответствующих генов в коммерческих линиях (4). Полная расшифровка генома кур облегчит выделение полиморфных участков ДНК, маркирование значимых нуклеотидных последовательностей, а на хромосомных картах — структурных ге нов и локусов сцепления, определяющих наследование признаков продуктивности. Последний известный вариант интегрированной карты хромосом у кур составлен с использованием цитогенетического картирования, групп сцепления на отдельных хромосомах, а также полиморфных маркерных последовательностей. Эта версия включает 1889 локусов, из которых 450
объединены в 50 групп сцепления (5).
2*1 2*2 23 24 25 26 27 28 29 30 3*1 32 33 34 3* 36 37 з" Z W
Рис. 1. Схематическое изображение кариотипа домашней курицы Gallus gallus с гаплоидным набором хромосом и двумя половыми хромосомами (38) .
Геномное сканирование в поисках локусов количественных признаков (QTL) выявило около 29 локусов, локализованных в пяти из восьми известных макрохромосом курицы — 2-й, 3-й, 4-й, 5-й, 8-й и в половой Z-хромосоме (рис. 1). При их генетическом картировании выявлены локусы сцепления генов для массы тела, раз мера яиц, конверсии корма, других признаков продуктивности. На все восемь макрохромосом и Z-хромосому приходится более 70 % нуклеотидных последовательностей генома, на микрохромосомы — оставшаяся доля.
Для микрохромосом установлена генетическая активность, а также многие косвенные признаки повышенной плотности локализации генов (6).
Половая хромосома курицы Z (или X) содержит гены, которые влияют на некоторые признаки продуктивности. На этой основе для яичных и мясных кроссов созданы гомозиготные родительские формы, ау-тосексные по окраске оперения (аллели К и к ) и скорости формирования перьевого покрова (аллели S и s ), у которых 1-суточные цыплята различаются по соответствующему признаку в зависимости от пола. Ген карликовости ( dw / DW) был использован при выведении материнской линии (с пониженной живой массой и повышенной яйценоскостью) для мясного кросса Hubbard ISA F15 (7).
При прогнозировании яичной продуктивности представляют интерес аллельные гены половой хромосомы Z, контролирующие ограниченную ( ro ) и нормальную ( RO ) овуляцию. В эксперименте отмечено, что куры-несушки, гомозиготные по аллелям гена ro , в ряде случаев не способны к овуляции яйцеклетки-желтка из-за точечной мутации. Это связано с недостаточным транспортом липопротеинов из печени в фолликулы яичника при образовании желтка (8). В другой работе установлено, что гены ограниченной овуляции воздействуют на увеличение размера некоторых костей крыльев и ног и содержания в них минеральных веществ. Это способствует повышению массы тела по сравнению с той, которая присуща высокопродуктивной птице (9).
Селекционная фирма «Hy-Line International» (США), имеющая собственную генетическую лабораторию и необходимую производственную базу, в течение последнего времени применяет методы геномной селекции по генам-маркерам, связанным с хозяйственно важными признаками и ранним прогнозированием продуктивности. Селекция в линиях при создании кроссов ведется на стабильно высокую пожизненную продуктивность гибридных кур (80 нед и более), устойчивость к стрессам и заболеваниям .
Овогенез у высокопродуктивных кур. Морфогенез репродуктивных органов (яичник и яйцевод), развитие и созревание первичной яйцеклетки, образование полноценного яйца и половая скороспелость курочек связаны с периодами их роста и развития. За последние 6070 лет интенсивной селекции возраст снесения первого яйца (половая скороспелость) у яичных кур снизился в 1,5 раза — с 6 мес (26-27 нед) до 4 мес (17-18 нед). Это, однако, сопровождалось уменьшением прочности костей ног и крыльев, отмечаемым у курочек в постэмбриональный период. Проблема поддержания нормального развития пока решается за счет селекции на крепость костяка в сочетании с ограниченным кормлением, а также контролируемой живой массой к началу яйцекладки (1,2-1,3 кг) и в течение продуктивного периода у несушек (10).
Некоторые сдвиги в функциональном развитии яичника отмечаются у гибридных кур еще в генеративную фазу овогенеза, что показано на примере кросса Loghmann Brown. Зачатки гонад у куриных эмбрионов появляются в 4-суточном возрасте, а дифференциация гонад на яичники и семенники начинается с 8-х сут. С 11-суточного возраста происходит превращение первичных зародышевых клеток в оогонии, численность которых при быстром митотическом делении достигает нескольких сот тысяч. К 19-м сут большая часть оогониев погибает, и лишь некоторые из них дифференцируются в ооциты, вокруг которых начинают формироваться фолликулы (11).
До вывода цыплят фолликулярные стадии генеративных элемен- тов (ооциты) становятся преобладающими и завершаются образованием первичных яйцеклеток. В 1-суточном возрасте в яичнике курочек насчитывают 3,5-4,0 тыс. первичных ооцитов (яйцеклеток), диаметр которых составляет 0,01-0,02 мм при общей массе 30-50 мг. Благодаря фолликулу осуществляется взаимосвязь между развивающимся ооцитом и яичником. Первичная яйцеклетка (без желтка) содержит ядро с крупным ядрышком и диплоидным набором хромосом. Непосредственно к ооциту прилегает желточная (вителлиновая) оболочка — уплотненный слой цитоплазмы (12, 13).
Рост и созревание первичной яйцеклетки в вегетативную фазу проходит в фолликулах яичника вплоть до овуляции зрелой яйцеклетки в яйцевод. В эту фазу овогенеза можно выделить три стадии развития яйцеклетки, сведения о продолжительности которых и соответствующих продуктах представлены в таблице 1. На первой стадии овогенеза (1-6-я нед) наблюдается незначительный рост яйцеклетки за счет цитоплазмы без формирования желточного материала. К этому времени образуется зародышевый диск, отмечается увеличение ядра и цитоплазмы, ее диаметр достигает 0,5-1,0 мм.
|
1. Характеристика этапов овогенеза у высокопродуктивных кур |
|||
|
Этап, процесс |
| Объект, орган |
| Продолжительность |
| Продукт овогенеза |
|
Генеративная фаза (эмбриональный период) |
Эмбрион—суточный цыпленок |
8-21 сут |
Первичные зародышевые клетки — оогонии ^ ооциты |
|
Вегетативная фаза (постэмбриональный период) |
Яичник |
От 1 сут до 17 нед |
Ооцит — первичная яйцеклетка ^ зрелая яйцеклетка |
|
Рост яйцеклетки |
Яичник |
От 1 сут до 6 нед |
Яйцеклетка с зародышевым диском и цитоплазмой |
|
Развитие яйцеклетки |
Яичник |
7-16 нед |
Яйцеклетка с формирующимся желтком как единая клетка |
|
Созревание яйцеклетки |
Яичник |
16-17 нед |
Зрелая яйцеклетка со сформировавшимся желтком |
|
Овуляция |
Яичник |
В среднем 30 мин |
Половая гамета с гаплоидным набором хромосом, способная к оплодотворению |
|
Продуктивный период онтогенеза у кур-несушек |
Яичник и яйцевод |
18-73 нед и более |
Первичные яйцеклетки на разных стадиях развития (ритмичная овуляция и яйцекладка) |
Вторая, наиболее длительная стадия овогенеза, продолжается от 7-до 16-недельного возраста курочек. Рост и созревание яйцеклеток сопровождается накоплением желточной массы в каждой из них при транспортировке кровотоком из печени в фолликул. В нем питательные вещества накапливаются в виде сложных протеинов (липопротеиды, гликопротеиды), триглицеридов, фосфолипидов, свободного холестерина, других биологически активных соединений.
Третья стадия овогенеза у кур — образование зрелой яйцеклетки (созревание) приходится на 16-17-ю нед. Ускоренное накопление желтка происходит последовательно в нескольких крупных фолликулах яичника и занимает около 1 нед. Период быстрого накопления желтка в яичнике происходит в фолликулах в течение 5-6 сут, где первичная яйцеклетка превращается в зрелую яйцеклетку (вторичный ооцит) и овулирует в яйцевод. Разрыв оболочки фолликула и перемещение зрелой яйцеклетки-желтка в воронку яйцевода (овуляция) занимают 15-30 мин.
Зрелая яйцеклетка в виде бластодиска находится на поверхности желтка и вместе с ним представляет собой одну клетку, окруженную плотной вителлиновой оболочкой. Переход от диплоидного набора хромосом к гаплоидному совершается в результате мейоза, первое деление которого проходит в ядре яйцеклетки до начала овуляции. После перемещения яй- цеклетки-желтка в воронку яйцевода происходит второе деление мейоза и образуется половая гамета с гаплоидным набором хромосом.
Так завершается образование яйцеклетки — половой гаметы, которая благодаря желтку имеет большой запас питательных и биологически активных веществ, необходимых для воспроизводства потомства. Отметим, что каждая яйцеклетка в естественной взаимосвязи с желтком проходит длительный процесс роста и развития в фолликуле яичника от 1-суточного до 16-17-недельного возраста, независимо от того, осуществится ли ее оплодотворение в яйцеводе или нет. С завершением образования яйца (белок и скорлупа) в яйцеводе формируется биологическая полноценность куриного яйца.
Биологическая особенность домашних кур заключается в способности откладывать полноценные яйца без оплодотворения. Именно этот фактор определяет длительность яйцекладки и биологические пределы яичной продуктивности, в равной мере относящиеся к производству оплодотворенных инкубационных яиц (племенное стадо) и неоплодотворенных пищевых яиц (промышленное стадо). Отметим, что у большинства видов класса Aves самки начинают откладывать яйца только после спаривания с самцами и оплодотворения яиц.
Современная концепция образования яиц. После овуляции яйцеклетки по мере продвижения желтка по яйцеводу образуются белок и скорлупа, что завершается снесением полноценного яйца. В яйцеводе яйцо формируется в среднем за 22-24 ч, в том числе в воронке — 20 мин, в белковом отделе — 3 ч, в перешейке — 1 ч, в матке — 18-19 ч. У высокопродуктивных кур образование полноценного яйца (белок, скорлупа) в яйцеводе занимает 22-23 ч. Время формирования скорлупы в яйцеводе составляет 17-18 ч, или около 2/3 суток, что служит вторым по значимости фактором, ограничивающим яичную продуктивность кур (14).
В ранних работах установлено, что максимальная скорость переноса питательных веществ из плазмы крови в фолликул в расчете на единицу его поверхности отмечается при массе ооцита 2,5 г (цит. по 13). Можно считать, что это начальный фолликул в каждой серии быстро созревающих в яичнике. С увеличением поверхности каждого последующего фолликула (до овуляции) скорость отложения желточного материала нарастает вследствие роста фолликулярной оболочки.
У высокопродуктивных кур (более 300 яиц на несушку) выделяется первый большой фолликул (рис. 2, А), который классифицируется как предовуляторный, все последующие фолликулы — по размеру (массе). При представленном на рисунке минимальном числе фолликулов овуляция самого крупного с максимальной массой желтка для этой серии произошла ранее. По данным литературы, масса желтка яиц у гибридных кур в среднем составляет 18 г (15).
При относительно невысокой яйценоскости (210-230 яиц), характерной для современных мясояичных пород и материнских линий мясных кроссов в яичнике одновременно формируются 8-10 и более крупных фолликулов. При таком их числе, особенно во второй половине продуктивного периода, отмечается десинхронизация овуляции и цикла яйцекладки, что служит одной из причин образования двухжелтковых яиц с повышенной массой — 75-80 г и более.
В яичнике линейных кур мясояичной породы белый плимутрок (см. рис. 2, Б) ни один из трех более крупных фолликулов не выделяется по величине, так что любой из них может стать предовуляторным. В по- добном случае также возможна практически одновременная (с небольшим интервалом) овуляция двух фолликулов, что приводит к образованию двухжелткового яйца. Это указывает на десинхронизацию процессов образования яйцеклетки-желтка и овуляции, снесения яйца и цикла яйцекладки. Яйца с двумя желтками могут откладывать в первые недели продуктивного периода молодые куры-несушки, у которых полный цикл образования и снесения яйца еще не установился. О десинхронизации косвенно свидетельствует появление нестандартных яиц с измененной формой и(или) отклонениями по другим морфометрическим показателям.
А
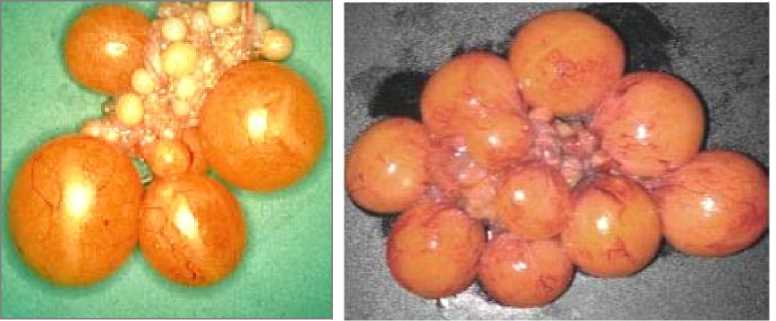
Рис. 2. Крупные фолликулы в яичниках у высокопродуктивных яичных кур (А) и линий мясояичных кур (Б) породы белый плимутрок ( кросс Cobb) (10).
К немаловажным факторам высокой и устойчивой продуктивности кур относится цикл яйцекладки, характерный для каждой несушки. Он определяется временем, в течение которого от курицы без перерыва получают последовательно снесенные яйца. Высокопродуктивная птица имеет длинные циклы яйцекладки, продолжающиеся, как правило, 50-80 сут с небольшим интервалом (1-2 сут) между ними. Частота повторения циклов с интервалами устанавливает ритм яйценоскости в продуктивный период.
Особенность высокопродуктивных яичных кур заключается в быстром накоплении питательных веществ и биологически активных компонентов в 5-6 больших фолликулах, образующихся последовательно, при 67-суточном периоде созревания яйцеклетки. В самом крупном фолликуле масса желтка в наибольшей степени повышается за 24 ч до овуляции, а его диаметр достигает 32-35 мм у молодых кур и 40-42 см у взрослых особей. Число фолликулов в яичнике служит одним из показателей физиологического состояния репродуктивных органов и биологического потенциала яйценоскости кур (16).
Образование белка и скорлупы в яйцеводе синхронизировано с овуляцией яйцеклетки-желтка и циклом яйцекладки, что определяет длительность формирования яйца при разном числе фолликулов в яичнике (рис. 3). Нами предложен коэффициент интенсивности роста фолликула как показатель суточного увеличения его массы. Он рассчитывается по разности прироста массы (18,0-2,5 г), поделенной на число фолликулов в яичнике (см. рис. 3).
При пяти фолликулах в яичнике коэффициент интенсивности роста наибольший и составляет 3,10, при шести — 2,58, семи — 2,21, восьми — 1,94 и при девяти фолликулах — 1,92. При пяти-шести фолликулах, одновременно находящихся в яичнике на стадии созревания яйце-
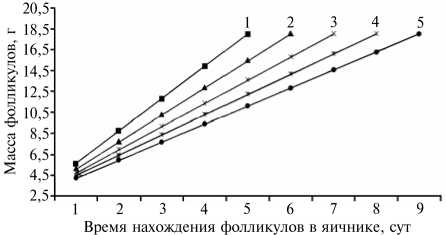
Рис. 3. Расчетная длительность формирования крупных фолликулов в зависимости от их числа в яичнике у кур-несушек: 1-5 — соответственно 5 шт., 6 шт., 7 шт., 8 шт. и 9 шт.
клетки-желтка, отмечается более высокий коэффициент интенсивности прироста их массы. Ранее считалось, что в яичнике у продуктивных кур должно быть семь-девять и более крупных фолликулов, число которых определяет яйценоскость у мясояичных и яичных пород. Таким образом, можно отметить, что чем меньше крупных фолликулов в яичнике (4-5 шт.), в которых последовательно происходит накопление желтка и созревание яйцеклетки, тем быстрее наступает ову-
ляция желтка и тем выше продуктивность кур.
Сложный и длительный процесс роста и созревания яйцеклетки-желтка в яичнике, определяемый геномом, служит главным ограничительным фактором при формировании продуктивности кур в процессе онтогенеза. У высокопродуктивной птицы интенсивный биосинтез питательных веществ и быстрое накопление желточной массы в крупных фолликулах обеспечивают их ускоренное формирование и возможность ежесуточной овуляции яйцеклетки-желтка при суточном ритме яйцекладки. Известный факт снесения курицей породы леггорн 361 полноценного яйца за 364 сут, то есть практически при 100 % яйценоскости, можно считать пределом яичной продуктивности.
Прогнозирование яичной продуктивности. Гибридные куры характеризуются весьма быстрым ростом и развитием, в результате чего в 16-17-недельном возрасте они начинают откладывать первые яйца. При групповом учете половую скороспелость устанавливают при достижении 5 % интенсивности яйценоскости, которая во многом определяет продуктивность кур в дальнейшем. Наследуемость яйценоскости к началу яйцекладки — фактор с наиболее высокой долей влияния на яичную продуктивность.
Функциональная активность яичника и яйцевода, других внутренних органов очень высока, особенно в начале и на пике яйцекладки. В печени происходит интенсивный синтез питательных веществ, биологически активных соединений, входящих в состав желтка, белка и скорлупы, которые через кровь поступают в фолликулы, яичник и яйцевод. При этом также важен возраст половой зрелости, когда особь способна давать потомство. Принято считать, что у высокопродуктивных кур половая зрелость наступает при достижении 50 % яйценоскости в стаде. На примере кроссов фирмы «Hy-Line International» (США) отметим, что за последние 30 лет возраст достижения этого значения снизился почти на 3 нед (17). То же свойственно другим яичным кроссам, что, в свою очередь, определяет быстрое нарастание пика продуктивности (85-90 % и более), присущее гибридным курам (табл. 2).
От 1-суточного возраста и до завершения продуктивного периода заданная живая масса признается главным показателем роста, развития и физиологического состояния организма. Постоянное давление селекции на повышение яичной продуктивности достигается и поддерживается в основном за счет оптимизации живой массы в течение всего времени продуктивного использования при ограниченном нормированном кормлении кур. Селекционное давление в закрытых линиях позволяет постоянно удлинять продуктивный период и сохранять яйценоскость на начальную несушку на уровне 85 % и более.
|
2. Изменение основных показателей продуктивности у гибридны мере кроссов фирмы «Hy-Line International», США) |
х кур (на при- |
||
|
Показатель |
1980 год |
2002 год |
| 2009 год |
|
Возраст достижения 50 % продуктивности, сут |
161 |
145 |
143 |
|
Интенсивность яйценоскости в пик яйцекладки, % |
92 |
95 |
96 |
|
Сохранность, % |
92 |
96 |
96-97 |
|
Получено яиц на начальную несушку, шт. |
263 |
323 |
326 |
|
Живая масса в возрасте 74 нед, г |
2470 |
2000 |
1980 |
При поведении международных конкурсов (Чехия, 2009-2010 годы) куры-несушки кросса Novogen Brown французской компании «Novogen S.A.A.» снесли за 385 сут (55 нед) 351 яйцо при интенсивности яйценоскости (процентное отношение валового сбора яиц к числу кур-несушек в стаде) 91,2 % (18). Столь высокая продуктивность кур максимально приближена к ее биологической границе (табл. 3). Результаты испытаний гибридных кур-несушек ранжированы нами по яйценоскости с учетом яйце-массы (кг) как общепринятого зоотехнического показателя яичной продуктивности.
|
3. Яичная продуктивость гибридных кур (55 нед, международные конкурсные испытания, Чехия, 2009-2010 годы) |
|||||
|
Группа кроссов |
Яйценоскость на 1 несушку шт. % |
Средняя масса яйца, г |
Яйцемасса, кг |
Сохранность, % |
Конверсия корма, корм. ед/кг яйцемассы |
|
Novogen Brown («Novogen S.A.A.», Франция) 351,1 91,2 61,5 21,59 99 2,32 Hisex («Hendrix Poultry Breeders», Голландия) 346,1 90,0 61,7 21,35 96 2,32 Lognmann («Loghmann Tier-zucht GmbH», Германия) 338,9 88,0 62,5 21,18 99 2,34 isa Brown («ISA A Hendrix Genetics Company», Голландия) 342,1 88,6 61,6 21,07 98 2,37 Hy-line («Hy-Line International» (США) 336,6 87,4 61,9 20,84 98 2,33 Super Nick («H&N International GmbH», Германия) 330,1 85,7 62,3 20,56 96 2,44 Tetra («B a bolna Tetra Kft., Венгрия) 330,1 85,7 62,1 20,50 95 2,41 Bovans («ISA A Hendrix Genetics Company», Голландия) 326,8 84,5 62,7 20,49 96 2,45 |
|||||
Итак, фенотипические изменения при селекционном давлении и влиянии технологически заданных условий разведения птицы свидетельствуют о расширении физиологических границ проявления признаков продуктивности. Для высокопродуктивных кур обеспечивается возможность ежесуточного последовательного созревания в яичнике яйцеклетки-желтка до овуляции с дальнейшим завершением образования белка и скорлупы в яйцеводе и снесением полноценного яйца в суточном циркадном ритме. Яичную продуктивности кур можно прогнозировать в постэмбриональном онтогенезе по числу крупных фолликулов, созревающих в яичнике (5-6 шт.), при условии синхронизации процесса яйцеобразования и достижения 85-90 % яйценоскости. При пяти-шести крупных фолликулах в каждой серии, быстро развивающихся в яичнике, период созревания яйцеклетки-желтка должен составлять 6-7 сут. Именно фактор совпадения указанных признаков, по нашей гипотезе, определяет биологический потенциал функционирования репродуктивных органов и служит основой для раннего прогнозирования высокой яичной продуктивности птицы.
