Проблема селекции сои на устойчивость к преждевременному вскрытию (растрескиванию) бобов (сообщение I)
 бобов (сообщение I)")
Автор: Зеленцов С.В., Мошненко Е.В.
Рубрика: Селекция и семеноводство сельскохозяйственных растений
Статья в выпуске: 2 (159-160), 2014 года.
Бесплатный доступ
Преждевременное вскрытие (растрескивание) бобов является важным фактором потерь урожая сои, которые могут достигать 34-99 %. Возделывание устойчивых к растрескиванию генотипов в иных климатических условиях часто приводит к потере этой устойчивости. Это требует более детального и критического анализа используемых в селекции методов и рекомендаций по снижению растрескивания бобов. Анатомо-морфологическое изучение бобов растрескивающихся и нерастрескивающихся генотипов проводили в 2012-2014 гг. во ВНИИМК, г. Краснодар. Исследования проводили на 25 растениях каждого из семи сортообразцов сои с разной устойчивостью к растрескиванию в фазы начала и конца налива семян, и через 5 и 30 суток после созревания бобов. Под микроскопом при увеличении *40 исследовали поперечные срезы бобов. Установлено, что вскрытие боба сои всегда начинается со стороны брюшного шва и только после этого переходит на дорсальную сторону, вызывая продольный разрыв срединной жилки. Ранее описанная в литературе положительная связь между размерами вентральных склеренхимных пучков в брюшном шве и слоя соединительной паренхимы между ними, а также с формой склеренхимных пучков в срединной жилке с одной стороны, и устойчивостью к преждевременному вскрытию бобов, с другой, в климатических условиях центральной зоны Краснодарского края не подтверждается нашими исследованиями. Обнаружено широкое внутрисортовое варьирование размеров и толщины склеренхимных пучков в брюшном шве и срединной жилке бобов. На примере изучения устойчивых и неустойчивых к растрескиванию сортов сои показано, что основные, описанные в литературе анатомо-морфологические признаки устойчивости бобов сои к растрескиванию непригодны для практического использования в селекции в климатических условиях Краснодарского края. Это определяет необходимость поиска дополнительных признаков, определяющих устойчивость бобов к преждевременному вскрытию.
Бобы сои, растрескиваемость бобов, преждевременное вскрытие бобов, анатомия бобов, брюшной шов, срединная жилка, склеренхимный пучок, соединительная паренхима
Короткий адрес: https://sciup.org/142151204
IDR: 142151204 | УДК: 633.853.52:631.5/.9:581.4:631.52
The problem of soybean breeding on resistance to untimely dehiscence (shattering) of pods (report I)
The untimely dehiscence (shattering) of pods is an important factor in soybean yield losses that can reach 34-99 %. Cultivation of indehiscent genotypes in diverse environments often leads to loss of this resistance. This requires a more detailed and critical analysis used in the breeding methods and recommendations to reduce shattering of the pods. Anatomical and morphological study of the pods of dehiscent and indehiscent genotypes was carried out in 20122014 in VNIIMK, Krasnodar. Studies conducted on 25 plants of each of seven soybean accessions with the different resistance to shattering at the beginning and end of a seed ripening phase and after five and 30 days after pod maturation. Cross sections of pods investigated under a microscope at magnification of *40. It has been established that the soybean pod dehiscence always starts from the ventral suture, and only then goes to the dorsal side, causing a longitudinal gap of midrib. Previously described in the literature positive correlation between the size of the ventral sclerenchymatic bundles in the ventral suture and the conjunctive layer of parenchyma between them and the form of sclerenchymatic bundle in midrib on the one hand, and resistance to untimely dehiscence of pods - on the other hand, under the climatic conditions of the central zone of Krasnodar region is not confirmed by our research. Wide intravariety var iation of sizes and thicknesses of sclerenchymatic bundles in the ventral suture and midrib of pod is found. On the example of the study of dehiscent and indehiscent soybean varieties was shown that the basic anatomical and morphological traits of the soybean stability to shattering, described in the literature are not suitable for practical use in the breeding under climatic conditions of the Krasnodar region. This determines the necessity find additional traits in determining the untimely pods dehiscence sustainability.
Текст научной статьи Проблема селекции сои на устойчивость к преждевременному вскрытию (растрескиванию) бобов (сообщение I)
Введение . Преждевременное вскрытие или растрескивание бобов у культурной сои ( Glycine max Merrill) является одним из значимых факторов, приводящих к невосполнимой потере части урожая семян. С этой проблемой первобытные земледельцы сталкивались ещё в начале доместикации дикорастущего предка сои, сохраняет свою актуальность это негативное явление и в настоящее время [31; 32]. В ряде случаев потери семян культурной сои у неустойчивых к растрескиванию сортов могут составлять 34–99 % [35].
У всех дикорастущих видов рода Соя – Glycine Willd, включая предковый вид культурной сои – сою уссурийскую G. soja Sieb. et Zucc., а также у полукультур-ных форм сои G. max ssp. gracilis (син.: соя тонкая – G. gracilis Skw.) признак преждевременного вскрытия бобов выражен в максимальной степени [5; 8; 9; 10; 13; 16; 17; 20; 31]. Многовековой процесс доместикации, а впоследствии и селекция сои, начиная с самых примитивных методов и заканчивая современными научными методами отбора, в целом позволили заметно снизить риск преждевременного растрескивания бобов сои.
Первые известные генетические исследования по изучению признака растрес-киваемости принадлежат японскому генетику I. Nagai, описавшему в 1926 г. доминантный ген растрескиваемости Sh2 у дикорастущей уссурийской сои и рецессивный ген sh2 у устойчивого к растрескиванию японского сорта Куродайзу. Позже, в 1937 г. W.J. Morse и J.L. Karter сообщили об обнаружении у сорта культурной сои Pi-22876 гена устойчивости к растрескиванию Sh1, доминирующего над геном растрескивания sh1 [цит. по: 9; 13].
Сформированное этими исследованиями представление о качественном характере признака преждевременного вскрытия бобов преобладало в селекции сои вплоть до последней четверти ХХ века. Включение в скрещивания выявленных доноров и источников устойчивости действительно приводило к общему последовательному снижению вероятности растрескивания бобов у вновь создаваемых сортов сои, особенно по сравнению с дикорастущими и стародавними формами сои, но не снимало проблему полностью. Например, в юго-восточных штатах США даже в 80-е годы ХХ века общие оценочные потери от раннего растрескивания бобов сои всё ещё оставались высокими и достигали 37 % [35]. В настоящее время большинство современных сортов культурной сои, из-за включения устойчивости к растрескиванию в обязательный перечень учитываемых при отборе признаков, при оптимальных условиях выращивания, как правило, отличаются достаточной устойчивостью к преждевременному вскрытию бобов [2; 20].
По мере изучения явления растрески-ваемости у сои накапливались сведения о его высокой зависимости от условий окружающей среды и наличия широкой дисперсии этого признака даже в пределах одного генотипа. Например, по данным Н.И. Корсакова, в многолетних опытах ВИР при изучении признака растрескивания доля генетического фактора в его реализации составляла не более 29 % [8]. Эти и многие другие исследования привели к ревизии представлений о генетике признака преждевременного вскрытия бобов в сторону его количественного характера наследования [5; 10; 25; 26; 29; 34; 36; 40].
Многие исследователи, изучавшие этот признак, обращали внимание на тот факт, что при перестое, особенно в условиях чередования сухих и дождливых дней, риск растрескивания бобов заметно уве- личивается. А в условиях длительных засух на последних этапах органогенеза сои преждевременное вскрытие бобов нередко наблюдалось даже у высокоустойчивых к растрескиванию сортов. [5; 10; 35; 36; 40]. При этом наибольшие потери сои от растрескивания бобов перестоявших растений на фоне засух выявлены в субтропических и тропических широтах [37; 38; 40]. Наши собственные многолетние наблюдения свидетельствуют, что выращивание длиннодневных сортов сои северного экотипа в условиях укороченных фотопериодов юга России также способствует быстрому и массовому растрескиванию созревших бобов.
Имеются все основания предполагать, что заметное влияние на снижение устойчивости сортов сои к растрескиванию оказывают изменения климата. В Западном Предкавказье непрерывные климатические изменения в направлении общего потепления и сезонной аридизации фиксируются уже несколько десятилетий [6; 15]. В условиях Краснодарского края практически все сорта сои, выведенные в более высоких широтах, склонны к быстрому растрескиванию. Известны факты, когда даже местные, селекции ВНИИМК, но выведенные в погодных условиях конца 90-х годов ХХ века высокоурожайные сорта сои Лира и Дельта спустя несколько лет после начала их промышленного возделывания практически утеряли устойчивость к преждевременному вскрытию бобов. Начиная с тех же 90-х годов ХХ века, в Краснодарском крае осуществлялись неоднократные попытки интродукции устойчивых к растрескиванию канадских и французских сортов сои. Однако на фоне почти ежегодно повторяющихся позднелетних высокотемпературных засух в центральной и северной зонах края практически все эти сорта быстро теряли устойчивость к растрескиванию, что приводило к значительным потерям зерна.
Поскольку соя является строгим самоопылителем и сорт сои не является пан- миктической популяцией, то потеря сортом признака из-за существенного изменения частот генотипических классов особей с различной устойчивостью к растрескиванию исключена. Наиболее вероятным объяснением снижения устойчивости целого ряда отечественных, канадских и французских сортов сои к преждевременному вскрытию бобов может быть только негативное влияние на этот признак заметных климатических сдвигов в динамике температур воздуха и распределении осадков в периоды налива и созревания бобов относительно тех климатических условий, при которых эти сорта создавались.
Использование классических генетических методов изучения признака растрес-киваемости в селекции сои хотя и обеспечивало общее повышение устойчивости новых сортов к преждевременному вскрытию бобов, однако не давало информации о широкой экологической вариативности признака в пределах генотипа и причинах и механизмах этого явления. Это привело к необходимости углублённого изучения признака растрескивания с использованием ботанических, цитологических и молекулярно-биологических методов. Ботанический подход позволяет провести сравнительный морфологический анализ бобов в сравнении с другими видами и родами в пределах семейства [1; 11; 12]. Одной из задач цитологии является изучение строения тканей и клеток растений [21]. В молекулярной биологии широко используются методы генного картирования и анализа локусов, кодирующих количественные и качественные признаки [22]. Поэтому именно использование анатомо-морфологических, цитологических и молекулярно-биологических методов исследования внесло значительный вклад в дальнейшее изучение признака растрескивания бобов.
По данным ботанических исследований, вскрытие созревших бобов не является уникальным свойством видов рода Соя. Этот признак в той или иной степени проявляется у многих представителей семейства Бобовые – Fabaceae Lindl., в частности, практически у всех видов родов Фасоль – Phaseolus L., Вигна – Vigna Savi, Горох – Pisum L., Чина – Lathyrus L., Люпин – Lupinus L., Вика – Vicia L. и др. [10;
11; 12; 14; 23].
Среди цветковых растений вскрытие плодов является одним из пяти основных способов распространения семян, известным под названием автохория, т.е. само-распространение семян без участия посторонних агентов – ветра, воды и животных [19]. Растения-автохоры из сем. Бобовые после созревания вскрываются по створкам и механически разбрасывают семена вокруг материнского растения. В ряде случаев подпружиненные створки при резком вскрытии бобов исполняют роль баллист, увеличивая дальность разброса семян [1; 12; 19].
В литературе, посвящённой изучению признака преждевременного растрескивания сои, традиционно указывается, что створки бобов сои соединены между собой брюшным (вентральным) и спинным (дорсальным) швами, по которым и происходит вскрытие и разделение бобов на отдельные створки [17; 23; 24; 33; 34]. Однако в специализированной ботанической и карпологической литературе боб описывается как многосемянный вскрывающийся плод с сухим околоплодником в виде двух симметричных створок. При этом для обратной брюшному шву стороны бобов вместо спинного шва разными авторами используются синонимичные термины «срединная», «средняя», «медианная» или «спинная жилка» [1; 11; 12; 14]. Такие, казалось бы, незначительные терминологические различия между смысловым наполнением терминов «спинной шов» и «срединная жилка» на самом деле более принципиальны и могут оказать существенное влияние на общее понимание процесса вскрытия бобов, и как следствие, на выбор алгоритма поиска источников устойчивости к растрескиванию.
Эволюционно плод боб происходит из наиболее простого среди сосудистых растений апокарпного гинецея в виде мономерного плодолистика [1; 11; 12; 14; 18]. Ещё в XIX веке Де Кандолем (цит. по: [4]) была разработана «листовая» теория происхождения составляющих плод частей, в которой проводилась параллель между анатомическим строением гинецея (плодолистика) и листом цветковых растений. На основе этой теории карполога-ми была разработана естественная классификация плодов цветковых растений, основанная не на внешних характеристиках, а на строении гинецея и сосудистого скелета составляющих его плодолистиков как единого органа [4; 12]. В рамках «листовой» теории, принятой за базис в современной карпологии, плод бобовых представляет собой не две самостоятельные створки, соединённые между собой брюшным и спинным швами, а один плодолистик – производное от листа, в процессе эволюции свёрнутый вдоль центральной жилки с целью формирования замкнутой камеры для защиты семязачатков от внешнего воздействия. В месте соединения краёв плодолистика сформировался брюшной шов, через вентральные сосудистые пучки и плаценту обеспечивающий питанием развивающиеся семязачатки [12; 14; 18]. Осознание филогенетической связи боба с плодолистиком привело к постепенному увеличению внимания исследователей к анатомии боба в области брюшного шва и срединной жилки.
Неоднократно предпринимались попытки выявления анатомо-морфологических и цитологических различий бобов у сортообразцов сои с различной устойчивостью к растрескиванию [25; 30; 34; 37; 38; 39]. В некоторых случаях обнаруживалась корреляционная связь между растрескиванием и отдельными элементами линейных размеров бобов [25; 39], которые впоследствии не подтверждались другими исследованиями [28; 30; 36]. В целой серии исследований было установ- лено, что повышение устойчивости к растрескиванию связано с увеличением толщины створок, размеров и ширины заполненного паренхимой зазора между створками в области склеренхимных пучков брюшного шва, а также с увеличением толщины, ширины и формы склеренхимных пучков брюшного шва и срединной жилки [27; 30; 38; 39].
Вклад молекулярной биологии в решение проблемы растрескивания бобов сои пока ещё не так значителен. Группой японских исследователей в 2008 г. в тайском сорте сои SJ2 был обнаружен локус qPDH1 , кодирующий устойчивость к растрескиванию у сои, локализованный между микросателлитными маркерами SAT_366 и SAT_093 в группе сцепления J, и обеспечивающий более 50 % всей изменчивости признака [28]. Однако дальнейшие исследования не позволили выявить морфологические различия в строении бобов у носителей этого локуса, поскольку изолинии с локусом qPDH1 практически не отличались от растрескивающихся аналогов ни по линейным размерам бобов, ни по анатомическому строению брюшного шва [36].
В целом, на основе выполненных в разных странах многочисленных исследований к настоящему времени довольно подробно изучены основные причины растрескивания бобов сои. Выделены, в том числе и с использованием методов микросателлитного маркирования, источники и доноры устойчивости к этому признаку, успешно включаемые в селекционные программы. Однако возделывание устойчивых к растрескиванию сортов сои в иных эколого-географических условиях или изменение климатических условий относительно исходных, в которых эти сорта выводились и оценивались на растрескивание, зачастую приводит к возвращению проблемы растрескивания и увеличению рисков потерь урожая семян от преждевременного вскрытия бобов. Это предопределяет необходимость более детального исследования признака рас- трескивания, включая анализ причин экологической нестабильности этого признака и недостаточной эффективности отбора по ранее предложенным анатомоморфологическим признакам, препятствующим преждевременному вскрытию бобов.
Материалы и методы . Исследования анатомо-морфологического строения бобов у сои проводили на базе ГНУ ВНИИМК (г. Краснодар) в 2012–2014 гг. с использованием семи сортообразцов сои. Отечественные сорта культурной сои Лира, Фора, французский сорт Сепия и сорто-образец Д-95 дикорастущей уссурийской сои G. soya в условиях центральной зоны Краснодарского края как в предшествующие годы, так и в годы проведения исследований отличались высокой склонностью к преждевременному вскрытию бобов. Слабо растрескивающийся сорт Славия и высокоустойчивые к растрескиванию линии селекции ВНИИМК – Д-3052 TD и Д-1809 TD были использованы в качестве стандартов. Посев изучаемых сортообразцов проводили в оптимальные сроки (III декада апреля) в селекционном севообороте ВНИИМК селекционной сеялкой СКС-6А. Делянки четырёхрядные, длиной 5 м, без повторений. Норма высева семян – 30–35 шт./м2. Наблюдения за развитием преждевременного вскрытия бобов и учёт проводили на 25 растениях каждого сорта с центральных рядов через 5 суток после полного созревания по методике ВИР [3]. Дополнительно на всех исследуемых сортах по этой же методике учитывали долю вскрытых бобов через 30 суток после полного созревания растений (таблица).
Цитологические исследования поперечных срезов бобов, отобранных в фазе формирования семян и в конце фазы полного налива семян, проводили под световым микроскопом в проходящем свете при увеличении ×40. Поперечные срезы бобов выполняли в средней и наиболее широкой части бобов через семяножки и рубчики семян.
Таблица
Полевая оценка изучаемых сортообразцов сои по признаку преждевременного вскрытия бобов
ВНИИМК, Краснодар, 2012–2014 гг.
|
Показатель, год |
Сортообразец |
|||||
|
Лира |
Флора |
Сепия |
Славия |
Л-1809 ТД |
Л-3052 ТД |
|
|
Устойчивость |
Слабая |
Слабая |
Слабая |
Средняя |
Высокая |
Высокая |
|
Доля вскрывшихся бобов через 5 суток после созревания |
||||||
|
2012 |
6 |
26 |
5 |
0 |
0 |
0 |
|
2013 |
14 |
19 |
3 |
0 |
0 |
0 |
|
2014 |
1 |
35 |
16 |
0 |
0 |
0 |
|
Среднее за 3 года |
7 |
27 |
8 |
0 |
0 |
0 |
|
Доля вскрывшихся бобов че |
рез 30 суток после созревания |
|||||
|
2012 |
78 |
91 |
87 |
4 |
0 |
0 |
|
2013 |
65 |
89 |
69 |
8 |
0 |
0 |
|
2014 |
53 |
97 |
81 |
6 |
0 |
0 |
|
Среднее за 3 года |
66 |
92 |
79 |
6 |
0 |
0 |
Результаты и обсуждение . Вскрытие боба сои по брюшному шву позволяет установить его морфологическую гомологичность листу, особенно листьям с выемчатой верхушкой, например, одного из видов рода Кислица ( Oxalis L.) – кислицы Ортгиса ( O. otrgiesii Regel) (рис. 1).

а б
Рисунок 1 – Гомологичность строения вскрытого по брюшному шву незрелого боба сои ( а ) и листовой пластинки кислицы Ортгиса ( б )
Подобно обычному листу мономерный плодолистик бобовых имеет аналог центральной жилки листовой пластинки – срединную жилку, а также аналогичный мезофиллу листа мезокарпий с сетью боковых жилок.
В то же время в отличие от листа плодолистик формирует мегаспорангии или семязачатки [14; 18]. Вторым принципиальным отличием плодолистика от настоящего листа является наличие у первого брюшного шва, включающего вентральные (боковые) склеренхимные пучки с сетью сосудов и плацентарными ответвлениями (семяножками) к семязачаткам. Вентральные пучки обеих створок объединены в общий брюшной шов тонким слоем соединительной паренхимной ткани [14] (рис. 2).
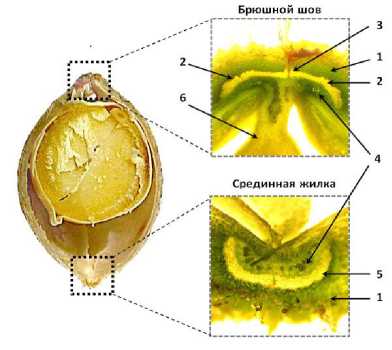
Рисунок 2 – Поперечное сечение боба сои, сорт Фора:
1 – мезокарпий створок; 2 – вентральные склеренхимные пучки; 3 – соединительная паренхима; 4 – проводящие сосуды; 5 – дорсальный склеренхимный пучок; 6 – семяножка
Отчётливая гомология между плодолистиком и листом прослеживается в строении срединной жилки боба и центральной жилки листовой пластинки (рис. 3).

а б
Рисунок 3 – Гомологичность анатомического строения поперечного сечения срединной жилки боба ( а ) и центральной жилки листовой пластинки ( б ) сои, сорт Фора
Между представленными на рисунке 3 типами жилок имеются и очевидные морфологические различия, накопившие- ся в процессе длительной эволюционной специализации. Так, трахеиды ксилемы сосудистых пучков центральной жилки листа у сои имеют гораздо больший диаметр по сравнению с сосудами срединной жилки боба. Кроме этого, сосуды центральной жилки листа равномерно распределены выше склеренхимного слоя, а у срединной жилки боба эти сосуды собраны в несколько обособленных пучков. Склеренхимная ткань центральной жилки листа представляет собой единое целое, а в склеренхимном слое срединной жилки боба в средней её части заметна поперечная тёмная полоса, состоящая из более толстостенных склеренхимных волокон уменьшенного диаметра.
Причина образования тёмного вертикального слоя клеток в срединной жилке боба не полностью ясна. Литературные данные [14] и наши цитологические наблюдения свидетельствуют об отсутствии в этой области слоя соединительной паренхимы. Возможно, это сформировавшийся в процессе эволюционного обособления плодолистика зачаток разделительного слоя, образовавшийся с целью ускорения естественного вскрытия бобов. Также образование этого слоя может быть следствием сдавливания средней части склеренхимной ткани срединной жилки за счёт встречного движении обеих створок при постепенном распирании створок развивающимися и увеличивающимися в диаметре семенами.
Ещё одним подтверждением «листовой» теории образования плодолистика может служить гомология некоторых типов аномалий бобов и листьев. Ранее нами на сое уже было изучено явление сростнолистности, вызванное дисбалансом эндогенных фитогормонов, в частности избытком цитокининов [7]. Изредка обнаруживаемые в естественных условиях или полученные искусственно полностью или частично сросшиеся листовые пластинки во всех обнаруженных случаях срастались наружными краями и никогда - центральными жилками. На некоторых сортах сои эпизодически обнаруживаются бобы, сросшиеся брюшными швами, которые представляют собой сросшиеся края плодолистиков, но никогда не встре- чаются бобы, сросшиеся срединными жилками (рис. 4).

Рисунок 4 - Г омологичность зон срастания у сросшихся брюшными швами бобов и сросшихся краями листовых пластинок сои, сорт Славия
Таким образом, морфологически боб сои имеет только один истинный шов -брюшной, соединяющий противоположные края плодолистика. Расположенная с дорсальной стороны срединная жилка по анатомическому строению гомологична центральной жилке листа и отличается повышенной, по сравнению с тканями брюшного шва, прочностью на разрыв. Поэтому инициация вскрытия бобов сои должна начинаться со стороны брюшного шва. Этот вывод подтверждают наши прямые трёхлетние наблюдения за группой сортов сои с различной склонностью к преждевременному вскрытию бобов (рис. 5). За весь период наблюдений не был обнаружен ни один боб с инициацией вскрытия со стороны срединной жилки.

а б
Рисунок 5 - Типичное вскрытие боба сои со стороны брюшного шва.
Стрелками показаны вскрывающийся брюшной шов (а) и срединная жилка (б) одного и того же боба, сорт Лира
Обычно вскрытие боба развивается со стороны средней или апикальной части брюшного шва и только затем через вер- хушку боба переходит на срединную жилку. При этом створки вскрывшихся бобов спиралевидно закручиваются наружу в обе стороны от продольной оси боба (рис. 6).

а б

в
Рисунок 6 – Основные этапы вскрытия боба сои, сорт Сепия:
а – начало вскрытия боба по брюшному шву; б – боб, вскрывшийся по брюшному шву и частично – со стороны срединной жилки;
в – полное вскрытие боба со спиральным закручиванием створок
Спиралевидное закручивание створок начинается с верхушек бобов и развивается к базальной части, завершаясь в зоне бывшей чашечки цветка, где сосудистоволокнистые пучки брюшного шва и срединной жилки переходят в общий сосудистый пучок плодоножки боба.
В ряде публикаций, посвящённых цитологическим исследованиям бобов сои, утверждалось, что одной из основных причин их растрескивания являются более тонкие в поперечном сечении склеренхимные пучки брюшного шва и слабо развитый слой соединительной паренхимы между ними. На этом основании предлагалось вести отбор на устойчивость к растрескиванию по пути увеличения размеров этих элементов боба [27; 30; 38; 39]. Однако наши данные показывают, что выраженная связь между толщиной склеренхимной ткани и соедини- тельной паренхимы со склонностью или устойчивостью к преждевременному вскрытию бобов отсутствует (рис. 7).
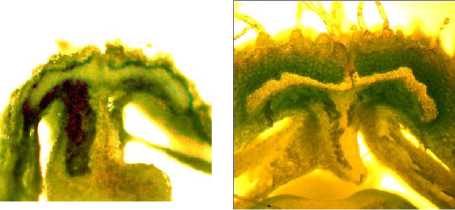
а б
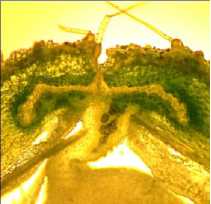
в
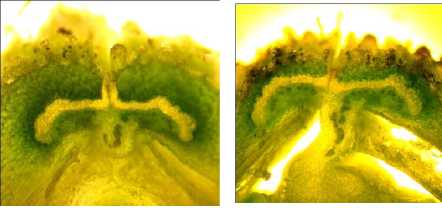
г д
Рисунок 7 – Форма поперечного сечения брюшного шва бобов у генотипов сои с различной устойчивостью к преждевременному вскрытию бобов:
а – сортообразец Д-95 дикорастущей сои ( G. soya ); б – растрескивающийся сорт Фора; в – слабо растрескивающийся сорт Славия; г – устойчивая к растрескиванию линия Д-1809 TD; д – устойчивая к растрескиванию линия Д-3052 TD
Как следует из рисунка 7, принципиальных различий по размерам поперечного сечения вентральных склеренхимных пучков между растрескивающимися и не-растрескивающимися сортообразцами сои практически нет. Более того, склеренхима брюшного шва у бобов дикорастущей уссурийской сои Д-95 (рис. 7 а ) оказалась более выражена по сравнению с генотипами культурной сои.
Относительные размеры соединительной паренхимы между вентральными склеренхимными пучками у склонного к растрескиванию сорта Фора, слабо растрескивающегося сорта Славия и устойчивой к растрескиванию линии Д-3052 TD также практически не отличаются. При этом две устойчивые к раннему вскрытию бобов линии Д-1809 TD и Д-3052 TD заметно отличаются между собой по размерам соединительной паренхимы (рис. 7г и 7д).
При этом внутрисортовое изучение линейных размеров вентральных склеренхимных пучков в брюшных швах бобов, наоборот, показало их варьирование в широких пределах (рис. 8).

Рисунок 8 – Внутрисортовая изменчивость формы и размеров вентральных пучков склеренхимной ткани и слоя соединительной паренхимы в брюшном шве бобов в фазе начала ( а ) и завершения налива семян ( б–г ), сорт Славия
В фазе начала налива семян вентральные пучки брюшного шва более мощные (рис. 8 а ), а в конце фазы они отчётливо дифференцированы, в большинстве случаев приобретают уплощённую форму. Кроме этого, у разных бобов в пределах сорта и даже одного растения наблюдаются значительные различия по толщине вентральных пучков, от очень тонких (рис. 8 б ) до имеющих в несколько раз большее поперечное сечение (рис. 8 г ). Также выявлены внутрисортовые различия по линейным размерам соединительной паренхимы между склеренхимными пучками, например на рисунках 8 в и 8 г .
Имеются работы, в частности статья G. Ghobrial и R.E. Dennis [30], где в качестве признака для отбора сои на повышенную устойчивость к растрескиванию предложено использовать сердцевидную форму срединной жилки бобов (дорсального шва). По версии авторов, такая форма поперечного сечения склеренхимного пучка в срединной жилке повышает устойчивость бобов сои к преждевременному вскрытию, а развёрнутая и уплощённая форма пучка, наоборот, свидетельствует о склонности генотипа к растрескиванию.
Однако в наших исследованиях наиболее близкую к сердцевидной форму имели склеренхимные пучки срединных жилок бобов дикорастущего сортообраз-ца Д-95 уссурийской сои, отличающегося склонностью к самому раннему растрескиванию (рис. 9 а ).
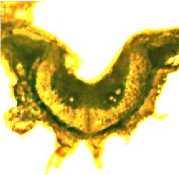
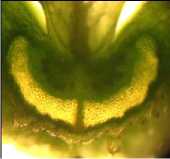
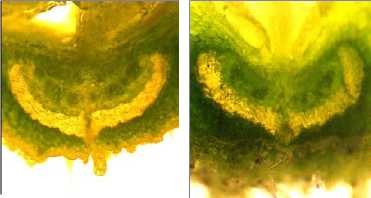
а б
в
г
Рисунок 9 – Форма поперечного сечения срединной жилки бобов у генотипов сои с различной устойчивостью к преждевременному вскрытию бобов:
а – сортообразец Д-95 дикорастущей сои ( G. soya ); б – растрескивающийся сорт Фора; в – слабо растрескивающийся сорт Славия; г – устойчивая к растрескиванию линия
Д-1809 TD
Формой дорсального склеренхимного пучка, близкой к сердцевидной, также обладали бобы стремительно растрескивающегося после созревания сорта Фора (рис. 9б). При этом конфигурация поперечного сечения склеренхимных пучков срединных жилок у слабо растрескивающегося сорта Славия и устойчивой к рас- трескиванию линии Д-1809 TD была более раскрытой и уплощённой (рис. 9в и 9г).
Аналогично широкой внутрисортовой вариативности размеров вентральных пучков брюшного шва (см. рис. 8) практически у всех изучаемых сортообразцов обнаружено внутрисортовое варьирование формы поперечного сечения склеренхимного пучка срединных жилок (рис. 10).
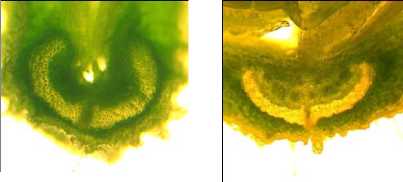
а б
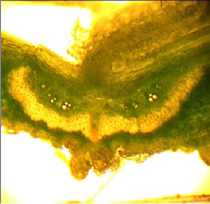
в
Рисунок 10 – Внутрисортовая изменчивость формы дорсального склеренхимного пучка в поперечном сечении срединной жилки у бобов в фазе начала ( а ) и завершения налива семян ( б – в ), сорт Славия
Отчётливо выраженная сердцевидная форма этого пучка (рис. 10 а ) в начале фазы налива бобов по мере развития и увеличения крупности семян приобретает более уплощённую форму. Причём у разных бобов в пределах сорта в конце фазы налива семян также обнаруживается изменчивость конфигурации поперечного сечения срединной жилки (рис. 10 б и 10 в ).
Выводы. На основе анализа карпологической литературы и собственных анатомо-морфологических исследований показана необоснованность применения термина «спинной шов» в отношении срединной жилки, расположенной на обратной брюшному шву продольной стороне боба.
Визуальные наблюдения показали, что инициация вскрытия боба сои всегда начинается со стороны брюшного шва и только после этого переходит на обратную сторону, вызывая продольный разрыв срединной жилки.
Описанная в литературе положительная связь между толщиной вентральных склеренхимных пучков в брюшном шве и слоя соединительной паренхимы между ними, а также с формой дорсальных склеренхимных пучков в срединной жилке, с одной стороны, и повышенной устойчивостью к преждевременному вскрытию бобов, с другой, в климатических условиях центральной зоны Краснодарского края не подтверждается.
Обнаружено широкое внутрисортовое варьирование линейных размеров и формы вентральных и дорсальных склеренхимных пучков, затрудняющее выявление межгенотипических различий по этим показателям.
В целом, проведённая оценка основных известных в литературе анатомических признаков боба, по мнению их авторов, успешно позволяющих выделять формы сои с повышенной устойчивостью к преждевременному вскрытию бобов, показала их непригодность для практического использования в селекции на устойчивость сои к растрескиванию в климатических условиях Краснодарского края. Это, в свою очередь, определяет необходимость поиска дополнительных признаков, определяющих устойчивость бобов к преждевременному вскрытию.
