Продуцирование и энергетические показатели биогеоценозов полупустынь Ганых-Айричайской долины Азербайджана

Автор: Сарыева Гюнель Рахиб
Журнал: Бюллетень науки и практики @bulletennauki
Рубрика: Биологические науки
Статья в выпуске: 9 т.6, 2020 года.
Бесплатный доступ
Представлены результаты фитопродуктивности в динамике и некоторые энергетические показатели Artemisetum-Ephemerosum и Alhagietum-Artemisosum на территории Ганых-Айричайской долины Азербайджана. Это типичные полупустынные ценозы. Анализ количественных показателей энергетических расходов в растительных сообществах за период исследований показал, что 2 вида погодно-климатических условий определили 2 контрастных периода развития. Даны сведения о надземной и подземной биомассе в исследуемых сообществах. Проведен сравнительный анализ.
Фитопродуктивность, прирост, запасы фитомассы, биоценоз, энергетический баланс
Короткий адрес: https://sciup.org/14117912
IDR: 14117912 | УДК: 631.46 | DOI: 10.33619/2414-2948/58/08
Production and some energy indicators in semi-desert biogeocenosis of the Ganykh-Airichai valley of Azerbaijan
The results of phytoproductivity in dynamics and some energy indicators of Artemisetum-Ephemerosum and Alhagietum-Artemisosum on the territory of the Ganykh-Airichai valley of Azerbaijan are presented. These are typical semi-desert coenosis. Analysis of quantitative indicators of energy consumption in plant communities over the period of research showed that 2 types of weather and climatic conditions determined 2 contrasting periods of development. Data on the aboveground and underground biomass in the studied communities are given. A comparative analysis has been carried out.
Текст научной статьи Продуцирование и энергетические показатели биогеоценозов полупустынь Ганых-Айричайской долины Азербайджана
Бюллетень науки и практики / Bulletin of Science and Practice
УДК 631.46
Исследования ресурсов биосферы на современном этапе, особенно с глобальным изменением климата, приобретает особую актуальность, в частности определение действительной и потенциально возможной первичной биологической продуктивности фитоценозов в целях сохранения и восстановления высокопродуктивных растительных сообществ.
Выявление основных закономерностей качественного и количественного распределения и воспроизводства органического вещества в интересах наиболее рационального использования продукции Земли человеком, является одной из основных проблем современной биологической науки, решению которого посвящено достаточно много работ [1–3].
Запасы фитомассы суши, по данным Н. И. Базилевич [2], составляют 2,4×1012 т (в океане — 2×108 т); запасы подстилки и торфа на порядок ниже общего количества фитомассы на планете и соответственно составляют 1,9×1011 и 2,2×1011 т, годичная продуктивность фитомассы Земли ровна 2,3×1011 т, на 74% — определяется надземными растительными формациями.
При этом биологическая продуктивность фито- и агрофитоценозов в основном изучена без учета энергетических ресурсов. Основные усилия исследователей уделялись изучению круговорота веществ в биогеоценозе. Энергетическая же сторона природных превращений оставалась мало изученной, хотя на важность этого подхода в свое время указал еще А. Г. Дояренко [4].
Объект и методика исследований
Объектом исследования являются естественные растительные сообщества эфемеро– полынная ( Artemisetum–Ephemerosum ) и полынно–верблюжьеколючковая ( Alhagietum– Artemisosum ) ассоциации, которые распространены в Ганых-Айричайской долине, расположенной на Южном склоне Большого Кавказа, входящей и занимающей наибольшую площадь в основном в Шекинском районе.
Описывая природные условия следует отметить, что данный регион достаточно сложный, где субальпийский и альпийские луга по снижению гипсометрического уровня и изменению климатических условий, постепенно заменяются лесами и далее горными степями равнинами.
В геологическом и геоморфологическом отношении регион имеет достаточно сложное строение, где высокие горы и отвесные склоне чередуются предгорными территориями и равнинами, что в свою очередь способствует развитию довольно пестрого климата то разнообразию растительного покрова.
Сложность геоморфологического строения, наличие крутых склонов, а также антропогенное воздействие, оказывают существенное влияние на интенсивность поверхностного стока, что в свою очередь усиливает развитие эрозионных процессов. Создаются широкие овраги, которые в конечной стадии представлены балками [5].
Склоны гор сильно расчленены и доминирует эрозионно-денудационный рельеф. Почвообразующие породы представлены сланцами, известняками и песками, встречаются базальты, габбро, гидрослюды, андезиты и др.
Низкогорье расположено между изогипсами 600–1000 м над у. м. Поверхность представлена узкими поясными островками, отделенные между собою широкими долинами. Почвы сформированы на отложениях третичного и четветичного периодов Кайнозоя.
Равнинная зона расположена на высоте 200–600 м над уровнем моря, где в основном представлены аллювиально-проалювиальные отложения, на которых развиты плодородные почвы, интенсивно использующиеся в сельском хозяйстве.
В Шеки-Закатальской зоне Э. М. Шихлинский [6] выделяет 3 типа климата:
-
1) умеренно-теплый, характерной для равнинных территорий зоны с относительно мягкой зимой;
-
2) умеренно-теплый влажный тип с равномерным распределением осадков в течение всего года, охватывает предгорную часть зоны;
-
3) холодный климат с влажной зимой, характерной для предгорной и горной области зоны.
Минимальная температура воздуха 0,5 °С приходится на январь, а максимальная в 23,6 °С на июль. Среднемесячная температура почвы колеблется в пределах -1°С … -30,7 °С. Минимальная температура почв в -1 °С приходится на январь, максимальная 30,0 °С — июль и август 30,7 °С. Годовое количество осадков изменяется с увеличением гипсометрического уровня и соответствует на равнине — 939 мм, а в высокогорьях — 1400 мм. Среднегодовая относительная влажность воздуха — 71%, колеблясь по временам года от 59% до 87%.
Первые геоботанические исследования в целом на Южном склоне Большого Кавказа были проведены Н. И. Кузнецовым [7], позже — Л. И. Гроссгеймом [8], В. Д. Гаджиевым [9], Л. И. Прилипко [10] и др.
Анализ и результаты
Ключевые участки выбранные нами имеют географические координаты N41°,08ʹ43.90ʺ E47°,02ʹ09.60ʺ (238 м н. у. м.); N41°,08ʹ41.55ʺ E47°,02ʹ08.75ʺ (242 м н. у. м.); N41°,06ʹ22.70ʺ E47°,04ʹ53.59ʺ (272 м н. у. м.);
Учет массы надземных частей исследуемых фитоценозов проводился путем срезания растений у поверхности растений у поверхности почвы на учетной площадке размером 1 м2 в трехкратной повторности. При определении подземной фитомассы растительных сообществ брались монолиты с площадки 25 см2 в трехкратной повторности до глубины 50 см, через каждые 10 см. Для отмывки подземных частей растений от почвы пользовались двумя ситами, из которых верхнее имело ячейки 3 мм, а нижнее — 0,25 мм. Корни взвешивались после отмывки в воздушно-сухом состоянии, а растительные остатки относились к надземной массе. Учет массы надземных и подземных частей фитоценозов проводились в апреле, мае, июне и октябре 2015–2016 гг.
Прирост растительной фитомассы и энергия, аккумулированная в растительном веществе принимались по В. Р. Волобуеву [11].
Состав ассоциации Artemisetum–Ephemerosum представлен следующими видами: Artemisia lerchiana Weber., Parapholis incurva (L.) C. E. Hubb., Alyssum desertorum Stapf., Sclerochloa dura (L) Beauv., Medicago minima (L.) Bartalini, Medicago orbicularis (L.) Bartalini, Medicago monspeliaca L., Avena fatua L., Lolium rigidum Gaudin., Poa bulbosa L., Bromus scoparius L., Bromus japonicus Thunb., Salvia viridis L., Hirschfeldia incana (L.) Lagr.-Foss., Hordeum geniculatum All., Asrtagalus hamosus L., Althaea hirsuta L., Papaver hybridium L.
Весенний период характеризуется обилием видов эфемеро–полынных растений, продуктивность которых зависит главным образом от количества выпавших осадков в весенний и осенний периоды и от динамики нарастания температуры воздуха. От весны к осени масса полыни увеличивается, а общая масса эфемеров уменьшается. Эти данные согласуются с данными Ю. А. Зейналова [12] и А. М. Керимова [13].
Исследования проводились в 2015–16 гг., резко различающихся по наличию осадков. Так, если в 2015 г. — атмосферных осадков было 543,7 мм, то в 2016 г. — 960,3 мм.
В засушливых условиях 2015 г. при минимальных осадках запасы фитомассы эфемеро– полынной ассоциации составили 11090,0 г/м2, а в 2016 г., при обильных осадках — 13061,0 г/м2.
Запасы полынно–верблюжьеколючковой ассоциации также изменились, составляя соответственно по годам: 11752,0 и 12375,0 (г/м2).
Как следует из Таблицы 1 — максимальные значения надземной фитомассы эфемеро– полынной ассоциации приходятся на май (292 г/м2 и 225 г/м2), в среднем — 259 г/м2, а минимальные — на октябрь (107 г/м2 и 72 г/м2).
В апреле значения надземной фитомассы эфемеро–полынной ассоциации составили 145 г/м2 и 103 г/м2 (в среднем 124 г/м2). Динамика запасов надземной фитомассы верблюжьей колючки и эфемеров показала несколько иную картину. Максимальные значения — в июне, составляя при этом по годам исследования 231,2 г/м2 и 337,2 г/м2, в среднем — 282,2 г/м2. Минимально — в октябре, составляя 127,6 г/м2 и 181,4 г/м2, в среднем — 154,5 г/м2 (Таблица 2).
Таблица 1.
|
Месяцы |
Надз. масса |
Растительные остатки |
Общая надземная масса |
Глубина, см |
Подземная масса |
% |
Общая растительная масса |
|
2015 |
|||||||
|
Апрель |
125,8 |
19,3 |
145,1 |
0–10 |
725,1 |
64,88 |
1262,7 |
|
10–20 |
202,3 |
18,10 |
|||||
|
20–30 |
124,4 |
11,13 |
|||||
|
30–40 |
36,2 |
3,23 |
|||||
|
40–50 |
29,5 |
2,62 |
|||||
|
Всего:117,6 |
100 |
||||||
|
Maй |
250,1 |
42,1 |
292,2 |
0–10 |
1013,5 |
67,60 |
1791,4 |
|
10–20 |
241,3 |
16,09 |
|||||
|
20–30 |
162,1 |
10,83 |
|||||
|
30–40 |
44,1 |
2,94 |
|||||
|
40–50 |
38,2 |
2,54 |
|||||
|
Всего: 1499,2 |
100 |
||||||
|
Июнь |
207,1 |
24,1 |
231,2 |
0–10 |
851,2 |
67,49 |
1492,4 |
|
10–20 |
230,1 |
18,24 |
|||||
|
20–30 |
101,5 |
8,06 |
|||||
|
30–40 |
41,3 |
3,27 |
|||||
|
40–50 |
37,1 |
2,94 |
|||||
|
Всего: 1261,2 |
100 |
||||||
|
Октябрь |
98,4 |
8,5 |
106,9 |
0–10 |
491,3 |
63,80 |
875,7 |
|
10–20 |
139,1 |
18,09 |
|||||
|
20–30 |
92,1 |
11,90 |
|||||
|
30–40 |
27,1 |
3,52 |
|||||
|
40–50 |
19,2 |
2,69 |
|||||
|
Всего: 768,6 |
100 |
||||||
|
2016 |
|||||||
|
Aпрель |
92,4 |
10,3 |
102,7 |
0–10 |
519,1 |
66,31 |
885,5 |
|
10–20 |
133,3 |
17,03 |
|||||
|
20–30 |
93,2 |
11,91 |
|||||
|
30–40 |
20,5 |
2,62 |
|||||
|
40–50 |
16,7 |
2,13 |
|||||
|
Всего: 782,8 |
100 |
||||||
|
Maй |
194,9 |
30,3 |
225,2 |
0–10 |
915,3 |
72,13 |
1494,1 |
|
10–20 |
179,1 |
14,11 |
|||||
|
20–30 |
118,7 |
9,35 |
|||||
|
30–40 |
31,9 |
2,53 |
|||||
|
40–50 |
23,9 |
1,88 |
|||||
|
Всего: 1268,9 |
100 |
||||||
|
Июнь |
137,1 |
19,1 |
156,2 |
0–10 |
675,8 |
71,32 |
1103,7 |
|
10–20 |
151,7 |
16,01 |
|||||
|
20–30 |
73,2 |
7,73 |
|||||
|
30–40 |
27,7 |
2,92 |
|||||
|
40–50 |
19,1 |
2,02 |
|||||
|
Всего: 947,5 |
100 |
||||||
|
Октябрь |
64,7 |
6,9 |
71,6 |
0–10 |
399,5 |
66,13 |
675,7 |
|
10–20 |
110,1 |
16,23 |
|||||
|
20–30 |
69,5 |
11,50 |
|||||
|
30–40 |
14,1 |
2,34 |
|||||
|
40–50 |
10,9 |
1,80 |
|||||
|
Всего: 604,1 |
100 |
||||||
Таблица 2.
|
Месяцы |
Надз. масса |
Растительные остатки |
Общая надземная масса |
Глубина, см 2015 |
Подземная масса |
% |
Общая растительная масса |
|
Aпрель |
123,8 |
12,7 |
136,5 |
0–10 |
125,2 |
33,47 |
510,5 |
|
10–20 |
102,3 |
29,48 |
|||||
|
20–30 |
88,5 |
20,50 |
|||||
|
30–40 |
30,5 |
8,62 |
|||||
|
40–50 |
27,5 |
7,93 |
|||||
|
Всего: 374,0 |
100 |
||||||
|
Maй |
250,1 |
42,1 |
292,2 |
0–10 |
1013,5 |
67,60 |
1791,4 |
|
10–20 |
241,3 |
16,09 |
|||||
|
20–30 |
162,1 |
10,83 |
|||||
|
30–40 |
44,1 |
2,94 |
|||||
|
40–50 |
38,2 |
2,54 |
|||||
|
Всего: 1499,2 |
100 |
||||||
|
Июнь |
207,1 |
24,1 |
231,2 |
0–10 |
851,2 |
67,49 |
1492,4 |
|
10–20 |
230,1 |
18,24 |
|||||
|
20–30 |
101,5 |
8,06 |
|||||
|
30–40 |
41,3 |
3,27 |
|||||
|
40–50 |
37,1 |
2,94 |
|||||
|
Всего: 1261,2 |
100 |
||||||
|
Октябрь |
116,4 |
11,2 |
127,6 |
0–10 |
142,3 |
37,45 |
507,6 |
|
10–20 |
133,2 |
35,05 |
|||||
|
20–30 |
82,4 |
21,68 |
|||||
|
30–40 |
22,1 |
5,82 |
|||||
|
40–50 |
— |
— |
|||||
|
Всего: 380,0 |
100 |
||||||
|
2016 г. |
|||||||
|
Aпрель |
162,4 |
22,3 |
184,7 |
0–10 |
219,1 |
41,57 |
711,8 |
|
10–20 |
173,3 |
32,88 |
|||||
|
20–30 |
96,4 |
18,29 |
|||||
|
30–40 |
22,7 |
4,30 |
|||||
|
40–50 |
15,6 |
2,96 |
|||||
|
Всего: 527,1 |
100 |
||||||
|
Maй |
210,7 |
26,3 |
237,0 |
0–10 |
547,2 |
57,96 |
1181,1 |
|
10–20 |
229,4 |
24,30 |
|||||
|
20–30 |
105,6 |
11,18 |
|||||
|
30–40 |
41,4 |
4,39 |
|||||
|
40–50 |
20,5 |
2,17 |
|||||
|
Всего: 944,1 |
100 |
||||||
|
Июнь |
337,1 |
29,1 |
336,2 |
0–10 |
675,8 |
71,32 |
1513,7 |
|
10–20 |
351,7 |
16,01 |
|||||
|
20–30 |
73,2 |
7,73 |
|||||
|
30–40 |
27,7 |
2,92 |
|||||
|
40–50 |
19,1 |
2,02 |
|||||
|
Всего: 1147,5 |
100 |
||||||
|
Октябрь |
164,2 |
17,2 |
181,4 |
0–10 |
464,1 |
58,92 |
969,1 |
|
10–20 |
197,4 |
25,06 |
|||||
|
20–30 |
82,5 |
10,47 |
|||||
|
30–40 |
26,5 |
3,37 |
|||||
|
40–50 |
17,2 |
2,18 |
|||||
|
Всего: 787,7 |
100 |
ДИНАМИКА НАДЗЕМНОЙ И ПОДЗЕМНОЙ ФИТОМАССЫ Alhagietum–Artemisosum (г/м 2 , 2015–2016 гг.)
Подземная фитомасса травяных фитоценозов аридных зон, как отмечается в литературе превышает надземную [1–3, 12–13]. Подземная фитомасса полынно–эфемерового сообщества в апреле соответственно по годам составила 1118 г/м2 и 783 г/м2 (в среднем — 951,0 г/м2).
В мае запасы подземной фитомассы достигли максимальных величин 1499 г/м2 и 1269 г/м2 (в среднем — 1384,0 г/м2), а минимальной в октябре 769 г/м2 и 604 г/м2 (в среднем — 687,0 г/м2). На верхний горизонт (0–20 см), где расположены преимущественно корневые системы сообщества, приходится до 80–85% всей подземной массы от его общего объема, что указывает на его экологическую приспособленной к окружающей среде. В слое 40–50 см почвы находится от 2,6% до 3,2 % всех корней.
К летнему периоду вся зеленая масса уменьшается. В июне надземная фитомасса согласно по годам 2015-16 гг. составила 231 и 156 (г/м2) (в среднем — 194,0 г/м2), а подземная фитомасса в слое 0–50 см 1261 г/м2 и 948 (г/м2) (в среднем — 1105,0 г/м2). При этом общая фитомасса составила 1492 г/м2 и 1104 (г/м2) (в среднем — 1298,0 г/м2). Это объясняется высокими значениями температуры воздуха и малым количеством атмосферных осадков (+26 °С и 7,7 мм).
Анализируя динамику надземной фитомассы верблюжьей колючки и полыни, следует отметить, что максимальных величин они достигаю в июне, составляя 231,2 г/м2 и 337,2 г/м2 (в среднем — 284,2 г/м2), а подземная фитомасса при этом составила в слое 0–50 см 1261,2 г/м2 и 1147,5 г/м2 (в среднем — 1204,4 г/м2). При этом запасы общей фитомассы в июне достигли до 1492,4 г/м2 и 1513,7 г/м2 (в среднем — 1503,1 г/м2).
Минимальные значения подземной фитомассы данной ассоциации приходится на апрель, составляя по годам 374,0 г/м2 и 527,1 г/м2 (в среднем — 450,6 г/м2) в слое 0–50 см почвы. Основная масса корней сосредоточена в слое почвы 0–20 см, составляя 60–65%.
Общая фитомасса полыни и верблюжьей колючки в июне составили 1492,4 г/м2 и 1513,7 г/м2 (в среднем — 1503,1 г/м2).
-
Н. И. Базилевич и Л. Е. Родин [14] отмечают, что связи биологической продукции с условиями увлажнения носят различный характер в зависимости от количества получаемого тепла. При высокой теплообеспеченности (более 35–40 ккал/см2 год) важнейшим фактором, регулирующим биологическую продуктивность, является влага: при низкой теплообеспеченности (до 35–40 ккал/см2 год) избыток влаги снижает производительность фитоценозов.
Как отмечает В. Р. Волобуев [11] прирост и разложение многолетних растений — очень сложный вопрос и до настоящего времени остается недостаточно изученной. В исследованиях была проведена оценка величины прироста и разложения растительного вещества по методике, предложенной В. Р. Волобуевым [11] (дифференциальный анализ кривой динамики растительных веществ) в биогеоценозе и разложения растительного вещества. Данный метод отличается тем, что при его помощи можно найти прирост и разложение для каждого месяца года, используя экспериментальные данные о динамике фитомассы.
На Рисунках 1–2 показана динамика накопления и разложения общей растительной массы эфемеро–полынной (Artemisetum–Ephemerosum) и полынно–верблюжьеколючковой (Alhagietum–Artemisosum) ассоциаций, которая отражает сложное соотношение во времени двух противостоящих процессов: накопления и разложения растительной массы. Из Рисунков 1–2 следует, что в эфемеро–полынном сообществе в 2016 г. запасы надземной и подземной фитомассы больше, чем в 2015 г. (130,6 т/га против 110,9 т/га), что связано с более благоприятными климатическими условиями в 2016 г., когда атмосферных осадков выпало выше нормы — 960,3 мм.
Кривая Рисунка 1 указывает на то, что до мая темп накопления растительной массы эфемерово-полынного сообщества превышает показатели ее разложения, а в последующие времена года соотношение обратное. Прибегнув к методу дифференцированного анализа кривой динамики растительной массы эфемеро–полынной ассоциации, можно дать количественную оценку этих процессов в отдельности.
По динамике растительной массы за периоды преобладания накопления растительного вещества над его разложением (т. е. по восходящим отрезкам кривой) определим ежемесячные приращения растительного вещества эфемерово-полынного сообщества, а за периоды количественного преобладания разложения растительных остатков (т. е. по нисходящим отрезкам кривой) их ежемесячную убыль. Снятые данные хорошо выявляют общий закономерный ход каждого из этих процессов, т. е. как накопления, так и разложения растительных веществ. Этот закономерный вид данных позволяет нам путем экстраполяции продолжить кривые и для периодов, на которые нет прямых данных, и таким путем раздельно выявить вероятный порядок изменения накоплении я и разложения растительных веществ. В частности, пользуясь кривой можно приближенно определить как величину накопления вещества за период количественного преобладания процессов их разложения, так и размеры разложения за период количественного преобладания процессов накопления растительных веществ.
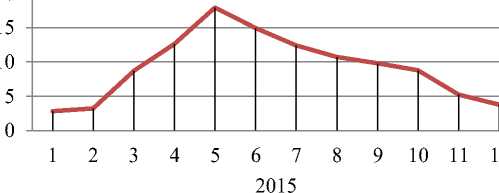
Рисунок 1. Динамика накопления и разложения растительной массы Artemisetum–Ephemerosum сообщества (надземной и подземной, т/га, 2015–2016 гг.) .
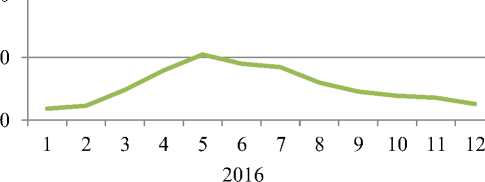
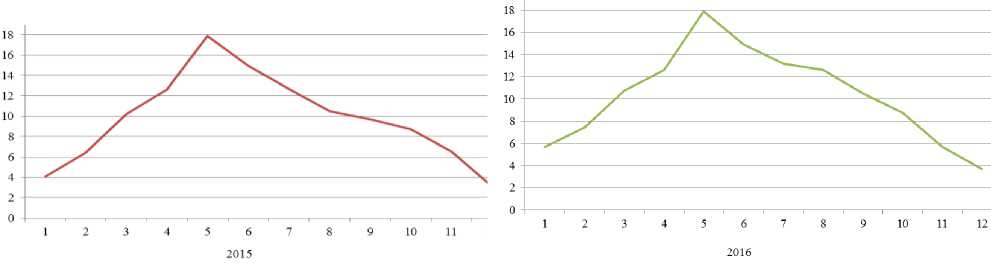
Рисунок 2. Динамика накопления и разложения растительной массы Alhagietum–Artemisosum (надземной и подземной, т/га, 2015–2016 гг.)
Действительный прирост растительного вещества эфемеро–полынного сообщества за период 2015–2016 гг. в данных условиях составил 14,2 т/га и 12,83 т/га.
Для выявления количества энергии, аккумулированной в приросте растительного вещества, необходимо иметь данные об энергии сгорания растительной массы. Для этой цели использовались калориметрическими данными А. П. Герайзаде [11, с. 56]. Как утверждает автор, за год в среднем для эфемеро–полынного сообщества — 5015 кал., а для верблюжьей колючки — 5420 кал. на г сухого вещества.
Данные расчетов энергетических затрат этого биогеоценоза прприведены в Рисунке 1. Исходя из этих данных и темпов прироста накопления и расхода энергии во времени в эфемеро–полынном сообществе наибольшее накопление энергии в фитомассе в 2015 г. приходится на март — 115,3 кДж, а 2016 г. на июнь — 127,7 кДж. Зимой же эти значения минимальны (8,6 кДж и 19,5 кДж). За год в эфемеро–полынном сообществе накопление энергии составило 337,0 кДж и 269,4 кДж (Рисунок 1).
В полынно–верблюжьеколючковой ассоциации прирост растительной массы за год составил 30,48 т/га и 11,64 т/га. Максимальные значения прироста растительной массы приходятся на июнь, составляя 8,11 т/га и 8,33 т/га, а минимальные на январь — 1,27 т/га и 1,54 т/га. Показатели разложения растительного вещества за год составили 9,42 т/га и 11,64 т/га.
Максимальные значения их в 2015 г. приходятся на июль, составляя 2,52 т/га, а минимальные 0,66 т/га — на декабрь. В 2016 г. максимальные показатели разложения растительной массы наблюдаются в сентябре — 2,67 т/га, а минимальные в декабре — 1,22 т/га.
На основе калориметрических данных выявлено, что накопление энергии в полынно– верблюжьеколючковой ассоциации за год составили 641,2 кДж и 691,3 кДж, а энергия разложения растительного вещества 213,6 кДж и 264,0 кДж. Максимальные значения накопления энергии исходя из данных прироста растительной массы приходятся на июнь, составляя 183,9 кДж и 188,9 кДж, а минимальные — на январь — 28,8 кДж и 34,9 кДж.
Показатели энергии разложения растительного вещества за год составили 213,6 кДж и 264,0 кДж. При этом максимальные значения энергии разложения в 2015 г приходятся на июнь, составляя 57,2 кДж. А в 2016 г. — на сентябрь — 60,6 кДж. Минимальные показатели энергии разложения приходятся на декабрь, по годам — 14,9 кДж и 27,7 кДж (Таблица 3).
Определенный интерес представляют расчеты энергетических затрат. Так, исходя из средних значений величины затрат энергии на скрытую теплоту парообразования в 580 кал/см3, определяем общий расход энергии на испарение в биогеоценозе. Таковыми по годам данные показатели составили 147,7 кДж и 294,0 кДж. Максимальные их значения приходятся на июнь, составляя 33,56 кДж и 55,31 кДж, а минимальные несколько отличны, показатели которых в 2015 г. приходятся на декабрь, составляя 2,65 кДж, и в 2016 г. — 4,51 кДж соответствуя январю (Таблица 3).
Установив величину суммарного расхода энергии на суммарное испарение + энергия аккумулированная в чистой первичной продукции, находим общую величину затрат энергии в биогеоценозе. В эфемеро–полынном сообществе расход энергии на почвообразование в 2015 г. составило 485,1 кДж, расход энергии на биологические процессы в почве 337,0 кДж, а относительная доля расхода энергии на биологические процессы 0,8% от общей суммы затрат энергии на почвообразование. Для 2016 г. данные показатели составили: затрат энергии на почвообразование — 563,4 кДж, биологические процессы — 269,4 кДж, и относительная доля формирования растительного вещества от общих затрат энергии на почвообразование 1,19% (Таблица 3).
ДИНАМИКА НАДЗЕМНОЙ И ПОДЗЕМНОЙ ФИТОМАССЫ Artemisetum-Ephemerosum (г/м 2 , 2015–2016 гг.)
Бюллетень науки и практики / Bulletin of Science and Practice Т. 6. №9. 2020
Таблица 3.
ЭНЕРГЕТИЧЕСКИЙ БАЛАНС Artemisetum–Ephemerosum НА ОСНОВЕ ДАННЫХ ПО ДИНАМИКЕ РАСТИТЕЛЬНОЙ МАССЫ И СУММАРНОГО ИСПАРЕНИЯ ЗА 2015–16 гг.
|
I |
II |
III |
IV |
V |
VI |
VII |
VIII |
IX |
X |
XI |
XII |
Год |
|
|
2015 |
|||||||||||||
|
S я л н ч □ а о о § Й ,я н Д й О |
ОО ci |
со сч |
ОО |
40 ri W |
О\ |
со |
?—( ci w |
с о" W |
ОО oi |
40 ОО |
со сч tn |
40 |
о" ?—< |
|
н й о ’о § 2 Й & К 5 Ч а о ч в о С л о о К S । и «^+ д |
tn 04^ о |
о |
04 tn |
со 04^ |
ОО сч tn |
in о 40 W |
|||||||
|
к i 3 =s к 1 « йВооий2^ инйВч*й йв2а«иЧя: в й 5S К g в й ЙйООКй.О |
04 04^ ci |
tn ci |
04 40^ W |
04^ о |
in о W |
tn |
40 W |
in W |
|||||
|
Ы Ом I I 2 й | 9< ? в в ° u й Ц св н Я И Д И о св U И св а & « |
^1 о? tn |
со 40^ |
°\ ci ОО W |
сч tn 40 СО |
40 |
40 О 40 СО |
W tn СО со |
о 40 О со |
04^ W |
40^ oi о W |
°\ ОО |
°\ ОО СО СО |
|
|
3 о и 2 ё « о Я Я 2 о ^ 2 ° ° И g я 8 я 3» |
04^ о? |
ю. ОО |
tn |
ОО |
ОО о |
о |
|||||||
|
а Й Ч « « о йз § g а 8 * |
ci 40 |
ci in |
tn tn |
oi W |
ci со |
я |
04 со |
||||||
|
к 8 2 о |
tn о in |
40^ ОО |
СО 40 |
О? СО |
С |
сс |
W |
о |
о? W |
°1 |
tn |
||
|
я щ ° я ^ 2 о а о г |
tn ci W |
ОО со |
tn ci 40 |
ОО 40 |
СЧ tn |
ОО W |
40^ ОО |
О\ tn |
ci |
со |
°\ |
о W |
о 40 |
|
Зв & £ в ^ а 5 я § С о К ^ |
in о |
со чо |
in сч tn |
СО 40 |
?—< ОО W |
40 »П |
ОО tn о со |
W W |
со о" W |
чо tn |
со ^ |
in ^ ci |
?—< |
|
й ' h 5^“чо |
°\ ci со |
tn |
40 о ?—< |
О oi 04 |
04 СО w |
40 »П |
ОО tn о со |
W W |
со о" W |
40 tn |
со |
in ^ ci |
W tn |
|
hQ 1 CQ 1 6х 5 § s § >а я ~ § 4 & S В ° Й О^^4йНЯц я S « Й « 2 я Й m И 2 Я О я & & О |
ОО о |
40 tn о |
ОО ОО о |
о |
40 ОО О |
1 |
1 |
1 |
1 |
1 |
1 |
1 |
О |
|
I |
II |
III |
IV |
V |
VI |
VII |
VIII IX |
X |
XI XII |
Год |
|||
|
2016 |
|||||||||||||
|
К X И S < g ft ч d Н |
СО 40 |
МП МП |
04 40^ of |
40 ОО МП W |
o' 04 |
о ОС W |
04 ОО 40 w |
О' of W |
of |
40 |
04 W |
W МП |
ОО |
|
>x + - н 2 о х О ® К И ° О о2моХХщй &ox«222ft KHftXceftgv- С н S и к 5 ч |
МП W |
04 о |
о |
W |
ОО О' 40 |
со of W |
|||||||
|
X 2 О X >Х S' и 3 к >s й 2 ° ЧйнОКХи- х о 2 й S о ^ g, 5-й |
С4 |
С4 W |
04 W W |
04 ОО o' |
гп o' |
о of |
О' of |
of W |
|||||
|
И S 5 и й и о а ч (ТЧ и о св и Св о^ & « |
<0 чо |
ю. МП 04 |
О' 04 О 04 |
W |
of |
О' ОС |
О' of МП 04 |
04 w |
О' 40 w |
МП of |
о W |
Ю. |
|
|
к л (Ud И К ч «2 5 g н S g | 2 2 2 ° й |
МП 04 ?—< |
МП |
О' 40 04 |
04 W |
of 40 04 |
||||||||
|
в - О - j ft ч ся н 2 м § к s « Н Ч С & х н “и |
04" ОО |
о |
МП 04 |
ОС W |
■вг |
МП МП |
°1 of |
of 40 04 |
|||||
|
к 8 2 О |
о W |
ОС |
W ос' |
МП |
04 04 |
40 МП W |
С4 W |
W МП 04 |
С4 o' о W |
of W |
04^ ? |
ОО of |
О 40 04 |
|
ft о 2 2 & И ч -к 2 « S К ® w ^ |
МП ОО W |
СП |
МП 04" ОО |
ОС 04 W |
Г1 МП W |
W 04 04 |
ю. 04 04 |
o' 04 W |
W |
40 |
МП |
О' 04 |
of О 04 W |
|
£ X | § ж П о К ^ |
МП |
ОО |
04 o' 04 |
МП 04^ W |
S СП |
W МП МП |
О МП МП |
40 04^ 04 04 |
04 |
w МП W |
o' |
О МП |
0^ 04 04 |
|
св Л св ы о & с о и св СП Щ о X а |
04^ ГП |
04 |
40 ОС |
МП |
ю. o' 40 w |
W МП МП |
о МП МП |
40 ^1 04 04 |
04 |
W МП W |
o' |
^ МП |
40 МП |
|
S X св S = й к 2 °х йхо 2 й us ч g о g ® н И и 5 2ft и св |
ОО o' |
W О |
04^ |
мп o' |
О o' |
1 |
1 |
1 |
1 |
1 |
1 |
1 |
04 W |
Таблица 4.
ЭНЕРГЕТИЧЕСКИЙ БАЛАНС Alhagietum–Artemisosum НА ОСНОВЕ ДАННЫХ
ПО ДИНАМИКЕ РАСТИТЕЛЬНОЙ МАССЫ И СУММАРНОГО ИСПАРЕНИЯ ЗА 2015–16 гг.
|
I |
II |
III |
IV |
V |
VI |
VII VIII |
IX |
X |
XI |
XII |
Год |
||
|
2015 |
|||||||||||||
|
a s SEO а л 2 н се g се S 2< инн Я се о |
CO w |
40 |
СЧ о |
40 ci w |
40 40 |
04^ OO w |
ci w |
tn 40 ci w |
tn o' |
40 OO |
^ tn |
OO |
о co |
|
И Л л ® К О Л л о, h а м 2 C|ggs^g |
co w |
w ci |
in о |
CO 04^ |
40 |
OO |
^4 OO co |
||||||
|
a J^ftH ® s Ba«sH |
co tn ci |
co |
tn ^ ?—< |
°1 ?—< |
tn о ?—< |
40 o' |
co oi |
||||||
|
h L i о Й Os h 6 « Н К ^s aSS 4 |
OO CO w |
о oi 40 ?—< |
40 ci |
40 OO co |
OO |
СЧ 40 О |
OO 04 CO |
°\ 40 OO co |
OO co |
OO 04 ?—< |
^ OO co |
CO |
40 о OO co |
|
и ® н g g о о ейвкйощЯий ^ В & И EI |
°°. OO CO |
ci tn |
o^ 04 |
40 w |
40^ ci |
°\ CO w |
?—< 40 |
||||||
|
^'lH4si о &« &o д ц |
in tn |
ci tn |
Cl |
co |
°s. co |
°\ |
^ CO |
||||||
|
О § |
ci |
tn о in |
40 OO |
co 40 |
oi co |
c |
?—< |
о |
?—< |
°1 |
tn |
||
|
g * о 3S 2 s^ ^S |
tn ci w |
OO co |
tn ci 40 |
oo 40 |
СЧ tn |
oo w |
40 OO |
°\ tn |
co |
°\ |
о |
о 40 |
|
|
§ s 2 6 ■ 2 2 о ° &2 о St[ о £*gs S£« |
О ci |
co 40 |
tn СЧ tn |
co 40 |
OO w |
40 tn |
OO tn o' co |
w ?—( |
Cl o' ?—< |
^ tn |
co ^ |
tn ^ co |
|
|
ce S 6 2 У' &3 S ш ^2 ^sB s^i^ m й С о w “ |
w |
CO 04 in |
СЧ о w |
OO co w |
°\ о 04 |
tn CO |
OO tn o' co |
w ?—( |
Cl o' |
4 tn |
co |
tn ^ ri |
°1 40 OO |
|
gsg&^^gggBo o^S^§^^« |
о o^ о |
04 OO o' |
40 OO О |
04^ о |
О 04^ о |
tn oo o' |
1 |
1 |
1 |
1 |
1 |
1 |
04 °^ o' |
|
I |
II |
III |
IV |
V |
VI |
VII |
VIII |
IX |
X |
XI |
XII |
Год |
|
|
2016 г. |
|||||||||||||
|
К K’S -S н о s □ К о ? а ац в н Л a s |
40 40 мп |
мп |
о W |
МП ОО оо" |
о W |
W W |
МП 40^ со" W |
МП о |
40 ос' |
40 МП |
ОО ^ |
со W |
|
|
Б к ’К К «X 5^ а н И о a S'-'M R-^ o’S Cq^s“*§8ck |
мп W |
ОО W |
40 |
4о" |
оо" |
МП оо" |
х? ей |
||||||
|
1 о s ° 5 о | S о а н И л s А п®2ййЬаК £ ^*&§§ |
04 W |
40 С4 ?—< |
40 ri |
со" |
40 МП ?—< |
со ?—< |
^ |
||||||
|
g^LaSg 1g h й к о о О ЙЙййилНИ« Г) й Ч & « R |
ОО со W |
о о? 40 w |
40 со |
о о со |
СО' о МП со |
оо ОО |
оо" 04 СО |
°\ 40 ОО со |
оо" со |
ОО 04 ?—< |
Ю. оо" со W |
с© |
о ОО со |
|
S й S g н ^ K4gagg Ч |
04 |
СО |
МП ОО |
40 W |
°\ оо" ОО W |
04 ?—< |
04 40 |
||||||
|
§ £ а £ н % k о § н S 8 * о «чЙ“йдй^ ^ | ^ё « |
40^ о |
МП |
40^ о 40 |
МП МП |
мп" |
со |
О^ СО |
||||||
|
к ч 2 3 2 о |
W о W |
X? |
W оо" |
МП |
со 04 |
40^ МП ?—< |
°\ ?—< |
?—< МП со |
С4 О о ?—< |
со" |
°1 |
ОО хг" |
о 40 04 |
|
i о 8 | айв „ s ё «К К ми^ |
МП ОО W |
ГП |
МП ci ОО |
оо" со W |
СЧ МП ?—< |
?—< со со |
40^ о? со |
о со W |
?—< |
40 |
О со |
oi о со W |
|
|
Or О О |sghpS П О s Й |
МП |
ОО |
СО о со |
МП сч W |
ГП |
?—< МП МП |
о МП МП |
40 СЧ oi со |
S со |
?—< МП ?—< |
о" ?—< |
мп |
О' 04 СО |
|
Й S ° И о ей К о 2 4 |
04 |
О МП |
МП W |
W |
СО со |
о 04" со |
МП МП |
со |
С4 со |
МП ?—< |
ОО о" |
<0 МП |
°1 МП ОО 04 |
|
□ щККчй®к§о^ 0л^^?,й>йНИи0 S^O^^°oU^H^ Он ^®“й^и |
04 ОО О |
ОО о |
о ОО о |
СО оо о |
МП оо о |
г? о |
1 |
1 |
1 |
1 |
1 |
1 |
^ о" |
Для полынно–верблюжьеколючковой ассоциации затраты энергии на почвообразование составили 786,2 кДж и 985,2 кДж. Энергия расходованная на биологические процессы соответственно составили 641,2 кДж и 691,3 кДж. Относительная доля формирования растительного вещества составили 0,89% и 0,83% (Таблица 4).
Итак, приведенные количественные показатели энергетических расходов в растительных сообществах за период исследований охватили две разные погодноклиматические условия, резко отличающиеся между собою.
Засушливый период — 2015 г., был заменен переувлажненным периодом — 2016 г., что позволило составить энергетический баланс биогеоценозов двух контрастных периодов.
Список литературы Продуцирование и энергетические показатели биогеоценозов полупустынь Ганых-Айричайской долины Азербайджана
- Базилевич Н. И. Биологическая продуктивность экосистем Северной Евразии. М.: Наука, 1993. 292 с.
- Алиев С. А. Экология и энергетика биохимических процессов превращения органического вещества почв. Баку: Элм, 1978.
- Титлянова А. А. Биологический круговорот азота и зольных элементов в травяных биогеоценозах. Новосибирск: Наука, 1979.
- Дояренко А. Г. Использование солнечной энергии полевыми культурами // Научно-агрономический журнал. 1924. №4. С. 259-268.
- Будагов Б. А. Геоморфология южного склона Большого Кавказа: (в пределах Азербайджанской ССР). Баку: Элм, 1969. 178 с.
- Мадатзаде А. А., Шихлинский Э. М., Кавецкая Г. Г. Климат Азербайджана. Баку: Изд-во АН АзССР, 1968. 343 с.
- Кузнецов Н. И. Принципы деления Кавказа на ботанико-географические провинции: (Доложено в заседании Физ.-мат. отд. 23 янв. 1908 г.). СПб., 1909. 174 с.
- Гроссгейм А. А. Растительный покров Кавказа. М., 1948. 267 с.
- Гаджиев В. Д. Субальпийская растительность Большого Кавказа (в пределах Азербайджанской ССР). Баку: Изд-во Акад. наук АзССР, 1962. 172 с.
- Прилипко Л. И., Родин Л. Е., Маилов Е. М. Динамика растительности горных лесных лугов Большого Кавказа. Баку, 1972.
- Волобуев В. Р. Введение в энергетику почвообразования. М.: Наука, 1974. 128 с.
- Зейналов Ю. А. Энергетика почв и агрофитоценозов в условиях Восточной Ширвани: дисс.. канд. с.-х. наук. Баку, 1980. 258 с.
- Керимов А., Самедов П. Экологические и энергетические пути повышения производительности почв. Их проблемы и прикладное значение. 2019. M.: LAP LAMBERT Academic Publishing.
- Базилевич Н. И., Родин Л. Е. Географические закономерности продуктивности и круговорота химических элементов в основных типах растительности суши // Общие теоретические проблемы биологической продуктивности. М.: Наука, 1969.
