Провизорность эндокринной части поджелудочной железы у крупного рогатого скота в эмбриогенезе

Автор: Хонин Геннадий Алексеевич, Гичев Юрий Матвеевич, Семченко Валерий Васильевич
Журнал: Вестник Омского государственного аграрного университета @vestnik-omgau
Рубрика: Ветеринарные науки
Статья в выпуске: 2 (22), 2016 года.
Бесплатный доступ
В результате гистологического, морфометрического и гистохимического исследования эндокрин- ной части поджелудочной железы у крупного рогатого скота в эмбриональном, раннем постнатальном периоде развития и у взрослых животных (112 зародышей, предплодов и плодов в возрасте от 30 до 270 сут, 12 новорожденных телят в возрасте от 12 ч до 30 сут и 3 коровы красной степной породы; после фиксации в нейтральном 10 %-ном формалине, жидкости Карнуа и ацетоне целые эмбрионы или кусочки органа уплотняли путем заливки в парафин; полученные на роторном микротоме срезы окрашивали ге- матоксилином и эозином, хромовым гематоксилином по Баргману, альдегид-фуксином по Габа, по До- миничи-Кедровскому; гликоген выявляли по Шабадашу, кислую и щелочную фосфатазу методами Го- мори; измеряли площадь эндокринных островков и инсулоцитов В; цифровые данные обработаны стати- стически), выполненного в морфологической лаборатории Института ветеринарной медицины и биотех- нологии Омского ГАУ, установлено, что первые эндокриноциты выявляются в зачатке поджелудочной железы зародышей 30-суточного возраста. Эти эндокриноциты по морфологическим и тинкториальным свойствам подобны эндокринным панкреоцитам А. Появление и специфическая дифференцировка эндо- кринных панкреоцитов А происходит до начала формирования системы протоков и ацинусов экзокрин- ной части органа, а в процессе формирования структур экзокринного отдела они находятся в их составе. Эти клетки дают начало островковым почкам и в последующем эндокринным островкам. Эндокринные панкреоциты В в формирующихся островках появляются в конце второго месяца эмбриогенеза. В про- цессе развития эндокринной части поджелудочной железы происходит смена двух генераций панкреати- ческих островков. Островки первой генерации развиваются между дольками экзокринного отдела или на краю его долек, где обеспечиваются лучшей васкуляризацией и вступают в контакт с нервными ганглия- ми. Наибольшее развитие этих островков происходит в последнем триместре беременности, когда осу- ществляется максимально интенсивный рост плода. В конце беременности начинается инволюция этих островков, которая завершается в постнатальный период. Островки второй генерации являются предше- ственниками дефинитивных островков. Они формируются внутри долек и отличаются от островков пер- вой генерации меньшими размерами и признаками невысокой функциональной активности. В течение эмбрионального периода развития и у взрослых животных эндокринные островки не утрачивают морфо- логическую взаимосвязь с ацинусами и мелкими протоками поджелудочной железы. Островки первой генерации являются провизорными в морфологическом и функциональном отношении структурами. Со- стоящие преимущественно из эндокринных панкреоцитов В, они, вырабатывая большое количество ин- сулина, обеспечивают снабжение интенсивно растущих тканей и органов плода глюкозой, а после рож- дения, когда интенсивность роста снижается, подвергаются обратному развитию.
Эндокринная часть поджелудочной железы, крупный рогатый скот, эмбриональ- ный и постнатальный онтогенез, провизорность
Короткий адрес: https://sciup.org/142199167
IDR: 142199167 | УДК: 619:611.013(043.3):636.2
The provisionality of endocrine part of cattle’s pancreas in embryogenesis
By histological, morphometrical and histochemical methods were investigated endocrine part of the pancreas of cattle in embryonic, early postnatal development and in adult animals. After fixation in 10% neutral formalin solution, Carnoy’s fluid and acetone, whole embryos or parts of their pancreas were embedded in paraffin. The sections obtained on a rotary microtome were stained with hematoxylin and eosin, chrome hematoxylin sensu Bargmann, aldehyde fuchsin sensu Gaba, sensu Dominici-Kedrovsky; glycogen was detected sensu Shabadash, acide and alkaline phosphatase - sensu Gomory methods. The area of endocrine islets and insulocytes B was measured. The results of measurement were processed statistically. The work was performed in the morpho- logical laboratory of Institute of Veterinary Medicine and Biotechnology of Omsk State Agrarian University. It was found that the first endocrinocytes were detected in pancreatic primordium of embryos of 30 days age. Morphological and tinctorial properties of these endocrinocytes are similar to pancreocytes A of definitive islets. The appearance and specific differentiation of endocrine pancreocytes A occurs before the formation of the tem of ducts and acini of pancreatic exocrine part and after the begining of its formation these cells are situated in epithelial layer of exocrine structures. These cells give rise to islets buds and, subsequently, endocrine islets. Endocrine pancreocytes B in pancreatic islets appear at the end of the second month of embryogenesis. In the development of the endocrine part of the pancreas there was a change of two generations of pancreatic islets. The islets of first generation developed between lobules of exocrine pancreas or at its edges. This place provides a better vascularisation and contact with the nerve ganglia. The greatest development of these islets occured in the last trimester of pregnancy, when done the most intensive growth of the fetus. At the end of pregnancy began involution of these islets, which culminated in the postnatal period. Islets of second generation are the precursors of definitive islets. They are formed inside the lobules and distinct from first generation islet by smaller area and features of low functional activity. During the period of embryonic development and in adult animals endocrine islets do not lose morphological relationship with small ducts and acini of exocrine pancreas. The islets of the first generation are provisional structure. Consisting mainly of endocrine pancreocytes B, they produce large amounts of insulin, ensure the supply intensively growing tissues and organs of the fetus by glucose, and after the birth, when the growth rate is reduced, these islets regress.
Текст научной статьи Провизорность эндокринной части поджелудочной железы у крупного рогатого скота в эмбриогенезе
Проблема изучения механизмов, обеспечивающих процесс реализации генетической информации и интеграции всех систем организма в его развитии, остается одной из наиболее важных проблем биологии развития животных и человека. Наряду с иммунологическим и генетическим контролем роста и дифференцировки органов и тканей [1, 5], межтканевыми индуктивными взаимодействиями [6], гуморальная регуляция является одним из наиболее важных интегрирующих механизмов в процессе эмбрионального развития. Раннее развитие органов эндокринной системы плодов млекопитающих и ранний синтез в них биологически активных гормонов определены значительной автономностью эмбрионов и относительной независимостью регуляции обмена веществ и дифференцировки их органов от гормонов материнского организма. Именно этим можно объяснить и провизорность в развитии многих органов и систем в эмбриональных морфогенезах [1]. Факт более ранней дифференцировки эндокринного аппарата поджелудочной железы, по сравнению с ее экзокринной частью, является общепризнанным. Доказана и функциональная активность эндокринных клеток в процессе эмбрионального развития. Вместе с тем, многие вопросы, связанные с гистогенезом эндокринной части органа, остаются изученными недостаточно, в том числе у крупного рогатого скота.
В задачу исследования входило изучение морфофункционального развития эндокринного аппарата поджелудочной железы с момента появления первых эндокринных клеток до дефинитивного состояния органа.
Объекты и методы
Для разрешения поставленной задачи исследована поджелудочная железа 112 зародышей, предплодов и плодов в возрасте от 30 до 270 сут, 12 новорожденных телят в возрасте от 12 ч до 30 сут и 3 коров красной степной породы. Фиксированные в нейтральном формалине, жидкости Карнуа и ацетоне целые эмбрионы или кусочки органа уплотняли путем заливки в парафин. Полученные на роторном микротоме срезы окрашивали гематоксилином и эозином, хромовым гематоксилином по Баргману, альдегид-фуксином по Габа, по Доминичи-Кедровскому. Гликоген выявляли по Шабадашу, кислую и щелочную фосфатазу методами Гомори. Измеряли площадь эндокринных островков и инсулоцитов В. Цифровые данные обработаны статистически. Работа выполнена в морфологической лаборатории института ветеринарной медицины и биотехнологии Омского ГАУ.
Результаты исследования
Исследования показали, что на 30-е сутки эмбрионального развития в зачатке поджелудочной железы, состоящем из тяжей недифференцированных клеток, отходящих от энтодермы средней части первичной кишки в дорсальную брыжейку, обнаруживаются клетки с признаками дифференцированных эндокриноцитов. Эти клетки имеют вытянутую форму, в их цитоплазме, обращенной к кровеносному капилляр, имеется мелкая оксифильная зернистость. По морфологическим и тинкториальным свойствам эти клетки похожи на А-клетки дефинитивной поджелудочной железы. В ранний предплодный период (35–50 сут) клеточные тяжи превращаются в эпителиальные трубочки, в клеточном составе которых содержатся такие же оксифильно- зернистые клетки. В этот же период названные клетки начинают формировать скопления (почки) и обособляться от экзокринных элементов в виде островков, которые не утрачивают с ними связь. В начале позднего предплодного периода (51–60-е сут) в островках при окраске альде-гид-фуксином начинают выявляться В-клетки, их количество постепенно увеличивается, но оно остается до конца предплодного периода меньшим, чем количество оксифильнозернистых клеток, которые считают А-клетками. На более раннее, по сравнению с инсулином, появление в зачатке поджелудочной железы глюкагона указывает Asplund K. et al. Возможно, более ранняя функция А-клеток связана с необходимостью потребления зародышем, не связанным еще плацентой с материнским организмом, углеводов из мест депонирования гликогена, которым в это время является энтодерма первичной кишки [2].
К 90-м суткам развития плода в поджелудочной железе формируется дольчатое строение. Одновременно с развитием дольчатости начинают проявляться различия в структурнофункциональном развитии эндокринных островков, находящихся в междольковой соединительной ткани, и островков, находящихся внутри долек среди экзокринной паренхимы. Островки, расположенные вне долек или по их периферии, формировались раньше, чем внут-ридольковые, в связи с чем мы их называем островками первой генерации, а островки внутри долек – островками второй генерации.
Островки первой генерации вначале состоят из широких разнообразной формы клеточных тяжей, в которых постепенно начинают преобладать В-клетки. А-клетки расположены в большем количестве в участках, связанных с экзокринными элементами. С возрастом плода клеточные тяжи становятся более тонкими, разветвленными, между ними появляется большое количество синусоидно расширенных и заполненных кровью капилляров. С 3-го по 5-й месяц эмбриогенеза площадь островков первой генерации существенно возрастает, а к концу 9-го месяца она достигает максимального значения (таблица). С самого начала формирования островки первой генерации имеют тенденцию сближения с расположенными в междольковой соединительной ткани нервными ганглиями, которые оказываются заключенными внутри некоторых островков, формируя так называемые нейро-инсулярные комплексы.
Эндокринные панкреоциты А и В с начала раннего плодного периода проявляют признаки активной инкреторной деятельности, которая усиливается к началу пятого месяца развития и достигает максимума в поздний плодный период. На активную функцию островков первой генерации указывает увеличение и переполнение кровью капилляров, наличие морфологических признаков активной инкреторной функции А- и В-клеток, а также интенсивная реакция на кислую и щелочную фосфатазы. Усиление инкреторной функции островков первой генерации в поздний плодный период сопровождается интенсификацией апоптоза эндокриноцитов и замещением их жировыми клетками. Инволютивные процессы особенно выражены у 9-месячных плодов. В течение первых суток постнатального онтогенеза островки первой генерации сохраняют признаки активной функции и, вместе с тем, в них становятся более выраженными деструктивные процессы. У взрослых животных островки первой генерации не выявляются. Все дефинитивные островки находятся внутри долек и сохраняют морфологическую взаимосвязь с ацинусами и внутридольковыми выводными протоками.
Островки второй генерации отличаются от островков первой генерации по топографии и строению (рис. 1, 2). Размер их меньше, они круглые или овальные, более компактные, их немногочисленные капилляры менее гиперемированы и расположены только по периферии островков. Эндокринные островки второй генерации в течение всего эмбриогенеза размещаются только внутри долек и тесно связаны с элементами экзокринной части перешейками. Тесная взаимосвязь с элементами экзокринной паренхимы, размещение внутри долек, размеры и форма типичны и для островков поджелудочной железы взрослых животных. Последнее позволяет считать островки второй генерации предшественниками дефинитивных островков. Увеличение эндокринных островков второй генерации происходит медленнее, чем увеличение островков первой генерации (таблица).
Эндокринные клетки островков второй генерации в эмбриональный период проявляют черты специфической дифференцировки. В них имеется А- или В-зернистость, однако признаки инкреторной активности в них выражены слабо. В течение всего эмбриогенеза В-клетки островков второй генерации отличаются от В-клеток первой генерации меньшим размером (таб- лица), невыраженной полярностью в строении, а также меньшей активностью и несколько иной цитотопохимией гидролитических ферментов.
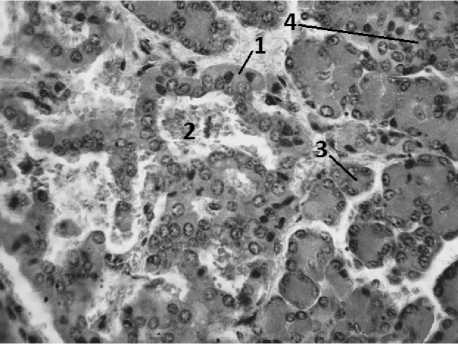
Рис. 1. Поджелудочная железа 8-месячного плод:
1 – островок первой генерации; 2 – синусоидно расширенные кровеносные капилляры; 3 – ацинусы экзокринной части; 4 – островок второй генерации. Окраска хромовым гематоксилином и эозином. Ув. 200х
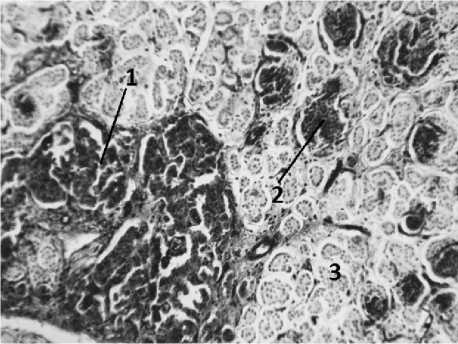
Рис. 2. Реакция на щелочную фосфатазу в поджелудочной железе 8-месячного плода:
1 – островок первой генерации; 2 – островок второй генерации; 3 – ацинусы экзокринной части.
Метод Гомори. Ув. 50х
Площадь эндокринных островков и инсулоцитов В в эмбриональной и дефинитивной поджелудочной железе у крупного рогатого скота (M ± m, мкм2)
|
Возраст, сутки |
Площадь островков |
Площадь инсулоцитов В |
||
|
Островки 1-й генерации |
Островки 2-й генерации |
Островки 1-й генерации |
Островки 2-й генерации |
|
|
90 |
15755 ± 613 |
2086 ± 312** |
50,6 ± 2,2 |
28,7 ± 0,8** |
|
120 |
38757 ± 842* |
2486 ± 281* ** |
– |
– |
|
150 |
383113 ± 9906* |
4131 ± 397* ** |
66,3 ± 2,5* |
32,9 ± 1,1* ** |
|
180 |
404876 ± 8921* |
5766 ± 408* ** |
– |
– |
|
210 |
465970 ± 21411* |
6008 ± 443* ** |
78,9 ± 3,3* |
39,7 ± 1,4* ** |
|
240 |
783576 ± 20573* |
5980 ± 420* *** |
– |
– |
|
270 |
1422191 ±42176* |
7792 ± 488* ** |
97,0 ± 3,8* |
31,7 ± 1,2* ** |
|
Взрослые животные |
– |
17105 ± 651* ** |
– |
– |
*Различия достоверны (р < 0,05) между показателями в разные сроки в сравнении с показателями через 90 суток; **различия достоверны (р < 0,05) между показателями второй и первой генерации эндокринных островков.
Наличие в поджелудочной железе плодов крупного рогатого скота эндокринных островков двух генераций, отличающихся друг от друга топографическим местоположением с сохраняющимися в течение всего плодного периода различиями в морфофункциональной дифференцировке их клеток, позволяет высказать предположение о различной их функциональной значимости в развитии плода. Островки первой генерации, по данным исследования, являются структурами провизорными, обеспечивающими повышенную выработку гормонов поджелудочной железы, особенно инсулина, в период наиболее интенсивного роста массы плода и развития его внутренних органов. По мере снижения потребности в повышенном количестве инсулина, возникающей после рождения, эти островки подвергаются обратному развитию.
Островки второй генерации являются предшественниками дефинитивных островков, на что указывают их топография и форма. Однако, несмотря на отсутствие в них ярких признаков инкреторной деятельности, они не являются структурами индифферентными, выполняя большую роль в процессах регуляции развития, дифференцировки экзокринных элементов в плодный период развития. Островки, рассеянные среди экзокринной части железы, расматриваются как центры, стимулирующие митотическую активность, регулирующие процессы синтеза и выделения секрета экзокринными панкреоцитами [1]. В исследованиях также отмечалось одно- временное с появлением в эндокринных островках В-клеток образование зимогенных гранул в экзокринных панкреоцитах.
Наиболее быстрое и интенсивное развитие, а также наиболее активная функция островков первой генерации в значительной мере объясняется их местоположением в междольковой соединительной ткани, где они обеспечиваются лучшим кровоснабжением и иннервацией, образуя нейро-инсулярные комплексы. Очевидно, что столь тесная взаимосвязь нервных ганглиев и эндокринных островков первой генерации неслучайна. С одной стороны, лучшая иннервация этих островков проявляется их более активной функцией, а, с другой стороны, близость к источнику инсулина способствует развитию ганглиоцитов. За последние годы доказано [6], что окружающая мезенхима оказывает регулирующее влияние на пролиферацию и дифференцировку клеток эндокринной части поджелудочной железы. Островки первой генерации находятся в более тесном, чем островки второй генерации, контакте с окружающей их эмбриональной соединительной тканью.
Выводы
Островки первой и второй генерации в процессе эмбриогенеза являются провизорными с точки зрения не только структуры, но и функции образованиями.
Островки первой генерации являются основными источниками гормонов поджелудочной железы во внутриутробный период развития и после рождения подвергаются инволюции.
Островки второй генерации в плодный период обеспечивают индуцирующее воздействие на рост и дифференцировку экзокринной части поджелудочной железы.
Провизорными следует считать у 30-суточных зародышей оксифильнозернистые клетки, являющся аналогами дефинитивных А-клеток.
Список литературы Провизорность эндокринной части поджелудочной железы у крупного рогатого скота в эмбриогенезе
- Принцип провизорности в морфогенезах/Г.С. Соловьев, В.Л. Янин, В.Д. Новиков, С.М. Пантелеев. -Тюмень: Академия, 2004. -127 с.
- Тельцов, Л.П. Полисахариды эпителия тонкого кишечника в эмбриогенезе/Л.П. Тельцов//Научные труды Омского ветеринарного института, 1971. -T. 28, Вып. 1. -С. 81-86.
- Утехин, В.И. Особенности синтеза белков в клетках различных отделов экзокринного эпителия поджелудочной железы/В.И. Утехин//Бюллетень экспериментальной биологии, 1974. -Вып. 3. -С. 63.
- Function of the fetal e Function of the fetal endocrine pancreas/K. Asplund, A. Andersson, C. Jarrus K. Asplund, A. Andersson, C. Jarrusse, C. Hellerstrom//Isr. J. Med. Sci. -1975. -V. 11, № 6. -Р. 581-590.
- Bona C. Genes controlling Bona, C. Genes controlling pancreas ontogeny / C. Bona, P.L. Herrera // Int. J. Dev. C. Bona, P.L Herrera // Int. J. Dev. Biol. : 2008, 52(7) ; 823-35 PMID :18956314.
- Pancreatic mesenchyme regulates epithelial organogenesis throughout developme organogenesis throughout development / L. Landsman et al. // PLoS Biol. : 2011, 9 (9) ; al. // PLoS Biol. : 2011, 9 (9) ; e1001143 PMID : 21909240|PLoS Biol. (http://ww 823-35 PMID : 18956314. 6. Pancreatic mesenchyme 21909240|PLoS Biol. (http://www.plosbiology.org/article/info:doi/10.1371/journal.pbio.10ticle/info:doi/10.1371/journal.pbio.1001143).
