Псевдоморфообразование по раковинам силурийских брахиопод

Автор: В.И. Каткова, Т.М. Безносова, В.Н. Филиппов
Журнал: Известия Коми научного центра УрО РАН @izvestia-komisc
Статья в выпуске: 3 (49), 2021 года.
Бесплатный доступ
Структурными и оптическими методами проведено исследование псевдоморфоз кварца по створкам четырех раковин раннесилурийских брахиопод Borealis sр. отряда Pentamerida из карбонатной толщи гряды Чернышева. Окременение стенки раковин брахиопод привело к полной замене первоначального карбонатного скелета с изменением его структурной организации. Установлено, что основной минеральной составляющей внутрираковинного вещества и вмещающей породы является кварц. В работе обсуждается участие раннепалеозойских и современных цианобактериальных симбионтов в процессах замещения и формирования первичных и новообразованных кристаллических агрегатов кварца на стенках раковин.
Псевдоморфозы, брахиоподы, створки, раковины, силур, кварц, минерал, кристалл
Короткий адрес: https://sciup.org/149132182
IDR: 149132182 | УДК: 548.6:564.8 | DOI: 10.19110/1994-5655-2021-3-49-55
Pseudo-morphogenesis in the shells of the Silurian rachiopods
The analysis of paleontological data, the study of mineral matter transformation involving live organisms in various rocks is important for the understanding of the events of the geological past. The aim of this work is to identify the bio-genic factor in the process of pseudo-morphogenesis in the shell sashes of the Early Silurian brachiopods from the carbonate rocks of the Chernyshev Ridge. Structural and microscopic methods (Shimadzu XRD 6000; Fourier spectrometer InfraLum FT-02; JSM 6400 JEOL; VEGA3 TESCAN; MIN-8) were used to study quartz pseudomorphoses in the sashes of four shells of the Early Silurian brachiopods Borealis sр. of Pentamerida order from the carbonate strata of the Chernyshev Ridge. The salicification of the brachiopod shell walls resulted in the complete replacement of the original carbonate skeleton with a change in its structural organization. It is established that quartz is the main mineral component of the intra-shell material and secondary dolomite crystals and newly formed calcite aggregates. The paper discusses the participation of Early Paleozoic and modern cyanobacterial symbionts in the replacement of primary calcite with quartz and the formation of quartz crystalline aggregates on the shell walls. It is shown that quartz on the surface of shell sashes and in the intra-shell substance of brachiopods has under-gone repeated transformations due to the combination of ancient and modern weathering processes
Текст научной статьи Псевдоморфообразование по раковинам силурийских брахиопод
Процессы фоссилизации нередко приводят к образованию различных типов псевдоморфоз по останкам организмов. Морфологические особенности и установление условий формирования фосси-лий всегда привлекали внимание минералогов. Исследование фоссилизированных скелетных организмов актуально не только для палеонтологов и стратиграфов, проводящих реконструкции палеоэкосистем, литологов для изучения геохимической обстановки литификации осадков, но и биоминералогов, исследующих биоминеральные взаимодействия в седиментогенезе. Анализ палеонтологических данных, исследование преобразований минерального вещества с участием живых организмов в различных породах важно для познания событий геологического прошлого. Фоссилизированные структурные элементы и формы биоорганизмов после захоронения становятся составной частью породы и часто подвергаются процессам минерализации или замещения. Однако варианты замещения и роста новообразованных кристаллов в псевдоморфозах остаются недостаточно изученными.
Ранее проведенные исследования показали, что в отложениях нижнего силура на гряде Чернышева встречаются окремененные раковины брахио-под удовлетворительной сохранности [1]. Одними из самых изученных представителей палеозойских морских беспозвоночных в Тимано-Североураль-ском палеобассейне являются брахиоподы отряда Pentamerida: Virgiana, Borealis, Pentamerus. Их раковины состоят из двух неравных створок — брюшной и спинной (педальной и брахиальной). Стенка раковины замковых брахиопод может состоять из одного, двух или трех слоев [2–4]. В работе В.П. Сапельникова [2] приняты следующие наименования слоев раковин пентамерид: внешний пластинчатый, призматический и внутренний пластинчатый. Внешний карбонатный слой считается условно первым после периостракума, редко сохранившимся на раковинах. У пентамеридин развит призматический слой. Автор отмечает [2], что типы слоев различаются внутри отдельных групп пентамерид. В частности, в брюшных створках у пентамерид развиты два слоя: наружный волокнистый и внутренний призматический, состоящий из кристаллов кальцита, ориентированных перпендикулярно к створкам раковины. Внутрираковинные структурные элементы (септа, зубы, пластины) также состоят из карбоната кальция.
Целью работы является выявление биогенного фактора в процессе псевдоморфообразования по створкам раковин раннесилурийских брахиопод из карбонатных пород гряды Чернышева.
Методы и материалы исследований
Объектами наших исследований послужили брюшные створки раковин, их фрагменты, внутрира-ковинное вещество четырех раковин брахиопод Borealis sр. отряда Pentamerida, извлеченных из бра-хиоподовой банки. Средняя длина створки составляет 2.5 см, а ширина – 2 см. Массовое захоронение раковин этих брахиопод было обнаружено в разрезе нижнего силура гр. Чернышева в бассейне р. Шаръю. Брахиоподы Borealis sр. в захоронении представлены преимущественно крупными особями с толстостенными раковинами, а также отдельными створками и их фрагментами. Исследования минерального вещества брахиопод проведены оптическими (МБС-10; МИН-8), рентгеноструктурным (рентгеновский дифрактометр Shimadzu XRD 6000) и ИК-спектроскопическим (фурье-спектрометр Инфра-Люм ФТ-02) методами. Морфологические особенности и химический состав минеральных фаз изучены с использованием СЭМ (JSM 6400 JEOL, VEGA3 TESCAN) и микрозондового анализа. Компонентный состав элементов внутрираковинного вещества и вмещающей породы в валовых пробах в виде порошка определен на приборе XRF-1800 (Shimadzu). Изучение образцов проводилось на базе ЦКП УрО РАН «Геонаука» ИГ Коми НЦ УрО РАН (г. Сыктывкар).
Результаты исследований
Структурными методами установлено, что наружная поверхность створок четырех раковин брахиопод имеет мономинеральный состав и представлен кварцем. Анализ СЭМ-изображений показал, что вместо структурно упорядоченного карбонатного слоя стенки на наружной поверхности одного из створок размещаются кристаллы кварца с формами положительного, отрицательного ромбоэдра и гексагональной призмы. Кристаллы имеют короткопризматический облик, а ямки и каверны на их гранях указывают на процессы травления. Кварц формирует сплошную кристаллическую щетку, состоящую из индивидов размерами от 5 до 50 мкм (рис. 1). Наряду с кварцевой щеткой визуализируются зоны, сформированные из неориентированных индивидов разных генераций. Фигуры травления на поверхностях призм и ромбоэдров имеют как неправильные, так и геометрические формы.
На гранях отдельных кристаллов и между индивидами выявлены редкие реликты биомине-ральных пленок со следовыми содержаниями алюминия и кремния. Полагаем, что их можно рассматривать как следы жизнедеятельности бактериальных сообществ в постседиментационный период (рис. 2).
Согласно микроснимкам РЭМ на поверхности двух других раковин преобладает ячеистая структура, сформированная из кварцевых сростков с диаметром 200–400 мкм (рис. 3). Размеры ограненных индивидов в агрегатах увеличиваются по мере удаления от центра сростка к периферии (рис. 4). Кварц можно наблюдать также в виде отдельных двухголовчатых индивидов между ячейками. На вершинах кристаллов в агрегатах часто наблюдаются ямки травления. Кроме того, следует отметить о явлениях выраженного растворения сростков, при которых видоизменяются облик и морфология индивидов.
Между друзами встречается обильное развитие внеклеточного полимерного вещества, представленное как гликокаликсом, так и чехлами современных цианофитов, разделяющих между собой

Рис. 2. Реликты окремененного гликокаликса между кристаллами кварца.
Fig. 2. Remnants of the silicified glycocalyx between quartz crystals.

Рис. 1. Кристаллы кварца разных генераций на поверхности створки брахиопода Borealis sр.
Fig. 1. Quartz crystals of different generations on the surface of the sashe of the brachiopod Borealis sp.

Рис. 3. Общий вид фрагмента створки раковины брахиопода Borealis sр.(поперечное сечение брюшной створки).
Fig. 3. General view of the fragment of the shell sashe of the brachiopod Borealis sp.
ячейки (рис. 3). В составе внеклеточного органического вещества цианофитов зафиксированы следовые содержания Mg, Si, Ca и Al.
Внутрираковинное вещество под тонким упорядоченным слоем кристаллов кварца представлено субмикрозернистым кремнеземом, в матрицу которой включены индивиды доломита, сростки и двухголовчатые монокристаллы кварца, а также агрегаты кальцита. Кроме того, в структуре кварцевой матрицы зафиксированы фрагменты гелеподобных кремнийсодержащих пленок. Кварц, как правило, является основным минералом, заместившим створки раковин брахиопода. Монокристалльные зерна кварца и сростки заполняют трещины в кремнеземе. При изучении внутрираковинного вещества в прозрачных шлифах выявлено, что кристаллы вторич-

Рис. 4. Ячеистая структура кварца на его поверхности.
Fig. 4. Cellular structure of quartz on its surface.
ного доломита чаще всего ромбической формы располагаются в кварцевой матрице неравномерно, образуя местами скопления различной формы и величины. Например, в одном из шлифов обнаружено, что одна половина створки полностью замещена кремнеземом, а на другой рассеяны индивиды доломита в микрозернистом кварце. На РЭМ-изображениях визуализируются частично корродированные кристаллы доломита ромбоэдрического облика размерами 50–100 мкм. Индивиды вторичного кальцита формируют зернистые агрегаты. В строении кристаллов кальцита проявляется слоистость, а морфология поверхности отдельных кристаллов внутри створки указывает на то, что они также подвергались травлению.
На микроснимках внутрираковинного вещества обнаружены множественные чехлы современных цианофитов (рис.5). Исследования показали, что частично окремненные чехлы имеют состав, мас.%: Si 11-20; Ni 1,64; S 0,97; Cu 0,89; Ca 0,45; Al 0,25. Следует отметить, что они распространены, как правило, в кварцевых прослойках. Наряду с ними встречаются гифы грибковой микрофлоры (актино-мицеты?). Формирование различных минеральных пленок и фрагментов «рубашек», содержащих оксиды железа на кристаллах кварца, свидетельствует об участии бактерий в образовании новообразованных минералов во внутренней полости раковин.
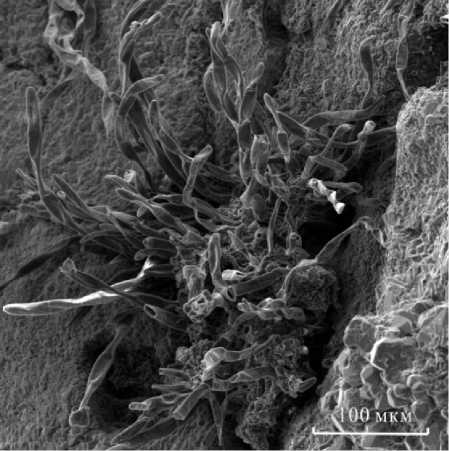
Рис. 5. Чехлы современных цианофитов в трещинах кварцевой матрицы внутри створки раковины.
Fig. 5. Covers of modern cyanophytes in the cracks of the quartz matrix inside the shell sashe.
Вмещающая порода брахиоподовой банки визуально на срезе представляет собой агатоподобную структуру. Исследования структурными методами показали, что преимущественным минералом в породе является кварц (см. таблицу). Содержание SiO2 в породе по данным рентгенофлюоресцентного анализа составляет 79–87, СаО – 8-15 и Mg – 3-6 мас. %.
Изучение породы в прозрачных шлифах показало, что в постседиментацинный период сформированные кристаллы кварца, игольчатого халцедона, кварцина и агрегаты кальцита инкрустируют пустоты в структуре кварцевой матрицы. На РЭМ-изображениях кварц наблюдается в виде субмикрозернистых образований и индивидов без определенной ориентации, размер которых составляет 1–10 мкм. Зерна, как правило, в значительной степени протравлены по сравнению с кварцем, сформированным на наружной поверхности створки раковины, а отсутствие гранных форм и облик индивидов указывают на процессы растворения.
Вторичный доломит представлен как индивидами, так и агрегатами, часто формирующими инкорпорационную структуру. Подобно внутрирако-винным индивидам кристаллы доломита во вмещающей породе сильно корродированы и их распределение неравномерное.
Из трех модификаций карбоната кальция в породе структурными методами идентифицирован минерал – кальцит. В палеополостях окремененной породы на подложке первичного доломита он формирует сростки и монокристаллы скаленоэдриче-ского облика с субиндивидами на гранях. Кроме того, согласно микроснимкам карбонат кальция (Са 33 масс. %) в виде новообразованных веерообразных агрегатов визуализируется на поверхности кристаллов кальцитовых агрегатов (рис. 6, а, б).
На кристаллах кварца обнаружены карбонатсодержащие пленки чешуйчатой структуры с толщиной слоя 1 мкм, состоящие из упорядоченных палочковидных наноразмерных частиц (рис. 7). Состав нанокристаллов по данным микрозондового анализа представлен в мас.%: Са 30–33; Mg 2,5.

Рис. 6, а. Сросток вторичного кальцита в палеополостях вмещающей породы в брахиоподовой банке.
Fig. 6, a. Aggregates of secondary calcite in the paleocavities of the host rock in the brachiopod bank.

Рис. 6, б. Веерообразные агрегаты новообразованного карбоната кальция на поверхности кристалла вторичного кальцита.
Fig. 6, б. Fan-shaped aggregates of newly formed calcium carbonate on the surface of a secondary calcite crystal.
Обсуждение результатов
Гряда Чернышева расположена в северовосточной части Тимано-Печорского бассейна, где распространены деформированные силурийско-

Рис. 7. Наноиндивиды карбоната кальцита на поверхности кристаллов кварца из вмещающей породы. Fig. 7. Calcium carbonate nanoindivids on the surface of quartz crystals from the host rock.
Рентгенографические данные новообразованных минералов в псевдоморфозах X-ray data of newly formed minerals in pseudomorphs
|
d |
Стенка раковины брахиопода |
Внутрираковинные минералы |
Вмещающая порода в брахиоподовой банке |
|||||
|
1 |
2 |
1 |
2 |
3 |
1 |
2 |
3 |
|
|
hkl(I) |
hkl(I) |
hkl(I) |
hkl(I) |
hkl(I) |
hkl(I) |
hkl(I) |
hkl(I) |
|
|
4,26 |
100(16) |
― |
100(20) |
― |
― |
100(30) |
― |
― |
|
3,34 |
101 100) |
― |
101(100) |
― |
― |
101(100) |
― |
― |
|
3,03 |
― |
― |
― |
― |
104 (4) |
― |
― |
104(2) |
|
2,89 |
― |
104(3) |
― |
104 37) |
― |
― |
104(64) |
― |
|
2,67 |
― |
― |
― |
006 (2) |
― |
― |
006(1) |
― |
|
2,54 |
― |
― |
― |
015(2) |
― |
― |
― |
― |
|
2,46 |
110 (8) |
― |
110(9) |
― |
― |
110(12) |
― |
― |
|
2,40 |
― |
― |
― |
110(1) |
― |
― |
― |
― |
|
2,29 |
102 (7) |
― |
102(9) |
― |
― |
102(9) |
― |
― |
|
2,23 |
111 (3) |
― |
111(4) |
― |
― |
111(5) |
― |
― |
|
2,19 |
― |
― |
― |
113(30) |
― |
― |
113(2) |
― |
|
2,13 |
200(5) |
― |
200(5) |
― |
― |
200(8) |
― |
― |
|
2,02 |
― |
― |
― |
202(2) |
― |
― |
202(1) |
― |
|
1,979 |
201 (4) |
― |
201(4) |
― |
― |
201(5) |
― |
― |
|
1,817 |
112 (13) |
― |
112(15) |
― |
― |
112(17) |
― |
― |
|
1,803 |
― |
― |
― |
018(5) |
― |
― |
018(3) |
― |
|
1,785 |
― |
― |
― |
116(3) |
― |
― |
116(2) |
― |
|
1,670 |
202(5) |
― |
202(5) |
― |
― |
202(5) |
― |
― |
|
1,542 |
― |
― |
211(6) |
― |
― |
211(11) |
― |
― |
Примечание: 1 – кварц; 2 – доломит; 3 – кальцит. (I) – относительная интенсивность дифракционных пиков; прочерк – дифракционные пики отсутствуют.
нижнепермские преимущественно карбонатные отложения с подчиненными прослоями песчаноглинистых пород и эвапоритов позднего ордовика [5]. Один из наиболее представительных по полноте разрезов силура находится в центральной части гряды Чернышева на р. Шаръю. Он расположен в районе «нижних ворот» среднего течения реки. Возраст самой древней толщи в этом разрезе – нижне-среднелландоверийский – предположительно определяется по находкам брахиопод Borealis sр., представленных массовым скоплением (ракуш-няковой банкой).
На развитие брахиопод и других представителей бентосных организмов, населявших Тимано-Североуральский бассейн в раннем силуре, влияли специфика тектонического режима территории, эв-статические изменения уровня Мирового океана, седиментологические условия, климатические изменения и другие [1]. Глобальная силурийская трансгрессия с начала лландовери способствовала широкому расселению брахиопод. Усиление регрессивных тенденций на большей части Североуральского палеобассейна в позднеаэронское время, вымирание бентосной биоты, в том числе и брахиопод Borealis, за исключением строматопорат и цианобактериальных сообществ, свидетельствуют о крупной экосистемной перестройке на рубеже аэрона и телича. В разрезе этот рубеж литологически фиксируется появлением тонкоплитчатых строматолитовых известняков в основании телича (фи-липпъельского горизонта) [1, 6–8].
Ранее были показаны особенности современного биоминералообразования в гликокаликсе цианопрокариот из различных водных сред [9]. Определенные современные виды цианофитов, продуцируя слизистые образования, обрастают к субстрату. Формирование микроминералов независимо от условий кристаллизации и места обитания цианопрокариот происходит в пределах их гликокаликса.
Первоначальное формирование внеклеточных зерен кварца при участии цианобактериальных симбионтов (губок, радиолярий), на наш взгляд, могло быть в пределах бактериальной биопленки на поверхности створок, возможно, еще живых форм брахиопод. В качестве источника кремния древние цианофиты использовали биогенный кремнезем – радиолярии и губки. Однако остатки организмов, служивших источником кремнекислоты, не считая единичных спикул, замещенных кремнеземом, нами не обнаружены в петрографических шлифах.
Изначально в доломитизированных брахио-подах на стадии диагенеза, в последующем, на наш взгляд, произошло первичное окременение. Повышенное содержание растворенного кремнезема может вызвать растворение доломита и окремене-ние раковин в определенных литологических условиях. После осаждения кремнезем проходит стадии постседиментационных трансформаций: опал → скрытокристаллический кварц или халцедон → суб-микрозернистый кварц.
Полагаем, что ячеистая структура агрегатов новообразованного кварца на наружной поверхности стенки створок брахиопод сформировалась в гликокаликсе современных цианофитов. Присутствие реликтов окремененного гликокаликса, выявленного на кристаллах и между агрегатами, а также полимерного органического вещества и частично минерализованных чехлов современных цианофи-тов, указывает на участие биоты в формировании псевдоморфоз кварца по створкам раковин раннесилурийских брахиопод.
Заключение
Таким образом, показано, что в постседиментационных условиях изменились минеральный состав и структура стенки раковин брахиопод. Окре-менение стенки раковин брахиопод привело к полной замене первоначального карбонатного скелета с изменением его структурной организации.
Наблюдаемая структурная упорядоченность, размеры кристаллов на стенках створок брахиопод, различающаяся не только на разных ископаемых образцах, но и в пределах одной створки раковины, свидетельствует о присутствии кварца разных генераций. На основе изученных микроснимков можно сказать, что кварц на створках раковин и во внутри-раковинном веществе брахиопод претерпел неоднократные преобразования, обусловленные совокупностью внешних факторов: процессов древнего и современного выветривания.
Реликты биоминеральных пленок на кварцевых щетках свидетельствуют о влиянии микробиальных сообществ на процессы замещения первичного карбонатного скелета на первичный кварцевый.
Генезис кристаллических агрегатов кварца на поверхности связан как с современными, так и раннепалеозойскими процессами биоминералообразования при участии цианобактериальных симбионтов (радиолярий, губки).
Авторы благодарят д.г.-м.н., гл.н.с. В.И. Ракина, Е.М. Тропникова за сотрудничество в исследованиях.
Список литературы Псевдоморфообразование по раковинам силурийских брахиопод
- Безносова Т.М. История развития пентаме-рид (Brachiopoda) в Тимано-Североуральском бассейне // Палеонтологический журнал. 2014. № 1. С. 49–54.
- Сапельников В.П. Морфология и таксономи-ческая эволюция брахиопод (отряд пентаме-риды). Свердловск, 1985. 231с.
- Ушатинская Г.Т., Малаховская Я.Е. Первые брахиоподы с карбонатным скелетом: появ-
- ление, расселение, строение стенки ракови-ны // Эволюция биосферы и биоразнообра-зия. М.: ПИН РАН, 2006. С. 177–192.
- Мадисон А.А. Микроструктура раковины строфоменид рода Bilovia Cooper (brachi-opoda) из позднего ордовика Балтоскандии // Палеонтологический журнал. 2019. № 1. С. 34–45.
- Соборнов К.О., Данилов В.Н. Гряда Черны-шева: суперловушка или зона рассеивания углеводородов? // SPE : Материалы конфе-ренции. М., 2015. С. 1–18.
- Безносова Т.М. Сообщества брахиопод и био-стратиграфия ордовика, силура и нижнего девона северо-восточной окраины палеокон-тинента Балтия. Екатеринбург: УрО РАН, 2008. 216 с.
- Цыганко В.С., Безносова Т.М., Лукин В.Ю., Соболев Д.Б. Палеозойская стратисфера севе-ро-востока Европы: стратиграфическая кор-реляция, биогеособытийная история, эволю-ция морских палеоэкосистем // Известия Коми НЦ УрО РАН. 2010. № 3 (3). С. 55–61.
- Матвеев В.А. Основные морфотипы и мик-роструктура лландоверийских строматолито-вых построек на западном склоне Припо-лярного Урала // Вестник ИГ Коми НЦ УрО РАН. 2013. № 2. С. 17–20.
- Каткова В. И., Митюшева Т. П., Филиппов В. Н., Симакова Ю.С. Минеральные включе-ния в цианобактериях из водных объектов юга Республики Коми // Вестник Кольского научного центра РАН. 2017. № 4. С. 65–71.
