Психофизиологическая методика изучения нарушений процесса прогностического кодирования

Автор: Телешева К.Ю., Рабинович Э.И.
Журнал: Психология. Психофизиология @jpps-susu
Рубрика: Психофизиология
Статья в выпуске: 3 т.17, 2024 года.
Бесплатный доступ
Обоснование. Теория прогностического кодирования является моделью функционирования мозга, описывающей его как орган, непрерывно генерирующий гипотезы на основе прошлого опыта, сверяющий их с полученной информацией, корректирующий и оптимизирующий прогнозы. Теория является перспективной основой для объяснения многих психопатологических механизмов, однако экспериментальные подходы к изучению процессов прогностического кодирования недостаточно разработаны. В статье описывается авторская психофизиологическая методика, направленная на изучение особенностей процесса прогностического кодирования в условиях предъявления визуальных стимулов с вариативной вероятностью. Приведены сценарии стимуляции, регистрации, обработки и интерпретации полученных данных. Описан дизайн эксперимента, приведены предварительные результаты пилотного исследования.
Прогностическое кодирование, электроэнцефалография, p300, условная негативная волна, саккадические движения глаз, шизофрения
Короткий адрес: https://sciup.org/147246082
IDR: 147246082 | УДК: 159.91, | DOI: 10.14529/jpps240310
Developing a psychophysiological method to examine violations of predictive coding processes
Introduction. The predictive coding theory posits that the brain functions as a “prediction machine”, continuously generating hypotheses based on past experiences and comparing them against actual input signals. This theory is a promising basis for explaining many psychopathological mechanisms. However, experimental approaches to investigate predictive coding processes have not been sufficiently explored. The article introduces a novel psychophysiological method designed to investigate the characteristics of predictive coding processes during variable visual stimulation. The paper outlines the experimental protocol, detailing the scenarios for stimulation, data acquisition, processing, and interpretation. The paper describes the experimental design and presents preliminary results from a pilot study.
Текст научной статьи Психофизиологическая методика изучения нарушений процесса прогностического кодирования
Теория прогностического кодирования предполагает, что мозг не пассивно обрабатывает поступающую информацию, а использует прошлый опыт для формирования прогнозов о том, что, вероятно, произойдет в будущем. Эти прогнозы позволяют интерпретировать зашумленные или двусмысленные данные, принимать решения, несмотря на неопределенность, и действовать эффективно и проактивно, не ожидая сенсорной обратной связи [1].
Нарушение процессов прогностического кодирования может являться основой психопатологической симптоматики при ряде психических расстройств: шизофрении [2], РАС [3], депрессии [4].
При большом количестве теоретических концепций и эмпирических исследований в русле теории отмечается дефицит методик изучения психофизиологических механизмов влияния вероятности на построение прогноза и формирование сигнала ошибки предсказания в зрительной модальности. Предлагаемые в статье способы стимуляции и обработки полученных записей ЭЭГ позволят расширить представления о топографии различных этапов процессов прогностического кодирования, выявлять нарушения на этапе предна-стройки, детекции ошибки предсказания и принятия решения, уточнить выявленные особенности нарушения прогностического кодирования у лиц с шизофренией за счет информации о влиянии вариации вероятности предъявления стимулов.
Обзор литературы
Теория прогностического кодирования получила активное развитие в связи с работами над усовершенствованием искусственного интеллекта, однако основные постулаты модели были сформулированы ранее. Так, в концепции бессознательного вывода Г. Гельмгольца предполагается, что в процессе зрительного восприятия мозг дополняет визуальную информацию на основе предыдущего опыта, чтобы придать ей смысл [5].
Когнитивные функции зависят от способности организма использовать прошлый опыт и делать выводы о будущих состояниях [6]. Прогностическое кодирование объясняет некоторые особенности внимания, восприятия, социального познания [1], принятия решений, контроля движений [7]. Предварительные ожидания возникают из нескольких источников информации, где хранятся соответствующие предварительные знания, и вовлекают различные зоны мозга [8], что определяет вовлечение широких нейронных сетей в процессы прогностического кодирования.
Значительную часть исследований в русле теории прогностического кодирования составляют исследования предсказания результата движения. Известно, что в этих процессах участвуют области головного мозга, ответственные за наблюдение за действиями, – системы зеркальных нейронов, сети наблюдения за действиями [9, 10]. Предполагается, что программирование собственных и понимание действий других происходит за счет моделирования наблюдаемых действий и их последствий [11]. Широко изучаются особенности процессов прогнозирования у спортсменов [12–14], танцоров [15, 16], водителей [17]. Также большое количество исследований в рамках теории прогностического кодирования сосредоточено на поиске механизмов принятия решений на моделях ожидания и предсказания вознаграждения [18, 19].
Процессы прогностического кодирования наиболее изучены в зрительной модальности, в связи с чем в текущем исследовании была выбрана визуальная стимуляция на основе парадигмы пространственной сигнализации М. Познера cost-benefit для изучения зрительного внимания, в которой предлагается два условия вероятности: 80 и 50 % [20].
Оценка влияния изменения вероятности на активность мозга была реализована в нескольких исследованиях. В контексте предъявления эмоциональных зрительных стимулов (фотографии лиц, 50, 75 и 100 % совпадения валентности без сообщения испытуемым об условиях соответствия) выявлено, что амплитуда условной негативной волны (contingent negative variation – CNV) максимальна в условиях умеренной вероятности (75 %), что расценивается как экономия ресурсов в полностью прогнозируемых условиях (100 %) [21]. В зрительнослуховой модификации задачи Познера при варьировании вероятности 50, 64 и 88 % были выявлены более быстрые и точные моторные ответы, большая амплитуда CNV, увеличение амплитуды Р300 на девиантный стимул в условиях более высокой вероятности [22].
Привлечение внимания для оптимизации сенсорной точности объясняет как нейронные, так и поведенческие эффекты, наблюдаемые в зрительно-пространственных задачах Познера [23]. Внимание модулирует реакцию определенной популяции нейронов на входные сигналы и зависит от условных ожиданий от более высоких иерархических уровней [24]. Предшествующие ожидания могут моделировать сенсорную обработку как на ранних, так и на поздних стадиях, как до, так и после появления стимула [8], в связи с чем для анализа были выбраны различные временные периоды и компоненты вызванных потенциалов.
Нейрофизиологическим показателем, отражающим изменения в ожиданиях (построение прогноза, сигнал «сверху вниз»), является CNV – нарастание негативности между двумя взаимосвязанными стимулами (предупреждающим и императивным) в процессе выполнения задач, предполагающих моторный ответ [25].
Показателем ошибки прогнозирования «снизу вверх» является негативность рассогласования (mismatch negativity, MMN) [26] – сиг- нал несоответствия между входящими данными и репрезентациями памяти. MMN, независимый от внимания, считается локальным эффектом кратковременного отклонения стимула, ослабляет прогнозируемую ошибку на нижнем уровне, но не может предсказать отклонения, возникающие в более длительных временных масштабах. Ошибка прогнозирования на более высоком уровне индексируется P300 и определяется как глобальный эффект долгосрочного отклонения стимула [27]. Таким образом, MMN и Р300 являются различными показателями идентификации ошибки прогнозирования, однако Р300 отражает процесс корректировки прогноза на более высоком уровне. Компоненты CNV и Р300 отражают предшествующее ожидание и несоответствие прогноза, что было показано в зрительно-слуховой модификации теста Познера [22].
Целью исследования является разработка методики выявления психофизиологических особенностей процесса прогностического кодирования и ее апробация на психически здоровых лицах и лицах с шизофренией.
Материалы и методы
В качестве материала исследования были выбраны лица с шизофренией, для которых доказано нарушение процессов прогностического кодирования [28, 29].
В пилотном исследовании приняли участие 22 человека: в экспериментальную группу вошли 12 человек с диагнозом параноидная шизофрения (F21.0, средний возраст –
33,0 ± 11,2 года), контрольную группу составили 10 человек без психических заболеваний, неврологических патологий, черепномозговых травм и нейроинфекций по результатам самоотчета (возраст – 28,0 ± 5,7 года). Все участники мужского пола с нормальным или скорректированным зрением, ведущей правой рукой и правым глазом. Все участники в полной мере проинформированы о целях исследования и подписали добровольное информированное согласие.
Для стимуляции использовался стимулятор STIM2 (Compumedics Neuroscan, США), зрительные стимулы предъявлялись на мониторе, находящемся в затемненном помещении на расстоянии 60 см от глаз испытуемых. Испытуемые выполняли саккадическую задачу в парадигме пространственной сигнализации: каждая реализация включала в себя четыре последовательно предъявляемых типа стимулов: 1) установочный стимул зелено-го/желтого цвета, появляющийся на 200 мс в центре экрана; 2) центральный фиксационный стимул белого цвета, появляющийся через 600–800 мс на месте установочного стимула на 900–1100 мс; 3) сигнальный стимул белого цвета (стимул-«подсказка», предупреждающий), появляющийся сразу после центрального фиксационного стимула на 5 см левее или правее последнего на 150 мс; 4) периферический стимул зеленого цвета (целевой, императивный стимул), появляющийся через 1300– 1500 мс в 1 см от края монитора на 1000 мс (рис. 1).
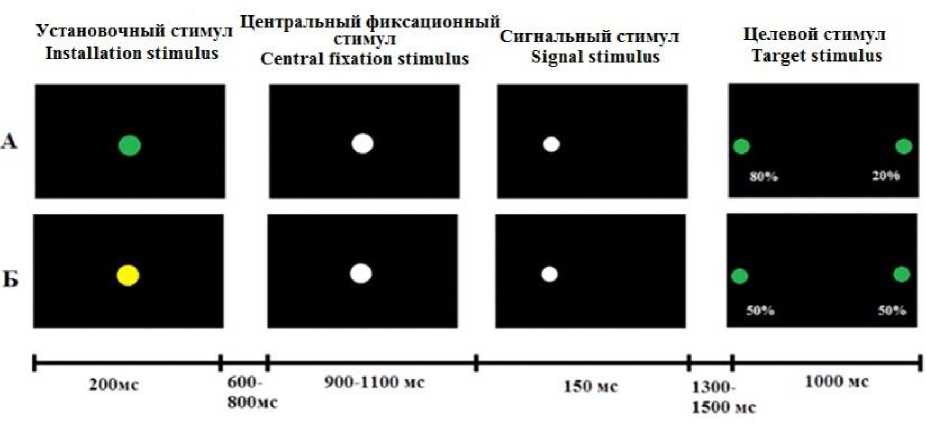
Рис. 1. Схема предъявления зрительных стимулов: А – условие, в котором целевой стимул появляется в том же полуполе зрения, что и сигнальный в 80 % случаев;
Б – периферический стимул в том же полуполе в 50 % случаев
Fig. 1. The scheme of visual stimuli: A – the target stimulus appears in the same half-field of vision as the signal in 80 % of cases; Б – 50 % of cases
Средние значения латентного периода саккад в мс (медиана, стандартная ошибка, значимость отличий по критерию Манна – Уитни)
The average latent period of saccades in ms (median, standard error, significance of differences –Mann–Whitney U-test)
|
Вариант стимула / Incentive option |
Контроль Control |
Шизофрения Schizophrenia |
P U-test |
|
|
80 % |
Стандартный / Standard |
206,0 ± 12,7 |
227,0 ± 12,0 |
0,251 |
|
Девиантный / Deviant |
225,0 ± 15,6 |
247,0 ± 15,3 |
0,349 |
|
|
50 % |
Стандартный / Standard |
222,0 ± 14,2 |
230,0 ± 16,5 |
0,654 |
|
Девиантный / Deviant |
219, ± 15,8 |
225,0 ± 15,2 |
0,705 |
|
В исследовании с равной вероятностью использовались две экспериментальные схемы: периферический стимул с 80%-ной вероятностью предъявляется с той же стороны, что и сигнальный стимул (установочный стимул зеленого цвета); вероятность совпадения стороны предъявления сигнального и периферического стимулов равняется 50 % (установочный стимул желтого цвета). Совпадение предъявления сторон в обеих схемах – стандартный стимул, несовпадение – девиантный.
Участники были проинструктированы при предъявлении первых трех стимулов фиксировать взгляд в центре экрана, при предъявлении целевого стимула как можно быстрее переводить на него взгляд. Каждая реализация начиналась с нажатия испытуемым на кнопку. Эксперимент состоял из 5 блоков, каждый из которых содержал 45 реализаций.
Регистрация осуществлялась на электроэнцефалографе Neuroscan Synamps System (Compumedics, США), ЭОГ регистрировалась с помощью Ag-AgCl кожных электродов, расположенных у латеральных углов обоих глаз, ЭЭГ – от 19 отведений, установленных по стандартной системе «10–20» в монополярном монтаже с референтными электродами на мочках ушей. Частота дискретизации сигнала ЭЭГ составляла 1000 Гц. Для анализа записи фильтровались в диапазоне 0–30 Гц. Удаление глазодвигательных артефактов производилось с помощью анализа независимых компонент (ICA). После этого записи мануально проверялись на наличие артефактов. Для анализа Р 300 записи фильтровались в полосе 1–30 Гц.
Статистические методы: U-критерий Манна – Уитни (анализ латентного периода саккад между группами), критерий знаковых рангов Вилкоксона (анализ латентного периода саккад внутри групп), дисперсионный анализ с повторными измерениями (rANOVA, выявление различий в показателях ЭЭГ), анализ средних значений (T-test, анализ различий), апостериорный анализ с использованием поправки Тьюки (анализ различий). Расчеты выполнены с использованием статистического SPSS Statistics 22.
Анализ ЭЭГ. Для CNV усреднялись эпохи от –1 до 0 с до целевого стимула с выравниванием базовой линии по периоду от –1 до –0,9 с. Средние значения амплитуды CNV были рассчитаны для двух периодов в соответствии с динамикой медленных потенциалов – I интервал соответствует раннему подготовительному периоду (0,9 – 0,7 с), II интервал – позднему (0,2 – 0 мс до стимула) периоду CNV [30]. Для Р 300 усреднение проводилось в периоде от –0,2 до 0,7 с от момента подачи периферического (целевого) стимула с выравниванием базовой линии по предсти-мульному интервалу от –0,2 до 0 с.
Для дисперсионного анализа были выбраны 9 каналов (F3, F4, Fz, C3, C4, Cz, P3, P4, Pz), наименее подверженные глазодвигательным, миографическим артефактам, артефактам моргания, позволяющие оценить ла-терализацию и локализацию активности. Выбраны межсубъектный фактор «Группа» (контроль, шизофрения) и внутрисубъектный фактор «Вероятность» (50 % и 80 % вероятности совпадения). Помимо перечисленных для анализа CNV выбраны факторы «Период» (I и II), «Полушарие» (левое и правое), «Сторона» предъявления стимула (левая и правая), «Локус» (n = 8: Fp; F; C; P; O; F; T; T). Post-hoc проводился анализ средних значений (T-test). Для MMN и Р 300 также выбраны факторы «Совпадение» (совпадение сигнального и целевого стимулов: стандартный, девиантный), «Расположение» электродов (фронтальное, центральные, теменное) и «Латеральность» электродов (n = 3: левое, центральное, правое расположение). Апостериорный анализ проведен с использованием поправки на множественные сравнения Тьюки.
Были рассчитаны средние значения латентного периода регулярных саккад (с латентностью более 160 мс от момента предъяв- ления стимула) по окулограмме для каждого из условий предъявления целевого стимула, количество экспресс-саккад (с латентным периодом менее 160 мс, возникающих после появления цели) и антиципаторных саккад (латентный период менее 160 мс, возникающих раньше появления целевого стимула) [31, 32]. Для сравнения латентности внутри групп в разных условиях был использован критерий Вилкоксона.
Результаты
Саккады . Различий между группами по латентному периоду регулярных саккад не выявлено (см. таблицу), однако отмечается специфика изменения латентного периода при изменении вероятности в группах. В группе контроля наблюдалось уменьшение латентности регулярных саккад при вероятности совпадения 80 % на стандартный стимул (совпадение) по сравнению с девиантным (несовпадение) (p = 0,004). Также выявлено увеличение процента экспресс- и антиципаторных саккад в условии совпадения стимулов при вероятности 80 % по сравнению с условием 50 % (p = 0,048). В группе лиц с шизофренией значимых отличий не выявлено, процент экспресс и антиципаторных саккад (с латентностью до 160 мс) в условии совпадения при вероятности 80 % по сравнению с 50 % не увеличивается (p = 0,195), общее количество экспресс-саккад больше.
CNV. Результаты проведенного дисперсионного анализа показали, что в обеих группах амплитуда CNV существенно возрастала от I (–900…–700 мс до стимула) ко II интервалу анализа (–200…0 мс до стимула, рис. 2).
В группе контроля в условии 50 % вероятности совпадения при предъявлении сигнального стимула слева зона максимальной негативности в I интервале (до 1,19 мкВ) включала медиальную и правую теменную и задневисочную области (см. рис. 1). Во II интервале амплитуда CNV достоверно возросла (p < 0,05), главным образом в лобно-центральной и в теменной области. Во II интервале зона максимальной негативности (до 5,1 мкВ) локализовалась в центрально-теменной области.
В группе больных шизофренией максимум CNV в I интервале (0,67 мкВ) зарегистрирован в отведении F8, во II интервале амплитуда CNV увеличилась, однако в меньшей степени (значимо в F3), а максимальное значение амплитуды составило 1,45 мкВ (в отве- дении T4). Таким образом, зона максимальной негативности характеризовалась низкой амплитудой и не имела отчетливой локализации.
Значимые различия по результатам дисперсионного анализа между группами зарегистрированы во II интервале в условиях стимуляции 50 % слева и 80 % справа по взаимодействию факторов «Область» × «Полушарие» (F = 8,60; p = 0,010), сравнение средних показало, что в группе больных амплитуда CNV была ниже в прецентральных и центральных областях коры (p < 0,05).
Вероятность совпадения сигнального и целевого стимулов также оказывает влияние на амплитуду CNV. В группе лиц с шизофренией были обнаружены различия между условиями 50 % и 80 % совпадения во II интервале. При сравнении условий предъявления стимулов были значимы фактор «Стимул» (F = 4,09, p = 0,078), взаимодействие «Стимул» × «Область» (F = 2,05, p = 0,064). Сравнение средних значений показало, что при 80 % совпадения амплитуда CNV выше в отведениях F3, Pz (p < 0,05).
Р300 . При оценке амплитуды компонента P 300 дисперсионный анализ показал значимое влияние группы при взаимодействии факторов «Совпадение» × «Сторона» × «Вероятность» (F = 5,5607, p = 0,029), факторов «Расположение» × «Вероятность» (F = 4,7335, p = 0,025), «Латеральность» × «Совпадение» × «Вероятность» (F = 3,9196, p = 0,032).
Апостериорный анализ с поправкой Тьюки показал, что значимых различий по амплитуде P 300 при совпадении и несовпадении стимулов в условии 50 % нет. При вероятности совпадения 80 % в группе контроля наблюдается значимое увеличение амплитуды P 300 при несовпадении сторон (t = 5,802, Ptukey = 0,001). Также в группе контроля наибольшее увеличение амплитуды компонента P300 при несовпадении сторон стимулов при вероятности 80 % наблюдается над центральными отведениями (риc. 3). В группе пациентов с шизофренией увеличение амплитуды P 300 наблюдается при несовпадении сторон стимулов при вероятности, равной 50 % над центрально-теменными отведениями, максимальные значения амплитуды наблюдаются над правым полушарием и по средней линии.
Обсуждение
В литературе на сегодняшний день не описаны методики, использующие модуля-
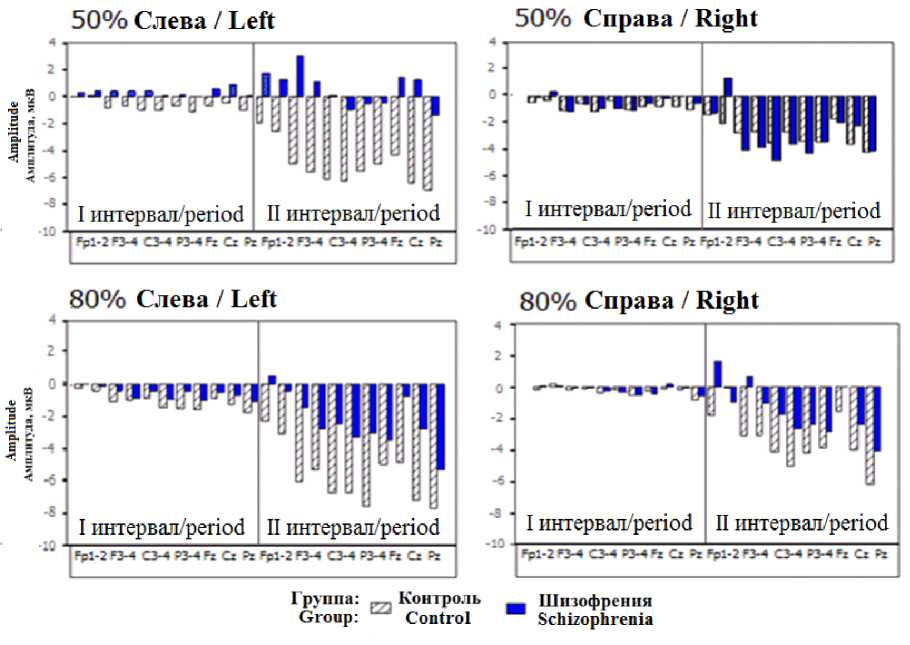
Рис. 2. Средние значения амплитуды CNV в I и II интервалах анализа: 50 % – 50% вероятность совпадения; 80 % – 80 % вероятность совпадения; Слева – сигнальный стимул предъявляется в левом полуполе;
Справа – сигнальный стимул предъявляется в правом полуполе
Fig. 2. Average values of the CNV amplitude in the I and II analysis period: 50 % – 50 % probability of coincidence; 80 % – 80 % probability of coincidence; Left – the signal stimulus is presented in the left half–field;
Right – the signal stimulus is presented in the right half-field
цию вероятности в парадигме вероятностного предъявления в зрительной модальности для оценки нарушений этапов прогностического кодирования и влияния изменения вероятности (изменение значимости стимула) на построение прогнозов. Авторами была разработана и апробирована методика стимуляции на основе зрительной задачи Познера, включающая установочный стимул, задающий вероятность совпадения (50 и 80 %), сигнальный стимул, указывающий сторону предъявления и целевой стимул, требующий моторного ответа (саккадического движения). Данное сочетание стимулов позволяет проанализировать этапы построения прогноза, сличения, детекции и сигнала ошибки, корректировки прогноза.
И хотя анализ результатов пилотного эксперимента выходит за рамки целей данной статьи, представляется необходимым кратко резюмировать выявленные различия. Психически здоровые лица отличаются устойчивой локализацией зоны максимальной негативности в периоде 200 мс до целевого стимула, существенным увеличением амплитуды CNV в период ожидания целевого стимула, а также более высокой амплитудой CNV в условиях 80%-ной вероятности совпадения стимулов по сравнению с 50 %, что отражает усиление процессов прогностического кодирования при увеличении значимости стимула. В группе больных с шизофренией амплитуда и локализация зон максимальной негативности характеризовались высокой вариативностью, также более высокие значения CNV выявлены при вероятности 80 %, наиболее низкие наблюдались в условиях предъявления сигнального стимула слева с 50%-ной вероятностью совпадения стимулов.
Увеличение амплитуды CNV в условии 80 % в группе психически здоровых участников и распределение максимальной негативности от теменной области на раннем этапе к центрально-теменной области на позднем этапе CNV соответствует литературным данным о генерации CNV в височно-теменных, сенсомоторных и моторных областях [33].
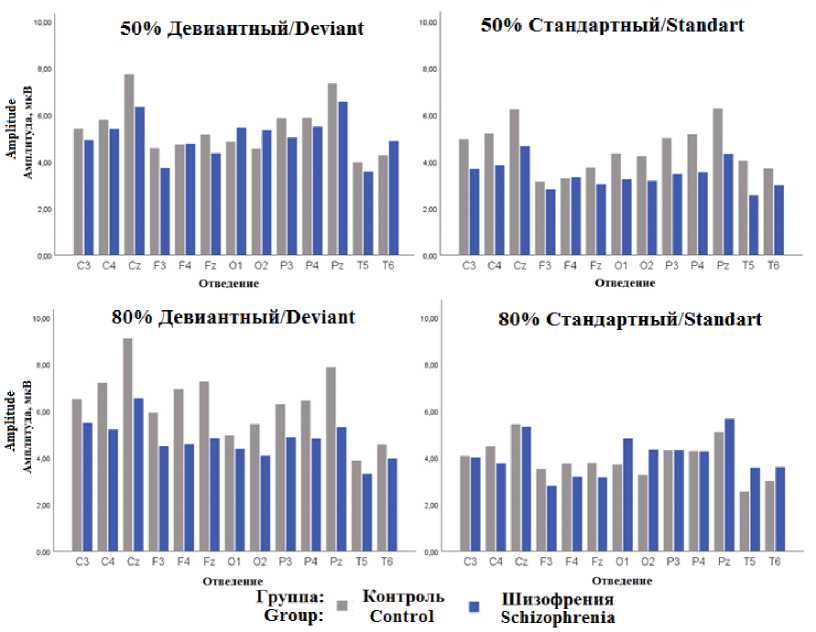
Рис. 3. Средние значения амплитуды Р 300 : 50 % – 50 % вероятность совпадения, 80 % – 80 % вероятность совпадения; Стандартный – сторона предъявления целевого и сигнального стимулов совпадает, Девиантный – сторона не совпадает
Fig. 3. Average values of the P 300 amplitude: 50 % – 50 % probability of coincidence, 80 % – 80 % probability of coincidence; Standard – the side of presentation of the target and signal stimuli coincides, Deviant – the side does not coincide
В течение периода ожидания наблюдалось градуальное нарастание амплитуды CNV, в большей степени выраженное в лобноцентральных и теменных зонах коры, что может отражать аспекты прогностического кодирования, связанные со зрительно-пространственным вниманием, включение корковой фронто-париетальной системы внимания и фронто-медио-таламической системы [34]. Показано изменение медленно-частотной активности в задней сенсомоторной, премоторной и медиальной лобной областях при ошибочных прогнозах, возникающих до выработки ответа, - снижение коэффициента фазовой синхронизации на фоне усиления синхронизации во фронтально-центральных областях [35].
В группе больных с шизофренией амплитуда CNV была ниже, чем в группе контроля, а значимые различия по амплитуде при сравнении в идентичных условиях получены для прецентральных зон коры. О снижении амплитуды CNV при шизофрении в лобно-центральных отделах коры неоднократно сообщалось в ис- следованиях других авторов [36]. Это соотносится с представлениями о «гипофронтальности» при шизофрении, согласно которым дисфункция префронтальной коры является ключевым фактором в патогенезе шизофрении, с которым связывают нарушения мышления и негативную симптоматику [37, 38].
Также в группе контроля наблюдалось достоверное увеличение амплитуды компонента Р300 при несовпадении стороны предъявления стимулов в условии вероятности их совпадения, равной 80 %, что соотносится с большим нарастанием CNV в данных условиях. Нарастание нисходящих ожиданий, приводящее к усилению внимания, является одним из условий, благодаря которому ошибки предсказания распространяются на уровень сознательного доступа [39]. Считается, что обработка информации на этом уровне объединяет информацию из распределенных лобно-теменных сетей, обеспечивая сознательный доступ к обнаруженным в них паттернам и отклонениям [40].
Использованная в исследовании схема стимуляции с различными уровнями вероятности позволила выявить особенности ранних этапов, которые отражаются в параметрах условного негативного отклонения – построения прогноза, предшествующего появлению целевого стимула, и дальнейшего сопоставления прогноза с сенсорными данными и сигнала ошибки прогнозирования, интенсивность которых отражается в изменении амплитуды компонента Р300.
Следует отметить некоторые ограничения исследования. В пилотном исследовании не были проанализированы некоторые показатели, согласно данным литературы также отражающие процессы прогностического кодирования. Распространенным биомаркером является феномен MMN [41, 42], который отражает нисходящую модуляцию [43]. Актуальным является поиск индикаторов мониторинга производительности (performance monitoring), который является одной из исполнительных функций – способности распознать ошибку вситуациях, когда происходит отклонение от цели [44]. Полагают, что неправильный мониторинг производительности является ключевым симптомом некоторых психических расстройств [45]. Биомаркером, индексирующим обнаружение ошибок, является негативность, связанная с ошибками (ERN) в медиальной лобной коре [46]. Предполагается дальнейшее расширение анализируемых показателей с исследованием феноменов MMN и ERN.
Также ограничением исследования является то, что инструкция предполагает достаточную сохранность когнитивных функций, что ограничивает целевую группу методики, так как когнитивное снижение является одним из симптомов как шизофрении, так и некоторых других психических заболеваний. Небольшой размер выборки может влиять на надежность полученных результатов, однако формат пилотного исследования предполагал апробацию методики и поиск нейрофизиологических коррелятов нарушения процессов прогностического кодирования. В дальнейшем предполагается расширение выборки и увеличение нозологических форм для углуб- ления понимания возможных механизмов нарушения процессов прогностического кодирования при различных психических заболеваниях и их роли в психопатологической симптоматике.
Заключение
В результате исследования были выявлены значимые различия между группой психически здоровых лиц и пациентами с шизофренией при выполнении задачи на вероятностное прогнозирование в условиях модуляции вероятности. Анализ условной негативной волны, отражающей нисходящие процессы ожидания стимула, выявил значимые различия в ее характеристиках между группами. В норме наблюдается волна с устойчивой фронто-теменной топографией, амплитуда которой увеличивается при увеличении вероятности совпадения стимула (увеличении его значимости). В группе больных с шизофренией амплитуда и локализация условной негативной волны характеризовалась высокой вариативностью и зависимостью от стороны предъявления стимула.
Различия в компоненте P 300 соответствуют различиям в условной негативной волне. Наибольшая амплитуда в группе нормы наблюдается при несоответствии сигнального и периферического стимула при вероятности их совпадения, равной 80 %. В группе пациентов с шизофренией увеличение амплитуды P300 наблюдается при несовпадении стимулов при вероятности, равной 50 %.
Результаты свидетельствуют о нарушении в группе пациентов с шизофренией процессов построения прогнозов, оценки вероятностей, распределения значимости между стимулами.
Целью исследования являлась разработка психофизиологической методики для выявления особенностей процесса прогностического кодирования и апробация методики на психически здоровых лицах и лицах с шизофренией. Полученные результаты исследования подтверждают способность разработанного метода количественно и качественно оценить указанные изменения при выбранном методе стимуляции и анализа полученных данных.
Список литературы Психофизиологическая методика изучения нарушений процесса прогностического кодирования
- D’Mello A.M., Bach P., Corlett P.R. et al. Editorial: Predictive mechanisms in action, percep-tion, cognition, and clinical disorders. Frontiers in Human Neuroscience. 2022;16:1005905. DOI: 10.3389/fnhum.2022.1005905.
- Wynn J.K., Green M.F. An EEG-Based Neuroplastic Approach to Predictive Coding in People With Schizophrenia or Traumatic Brain Injury. Clinical EEG Neuroscience. 2024;6:15500594241252897. DOI: 10.1177/15500594241252897.
- Sapey-Triomphe L.A., Sanchez G., Hénaff M.A. et al. Disentangling sensory precision and prior expectation of change in autism during tactile discrimination. NPJ Scientific Learning. 2023;8(1):54. DOI: 10.1038/s41539-023-00207-5.
- Spaeth A.M., Koenig S., Everaert J. et al. Are depressive symptoms linked to a reduced pupillary response to novel positive information? – An eye tracking proof-of-concept study. Frontiers in Psychology. 2024;23(15):1253045. DOI: 10.3389/fpsyg.2024.1253045.
- Tucker D.M., Luu P. Motive control of unconscious inference: The limbic base of adaptive Bayes. Neuroscience and Biobehavioral Reviews. 2021;128:328–345. DOI: 10.1016/j.neubiorev.2021.05.029.
- Hipólito I., Kirchhoff M. Breaking boundaries: The Bayesian Brain Hypothesis for perception and prediction. Consciousness and Cognition. 2023;111:103510. DOI: 10.1016/j.concog.2023.103510.
- Sprevak M., Smith R. An Introduction to Predictive Processing Models of Perception and Decision-Making. Topics in Cognitive Science. 2023;(28):1–28. DOI: 10.1111/tops.12704.
- de Lange F.P., Heilbron M., Kok P. How Do Expectations Shape Perception. Trends in Cognitive Science. 2018;22(9):764–779. DOI: 10.1016/j.tics.2018.06.002.
- Syrov N., Yakovlev L., Miroshnikov A. et al. Beyond passive observation: feedback anticipation and observation activate the mirror system in virtual finger movement control via P300-BCI. Frontiers in Human Neuroscience. 2023;4(17):1180056. DOI: 10.3389/fnhum.2023.1180056.
- Qin C., Michon F., Onuki Y. et al. Predictability alters information flow during action observation in human electrocorticographic activity. Cell Reports. 2023;42(11):113432. DOI: 10.1016/j.celrep.2023.113432.
- Scott M. Sensory attenuation from action observation. Experimental Brain Research. 2022;240(11):2923–2937. DOI: 10.1007/s00221-022-06460-1.
- Ashraf R., Abdoli B., Khosrowabadi R. et al. The Effect of Modeling Methods on Mirror Neuron Activity and a Motor Skill Acquisition and Retention. Basic Clinical Neuroscience. 2023;14(5):631–646. DOI: 10.32598/bcn.2021.3245.1.
- Ji Q., Liu L., Lu Y. et al. Mechanisms of Action Anticipation in Table Tennis Players: A Multivoxel Pattern Analysis Study. Neuroscience. 2024;16:33–40. DOI: 10.1016/j.neuroscience.2024.03.016.
- Murphy C.P., Runswick O.R., Gredin N.V. et al. The effect of task load, information reliability and interdependency on anticipation performance. Cognitive Research: Principles and Implications. 2024;9(1):22. DOI: 10.1186/s41235-024-00548-8.
- Amoruso L., Pusil S., García A.M. et al. Decoding motor expertise from fine-tuned oscillatory network organization. Human Brain Mapping. 2022;43(9):2817–2832. DOI: 10.1002/hbm.25818.
- Wang Y., Zhao Q., Ji Q. et al. fMRI evidence of movement familiarization effects on recognition memory in professional dancers. Cerebral Cortex. 2024;34(1):bhad490. DOI: 10.1093/cercor/bhad490.
- Ueda S., Sato T., Kumada T. Model-Based Prediction of Operation Consequences When Driving a Car to Compensate for a Partially Restricted Visual Field by A-Pillars. Frontiers in Human Neuroscience. 2021;15:697295. DOI: 10.3389/fnhum.2021.697295.
- Schwartenbeck P., FitzGerald T.H., Mathys C. et al. The Dopaminergic Midbrain Encodes the Expected Certainty about Desired Outcomes. Cerebral Cortex. 2015;25(10):3434–3445. DOI: 10.1093/cercor/bhu159.
- Chew B., Hauser T.U., Papoutsi M. et al. Endogenous fluctuations in the dopaminergic midbrain drive behavioral choice variability. Proceedings of the National Academy of Sciences of the USA. 2019;116(37):18732–18737. DOI: 10.1073/pnas.1900872116.
- Posner M.I. Orienting of attention: Then and now. Quarterly Journal of Experimental Psychology. 2016;69(10)1864–1875. DOI: 10.1080/17470218.2014.937446.
- Del Popolo Cristaldi F., Mento G., Sarlo M. et al. Dealing with uncertainty: A high-density EEG investigation on how intolerance of uncertainty affects emotional predictions. PLoS One. 2021;16(7):e0254045. DOI: 10.1371/journal.pone.0254045.
- Gómez C.M., Arjona A., Donnarumma F. et al. Tracking the Time Course of Bayesian Inference With Event-Related Potentials:A Study Using the Central Cue Posner Paradigm. Frontiers in Psychology. 2019;10:1424. DOI: 10.3389/fpsyg.2019.01424.
- Brown H.R., Friston K.J. The functional anatomy of attention: a DCM study. Frontiers in Human Neuroscience. 2013;7:784. DOI: 10.3389/fnhum.2013.00784.
- Auksztulewicz R., Friston K. Attentional Enhancement of Auditory Mismatch Responses: a DCM/MEG Study. Cerebral Cortex. 2015;25(11):4273–4283. DOI: 10.1093/cercor/bhu323.
- Sanquist T.F., Beatty J.T., Lindsley D.B. Slow potential shifts of human brain during forewarned reaction. Electroencephalography and Clinical Neurophysiology. 1981;51(6):639–649. DOI: 10.1016/0013-4694(81)90208-x
- Fitzgerald K., Todd J. Making Sense of Mismatch Negativity. Frontiers in Psychiatry. 2020;11:468. DOI: 10.3389/fpsyt.2020.00468.
- Chennu S., Noreika V., Gueorguiev D. et al. Expectation and attention in hierarchical auditory prediction. Journal of Neuroscience. 2013;33(27):11194–11205. DOI: 10.1523/JNEUROSCI.0114-13.2013.
- Нарушения обработки информации у больных шизофренией: обзор литературы / М.В. Иванов, М.А. Тумова, Л.М. Муслимова, Т.В. Капустина // Сибирский вестник психиатрии и наркологии. 2020. Vol. 2(107). P. 41–50. [Ivanov M.V., Tumova M.A., Kapustina T.V., Muslimova L.M. Information processing disorders in patients with schizophrenia: a literature review. Sibirskii vestnik psikhiatrii i narkologii = Siberian Herald of Psychiatry and Addiction Psychiatry. 2020;2(107):41–50. (in. Russ.)] DOI: 10.26617/1810-3111-2020-2(107)-41-50.
- Гусейнова З.Т., Тумова М.А., Шошина И.И. Особенности зрительной системы у больных параноидной шизофренией // Психиатрия. 2022. Т. 20, № S3(2). С. 12–14. [Guseinova Z.T., Tumova M.A., Shoshina I.I. Features of the visual system in patients with paranoid schizophrenia. Psikhiatriya = Psychiatry (Moscow). 2022;20(S3(2)):12–14. (in. Russ.)].
- Kirenskaya A.V., Myamlin V.V., Novototsky-Vlasov V.Y. et al. The contingent negative variation laterality and dynamics in antisaccade task in normal and unmedicated schizophrenic subjects. Spanish Journal of Psychology. 2011;14(2):869–883. DOI: 10.5209/rev_sjop.2011.v14.n2.34.
- Errington S.P., Schall J.D. Express saccades during a countermanding task. Journal of Neurophysiology. 2020;124(2):484–496. DOI: 10.1152/jn.00365.2020.
- Gouret F., Pfeuffer C.U. Anticipatory Saccades Towards the Future Consequences of One's Actions – an Online Eye Tracking Study. Journal of Cognition. 2023;6(1):15. DOI: 10.5334/joc.261.
- Thibault N., Albouy P., Grondin S. Distinct brain dynamics and networks for processing short and long auditory time intervals. Scientific Reports. 2023;13(1):22018. DOI: 10.1038/s41598-023-49562-8.
- Славуцкая М.В., Шульговский В.В., Семина Т.К. Влияние направленного внимания на потенциалы головного мозга человека при вероятностном предъявлении зрительных стимулов // Журнал высшей нервной деятельности им. И.П. Павлова. 2005. Т. 55, № 6. С. 796–805. [Slavutskaya M.V., Shulgovsky V.V., Semina T.K. Directed attention influence on the human brain potentials under conditions of probability visual stimulation. Zhurnal vysshei nervnoi deyatel'nosti im. I.P. Pavlova = I.P. Pavlov Journal of Higher Nervous Activity. 2005;55(6):796–805. (in. Russ.).]
- Yordanova J., Falkenstein M., Kolev V. Motor oscillations reveal new correlates of error processing in the human brain. Scientific Reports. 2024;14(1):5624. DOI: 10.1038/s41598-024-56223-x.
- Osborne K.J., Kraus B., Lam P.H. et al. Contingent Negative Variation Blunting and Psychomotor Dysfunction in Schizophrenia: A Systematic Review. Schizophrenia Bulletin. 2020;46(5):1144–1154. DOI: 10.1093/schbul/sbaa043
- Dharani R., Goyal N., Mukherjee A. et al. Adjuvant High-Definition Transcranial Direct Current Stimulation for Negative Symptoms in Schizophrenia: A Pilot Study. The Journal of ECT. 2021;37(3):195–201. DOI: 10.1097/YCT.0000000000000756.
- Kumar N., Vishnubhatla S., Wadhawan A.N. et al. A randomized, double blind, sham-controlled trial of repetitive transcranial magnetic stimulation (rTMS) in the treatment of negative symptoms in schizophrenia. Brain Stimulation. 2020;13(3):840–849. DOI: 10.1016/j.brs.2020.02.016.
- Vidal-Gran C., Sokoliuk R., Bowman H. et al. Strategic and Non-Strategic Semantic Expectations Hierarchically Modulate Neural Processing. eNeuro. 2020;7(5):ENEURO.0229-20.2020. DOI: 10.1523/ENEURO.0229-20.2020.
- Dehaene S., Changeux J.-P. Experimental and theoretical approaches to conscious processing. Neuron. 2011;70(2):200–227. DOI: 10.1016/j.neuron.2011.03.018
- Yasoda-Mohan A., Vanneste S. Development, Insults and Predisposing Factors of the Brain's Predictive Coding System to Chronic Perceptual Disorders-A Life-Course Examination. Brain Sciences. 2024;14(1):86. DOI: 10.3390/brainsci14010086.
- Tivadar R.I., Knight R.T., Tzovara A. Automatic Sensory Predictions: A Review of Predictive Mechanisms in the Brain and Their Link to Conscious Processing. Frontiers in Human Neuroscience. 2021;15:702520. DOI: 10.3389/fnhum.2021.702520.
- Sikkens T., Bosman C.A., Olcese U. The Role of Top-Down Modulation in Shaping Sensory Processing Across Brain States: Implications for Consciousness. Frontiers in Systems Neuroscience. 2019;13:31. DOI: 10.3389/fnsys.2019.00031.
- Fu Z., Sajad A., Errington S.P. et al. Neurophysiological mechanisms of error monitoring in human and non-human primates. Nature Reviews Neuroscience. 2023;24(3):153–172. DOI: 10.1038/s41583-022-00670-w.
- Gillan C.M., Fineberg N.A., Robbins T.W. A trans-diagnostic perspective on obsessive-compulsive disorder. Psychological Medicine. 2017;47(9):1528–1548. DOI: 10.1017/S0033291716002786.
- Sajad A., Godlove D.C., Schall J.D. Cortical microcircuitry of performance monitoring. Nature Neuroscience. 2019;22(2):265–274. DOI: 10.1038/s41593-018-0309-8.
