Пуринергическая регуляция синаптической передачи фазных и тонических мышц позвоночных

Автор: Гришин С.Н., Морозов О.Г., Анфиногентов В.И., Камалиев Р.Р., Морозов Г.А., Зиганшин А.У.
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Биотехнологии и биоинженерия
Статья в выпуске: 4-3 т.12, 2010 года.
Бесплатный доступ
С помощью компьютерного пакета программ «Mathcad Professional 2000» проводилось моделирование действия пуринов - АТФ и ее конечного производного аденозина на амплитуду постсинаптических ответов в нервно-мышечных переходах фазных и тонических мышц озерной лягушки. Известно, что АТФ, как и аденозин, по принципу отрицательной обратной связи угнетает свой квантовый выброс в синаптическую щель. В синапсах тонической мускулатуры АТФ, в отличие от сохраняющего свой эффект аденозина, вызывает повышение амплитуды постсинаптических ответов. Система кинетических уравнений решалась методом Рунге-Кутта с фиксированным шагом. Результирующие кривые математического моделирования в общем повторяют экспериментальные данные. Были получены конкретные значения коэффициентов полинома и значения равновесных констант.
Атф, аденозин, фазная и тоническая мышцы, математическое моделирование, атp
Короткий адрес: https://sciup.org/148199462
IDR: 148199462 | УДК: 629.782.519.711
Purinergical regulation of synaptic transfer in phase and tonic muscles of vertebrates
By means of the computer software package «Mathcad Professional 2000» modeling of purine action - АТP and its final derivative adenosine on amplitude of post-synaptic answers in nervously-muscular transitions of phase and tonic muscles in a lake frog was spent. It is known, that АТP, as well as adeno-sine, by a principle of negative feedback oppresses the quantum emission in synaptic gap. In synapses of tonic musculation АТP, unlike the adenosine keeping the effect, causes rising amplitude of post-synaptic answers. The system of the kinetic equations was solved by Runge-Kuttа method with the fixed step. Resulting curves of mathematical modeling in general repeat experimental data. Concrete value of poly-nom coefficients and value of equilibrium constants have been received.
Текст научной статьи Пуринергическая регуляция синаптической передачи фазных и тонических мышц позвоночных
А1 типа, напрямую сопряженных с калиевыми каналами А типа [5]. Эффект аденозина не опосредуется изменением активности Са-каналов, он не снижает амплитуду Са-токов. Ранее мы выявили активность Р 2 Y рецепторов АТФ на пресинапсе озерных лягушек, сопряженных через G i/o подтип ГТФ-белка с системой вторичных посредников, включающих: фосфолипазу С (расщепляет фосфолипиды мембраны), диацилглицерол, протеинкиназу С (встраивает фосфолипазу А2 из цитоплазмы в мембрану), фосфолипазу А2 (фосфорилирует фосфолипиды мембраны), арахидоновую кислоту (расщепляется циклооксигеназой на простагландины Е 2 ) [6].
Методы. Рабочим инструментом явился пакет прикладного компьютерного математического моделирования «Mathcad Professional 2000». Результаты моделирования сравнивались с усредненными экспериментальными данными, полученными в электрофизиологических экспериментах.
-
1. Моделирование ингибиторного действия АТФ и аденозина в синапсах фазных мышц. АТФ оказывает ингибирующее действие на амплитуду ТКП в синапсах фазной мышцы лягушки (рис. 1). Рассмотрим кинетику действия агонистов на синаптический выброс:
k1 k2k3 m3
P Y + ATP ⇔ P YATP ⇔ ÄÀÃ ⇔ ÏÊÑ ⇒ ÂÂ ↓
-
2 l1 2 l2l3
k1 k2 k3 k4 k5 k6m6
P Y + ATP ⇔ P YATP ⇔ ÄÀÃ ⇔ ÏÊÑ ⇔ ÔËÀ ⇔ ÀÊ ⇔ ÏÃÅ ⇒ ÂÂ ↓
-
2 l1 2 l2 l3 l4 2 l5 l6 2
k7m7
PY ′+ Adenosine ⇔ PYAdenosine ⇒ ÂÂ ↑
-
1l7
k8k9 m8
PY + Adenosine ⇔ PY ′ Adenosine ⇔ K + ⇒ ÂÂ ↓
-
1 l8 1l9
Система кинетических уравнений будет выглядеть следующим образом:
-
d [P2YATP] = k 1 ⋅ a ( t ) ⋅ (1 - [P YATP]) -l1 ⋅ [P YATP]
dt 22
-
d [ДАГ] = k 2 ⋅ [PYATP] ⋅ (0.5 - [ДАГ]) -l2 ⋅ [ДАГ]
dt
-
d [ПКС] = k 3 ⋅ [ДАГ] ⋅ (0.5 - [ПКС]) -l3 ⋅ [ПКС]
dt
-
d [ФЛА2] = k 4 ⋅ [ПКС] ⋅ (0.5 - [ФЛА ])-l4 ⋅ [ФЛА ]
dt 22
-
d [АК] = k 5 ⋅ [ФЛА ] ⋅ (0.5 - [АК])-l5 ⋅ [АК]
dt
-
d [ПГЕ] = k 6 ⋅ [АК] ⋅ (0.5 - [ПГЕ ])-l6 ⋅ [ПГЕ ]
dt 22
d [P YAdenosine]
-
1 = k 7 ⋅ b ( t ) ⋅ (1 - [P YAdenosine]) -l7 ⋅ [P YAdenosine]
dt 11
d [P Y ′ Adenosine]
-
1 = k 8 ⋅ b ( t ) ⋅ (1 - [P Y ′ Adenosine]) -l8 ⋅ [P Y ′ Adenosine]
dt 11
-
d [K + ] = k 9 ⋅ [PY ′ Adenosine] ⋅ (0.5 - [K + ]) -l9 ⋅ [K + ]
dt 1
-
d[В[В =-m3⋅[ПКС]-m6⋅[ПКС]+m7⋅[PYAdenosine]-m8⋅[K+]+kk⋅(1-[ВВВ dt
-
2. Моделирование облегчающего действия АТФ в синапсах тонических мыщц. В синапсах тонических мышц теплокровных и
- холоднокровных АТФ, в отличие от сохраняющего свой ингибиторный эффект конечного продукта своего распада – аденозина, потенцирует амплитуду вызванных постсинаптических потенциалов – ПСП [2]. Так, при аппликации АТФ в концентрации 100 мкМ амплитуда ПСП мышцы круралис озерной лягушки возрастала до 127±5% (n=6) относительно контроля, в то время как аденозин в той же концентрации снижал амплитуду ПСП до 70±6 (n=6). Неселективный антагонист Р2 рецепторов сурамин и блокатор протеинеиназы С хелеритрин устраняли потенцирующий эффект АТФ на амплитуду постсинаптических ответов синапсов тонических мышц.
Система уравнений решалась методом Рунге-Кутта с фиксированным шагом. Проведенное нами математическое моделирование показало, что наиболее точное воспроизведение амплитудных и временных характеристик опытных данных наблюдается при следующих значениях равновесных констант: k1=0,9; l1=0,5; k2=0,8; l2=0,5; k3=0,7; l3=0,55; k4=0,7; l4=0,75; k5=0,65; l5=0,7; k6=0,5; l6=0,4; k7=0,9; l7=0,87; k8=0,58; l8=0,9; k9=1; l9=1; m3=3; m6=10; m7=0,2; m8=4; kk=1.
Аденозин действует непосредственно на синаптический выброс (А2) или же через активацию калиевых каналов (А1), что отражается на амплитудно-временной характеристике более резким ингибированием выброса, нежели при введении АТФ, влияние которого осуществляется через систему посредников, включающей целый каскад реакций. Хотя каждая реакция эффективно катализируется, в совокупности они приводят к запаздывающему действию агониста.
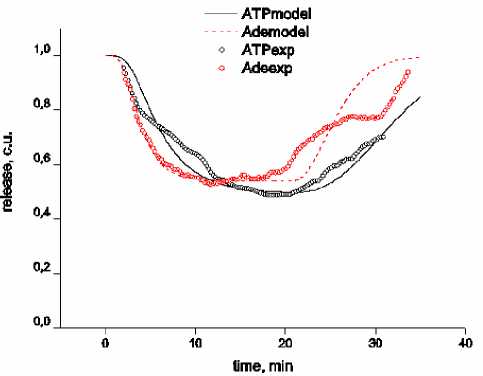
Рис. 1. Кривые действия АТФ и аденозина в концентрации 100 мкМ на амплитуду токов концевой пластинки, полученные в результате эксперимента (exp) и моделирования (model)
Рассмотрим кинетику облегчающего действия АТФ на синаптический выброс, задействующего пресинаптические Р2 рецепторы и протеинкиназу С, в синапсах тонических мышц. По экспериментальным данным изменения амплитуды постсинаптических ответов проведена полиномиальная аппроксимация методом наименьших квадратов. По заданным в дискретные моменты времени ti значениям функции yi(i=1,2,…,n) требуется определить коэффициенты полинома y(t)=a0 +a1t +a2t2 +…+amtm степени m≤n так, чтобы сумма квадратов отклонений экспериментально измеренных значений yi от значений функции y(t) в заданных точках ti была минимальной, т.е. чтобы величина среднеквадратического отклонения
n
-
5 2 = Z( У ( t J - y i )2 i = 1
достигала минимального значения. Коэффициенты полинома a0,a1,a2, ,am доставляющие величине δ2 наименьшее значение, находятся из решения системы линейных алгебраических уравнений n n nn a 0 ' n + a 1 'Z ti + a 2 'Z ti2 + ... + am 'Z tm = Z Vi i=1 i=1 i=1
n n nnn a о 'Z ti + a 1 'Z tf + a 2 'Z t3 + ... + am 'Z tm+1 = Z У. ' ti i=1 i=1 i=1 i=1
n n nnn m +1 *m + 2 V^.2 mV''
ti + a 1 'Z ti + a 2 'Z ti + ... + am 'Z ti =Z yi' i=1 i=1 i=1 i=1
На рис. 2 приведены результаты математического моделирования аппроксимации динамики действия АТФ полиномом седьмой степени. Значения коэффициентов аппроксимирующего полинома приведены в таблице 1.
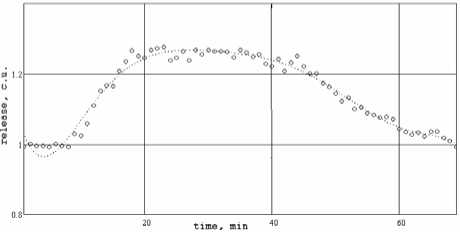
Рис. 2. Точками отмечены экспериментально измеренные значения амплитуды постсинаптических ответов под действием АТФ в синапсах тонических мышц озерной лягушки, непрерывная кривая – аппроксимирующий полином седьмой степени
Выводы: результирующие кривые математического моделирования в общем повторяют экспериментальные данные. Были получены конкретные значения коэффициентов полинома и значения равновесных констант. Поддержано грантом РФФИ.
Список литературы Пуринергическая регуляция синаптической передачи фазных и тонических мышц позвоночных
- Зиганшин, А.У. Р2-рецепторы: перспективная мишень для будущих лекарств/А.У. Зиганшин, Л.Е. Зиганшина. -М.: Геотар-Медиа, 2009. 136 с.
- Гришин, С.Н. Влияние АТФ на тонические мышечные волокна озерной лягушки/С.Н. Гришин, Р.Р. Камалиев, А.У. Зиганшин//Сборник докладов XX Съезда Физиологического Общества им. И.П. Павлова. -Калуга, 2007. С. 204.
- Giniatullin, R.A. ATP and adenosine inhibit transmitter release at the frog neuromuscular junction through distinct presynaptic receptors/R.A. Giniatullin, E.M. Sokolova//Br. J. Pharmacol. 1998. V. 124, No. 4. P. 839-844.
- Sokolova, E. Distinct receptors and different transduction mechanisms for ATP and adenosine at the frog motor nerve endings/E. Sokolova, S. Grishin, A. Shakirzyanova et al.//Eur. J. Neurosci. 2003. V. 18, No. 5. P. 1254-1264.
- Grishin, S.N. Role of adenosine receptors in modulation of synaptic transmission/S.N. Grishin, A.V. Galkin, E.G. Priazhnikov et al.//Neurophysiology. 2002. V. 34, № 2/3. P. 155-157.
- Grishin, S. Mechanisms of ATP action on motor nerve terminals at the frog neuromuscular junction/S. Grishin, A. Shakirzyanova, A. Giniatullin et al.//Eur. J. Neurosci. 2005. V. 21, No. 5. P. 1271-1279.
