Пути повышения выносливости в силовых видах спорта

Автор: Махов С.Ю., Баркалов С.Н.
Журнал: Автономия личности @avtonomiya-lichnosti
Рубрика: Физическая культура, спорт и здоровье человека
Статья в выпуске: 3 (33), 2024 года.
Бесплатный доступ
В статье рассмотрены пути повышения выносливости в силовых видах спорта.
Выносливость, аденозинтрифосфорная кислота, креатин - фосфатная система, гликолитическая система
Короткий адрес: https://sciup.org/142243563
IDR: 142243563
Ways to increase endurance in power sports
The article discusses ways to increase endurance in power sports.
Текст научной статьи Пути повышения выносливости в силовых видах спорта
Практически любая достаточно длительная работа вызывает утомление. Вынос-ливость - способность продолжать работу, несмотря на возрастающее утомление. Тема повышения выносливости является одной из наиболее актуальных, не только для спортсменов, но и обычных людей. «Меньше уставать и лучше справляться с утомлением» - кто из нас откажется от такой способности? Неудивительно, что вопросы, связанные с выносливостью, активно и всесторонне изучают. Совершенствуют методы исследований. Углубляют понимание физиологических процессов. Накапливают методические наработки.
Но вместе с повышением объема знаний становится сложнее и интерпретировать, и использовать на практике эти знания. «Общая выносливость»
смешивается с «аэробной емкостью», взбалтывается с «молочной кислотой», употребляется со «специфичным утомлением» и запивается «последовательным вовлечением систем энергообеспечения».
С выносливостью, как с любым сложным и многогранным явлением, часто происходит следующее. Сначала создаются модели для упрощения комплексных процессов и их понимания. Затем эти модели и абстракции возводят в абсолют. Люди начинают считать, что модели тождественны описываемым явлениям. Возникает винегрет из моделей и концепций разного рода, которые должны упростить, но по факту все усложнили настолько, что остается нехитрый выбор: методический паралич, или работа по наитию, или «как всегда делали».
Абстрактные понятия часто не помогают при принятии практических взвешенных решений. Абстракции не помогают и разговаривать специалистам на одном языке: слишком разных смысл вкладывают в одни и те же модели и концепции. Чтобы найти этому подтверждение, достаточно послушать практически любой разговор двух специалистов на тему «общей выносливости» или «аэробной емкости».
Общее понятийное пространство для разных абстрактных концепций и школ мысли создает физиология физической работоспособности. Нам не обязательно понимать, как относится ударный объем сердца к «общей выносливости» или «аэробной емкости». Это вообще не важно, если мы знаем, какую роль играет ударный объем сердца в физической работе.
В тренировочном процессе можно решать разные задачи: как снизить процент подкожной жировой клетчатки, нарастить мышечную массу, улучшить метаболизм и маркеры здоровья, бежать дольше, плыть быстрее, поднимать более тяжелые снаряды. Для человеческого организма во всех этих случаях приоритет всегда один, и он не меняется. Этот приоритет - выживание.
В процессе эволюции и естественного отбора выживают наиболее приспособленные к текущим условиям виды, которые успешно решают задачи поиска пищи, избегания угроз и воспроизводства. Неисчислимое разнообразие видов решает эти задачи совершенно отличными способами: паразитируют, мимикрируют, объединяются в колонии и стаи, занимают свободные биологические ниши и т. д.
Многим биологическим видам не нужно движение. Но млекопитающие обяза-ны двигаться, чтобы выжить. И люди, которые в современных условиях могут себе позволить вести очень пассивный образ жизни, исключением не являются. Несмотря на кардинальное изменение среды обитания, механизмы энергообеспечения, двигательного контроля и утомления, которые сформированные за миллионы лет эволюции, продолжают действовать.
Произвольные движения обычно решают одну или несколько задач: поиск пищи, воспроизводство, избегание опасности, груминг. Целесообразные произвольные движения называют двигательными задачами .
Любая двигательная задача начинается с цели. Выкопать питательный клу- бень, перетащить тушу кабана к стоянке, убежать от хищника - все это конкретные, биологически целесообразные двигательные задачи.
Для жизнедеятельности организма требуется энергия, а запас ее в организме ограниченный. Для того, чтобы не допустить истощения и не создать угрозу выживания, в организме действуют защитные предохранители - механизмы утомления. А выносливость - это система компенсации механизмов утомления, которая позволяет продолжать биологически целесообразную работу [1].
При выполнении физической работы определенной интенсивности одновременно происходит множество процессов, складывающихся в сложный физиологический паззл. Соответственно, когда решается задача повышения выносливости, важно понимать, за счет каких адаптаций можно получить прирост работоспособности. Ответ никогда не выглядит как «надо больше митохондрий» или «нужен больше ударный объем» или «надо больше терпеть».
Способов повлиять на выносливость много, и полезно знать их все. Тогда в конкретных, индивидуальных обстоятельствах можно сделать акцент на наиболее целесообразных адаптациях.
Поддержание постоянного уровня энергии в клетке
Не только двигательная активность, но и функционирование вообще любых органов и тканей требуют энергии. С пищей мы получаем из окружающей среды питательные вещества - белки, жиры, углеводы. В процессе метаболизма эти пита-тельные вещества усваиваются, и дальше организм их использует как строительный материал или топливо. Что бы ни утверждали солнцееды и прочие автономы, без регулярного поступления питательных веществ выжить не получится. С другой стороны, запас уже усвоенных пластических и энергетических ресурсов всегда ограничен.
Энергия, которая питает процессы жизнедеятельности в организме человека, связана в виде молекулы аденозинтрифосфорной кислоты (АТФ). Говорят, что АТФ - универсальная энергетическая «валюта» организма. Что это значит? Несмотря на то, что энергию могут накапливать и переносить разные химические соединения, клетки организма человека могут использовать только АТФ. Это как национальная валюта и все остальные иностранные валюты, имеющиеся на территории государ-ства: ценность имеют все валюты, но что-то купить можно только за национальную валюту. И если надо использовать иностранную, ее сначала надо обменять.
АТФ используется вообще во всех процессах жизнедеятельности клетки. В результате так называемой реакции гидролиза с участием воды и фермента АТФ-азы молекула АТФ расщепляется на аденозиндифосфат (АДФ) и неорганический фосфат. Процесс сопровождается выделением энергии, которая идет на нужды клетки. [2].

Энергия для жизни и работы клетки требуется непрерывно. Потому и процессы расщепления и высвобождения энергии из АТФ происходят постоянно, каждую секунду. Сохранение достаточной концентрации АТФ в клетке - это вообще первичный приоритет выживания. Если запас энергии в клетке упадет ниже критического, остановятся процессы жизнедеятельности, и клетка попросту погибнет. При этом запасы АТФ в клетке ограничены: при максимальной интенсивности физиологи-ческих процессов АТФ в мышечном волокне хватает всего на 1-3 секунды.
Но если энергия нужна непрерывно, а запасы АТФ ограничены, как организм справляется? АТФ после гидролиза можно восстановить и снова использовать (ре-синтезировать): АДФ может присоединить обратно Ф, и получится целая молекула АТФ [2, 3].
Процесс присоединения неорганического фосфата называется фосфорилированием . Но есть нюанс - для восстановления АТФ нужна энергия. Поскольку поддер-жание уровня энергии - залог выживания клетки, ресинтез АТФ может осуществлять-ся не одной, а сразу 3 системами: креатинфосфатной, гликолитической и аэробной.
Креатинфосфатная система (другие названия - фосфагенная или система АТФ-КФ) использует креатинфосфат (КФ) для ресинтеза АТФ. Креатинфосфат -еще одно соединение, способное накапливать и передавать энергию, только эту энергию нельзя напрямую использовать в клетке. Зато ее можно задействовать для восстановления АТФ. По этой причине КФ выступает своеобразным буфером энергии. Если вернуться к аналогии с валютой - за креатинфосфат нельзя ничего «купить», но можно свободно и почти мгновенно конвертировать при необходимости в национальную валюту и совершить покупку.

В каждый момент времени в клетке требуется энергия, для получения которой расщепляется АТФ. Эту АТФ сразу надо восстанавливать, ведь запасы ограничены. Самый быстрый способ - через расщепление креатинфосфата. Как только в клетке образуется АДФ, при участии фермента креатинкиназы КФ с легкостью отдает молекулу фосфора и энергию, восстанавливая АТФ. Но запасы КФ в клетке тоже ограни-чены. При максимальном энергозапросе их хватает всего на 8-10 секунд. По этой причине, как только молекула КФ расщепляется на креатин и фосфат, приоритетно ее восстановить. В состоянии покоя энергии тратится немного, и восстанавливается КФ почти мгновенно. Ресинтез КФ - залог того, что клетка всегда будет готова к максимальному запросу энергии.
Для ресинтеза КФ тоже нужна энергия (как и для всего в организме). Откуда берется эта энергия и что восстанавливает креатинфосфат? Тут в дело вступают гликолитическая и аэробная системы. Спусковым крючком для запуска этих систем тоже становится повышение концентрации АДФ в клетке [3].
Актуальные данные говорят в пользу того, что гликолитическая и аэробная системы, как минимум в состоянии покоя, не только производят энергию для потребностей клетки, но и обеспечивают восстановление КФ. Это, на первый взгляд, нелогично: зачем тратить АТФ на восстановление КФ, чтобы потом КФ отдавал энергию АТФ? Тут есть 2 объяснения.
Во-первых , КФ гораздо свободнее перемещается по внутриклеточному пространству, чем АТФ. И если АТФ производится в митохондрии, а нужна в другой области клетки (например, для работы ионных натрий-калиевых насосов), быстрее восстановить КФ, чем транспортировать АТФ через мембраны митохондрии и внутри клетки.
Во-вторых , запас креатинфосфата - это гарантия, что клетка будет готова к максимальному энергозапросу. Если возникнет ситуация из категории «бей-беги», необходимо иметь запас энергии. Такие ситуации - почти всегда непредвиденные и всегда требуют быстрого проявления высокого уровня силы, а потому надо поддерживать запас креатинфосфата в максимально полном объеме.
Следует отметить, что роль энергосистем меняется при высоком энергозапросе: тогда вся АТФ, производимая одновременно тремя системами, идет на нужды клетки. Ресинтез КФ отходит временно на второй план, и его запасы снижаются. Даже при максимальном энергозапросе уровень АТФ в клетке падает не больше, чем на 15-20%. А вот запас креатинфосфата при этом может упасть на 60-80%3.
Гликолитическая система - анаэробная, как и креатинфосфатная. Для ее работы не требуется кислород. Необходимы только топливо в виде глюкозы и ферменты. Ну и немного энергии, конечно. В процессе расщепления глюкозы (гликолиз), который требует 2 АТФ и состоит из 10 последовательных реакций, одна молекула глюкозы распадается на 2 половинки - молекулы пирувата -и дает 4 АТФ. То есть в результате реакции гликолиза клетка получает всего 2 АТФ с каждой молекулы глюкозы. Это не очень экономично, зато быстро. А еще в процессе гликолиза почти не вырабаты-вается метаболическое тепло [1, 2].
Если энергозапрос в моменте высокий, и клетке не хватает окислительных способностей или кислорода - гликолиз позволяет выполнить задачу.
Глюкоза + 2NAD+ + 2АДФ + 2Ф —► 2пируват +2NADH + 2Н+ + 2АТФ + 2Н2О
Запасы глюкозы в организме конечны, и при высокой мощности работы заканчиваются относительно быстро. Но гораздо быстрее в клетке снизится концентрация кофермента NAD + , который необходим для протекания гликолиза. NAD + - окисленная форма никотинамиддинуклеотида, и в результате гликолиза восстанавливается до NADH. Как только концентрация NAD+ снизится до определенного уровня, глико-лиз попросту остановится. В условиях нехватки кислорода или окислительных возможностей, это означает прекращение синтеза энергии, что не может быть хорошо для клетки. Но благодаря образованию лактата, этого можно избежать.
Лактат в спорте - традиционно антагонист выносливости и источник всех спор-тивных бед. Но лактат в организме выполняет ряд полезных функций. Окисление NADH до NAD + лишь одна из них.
При участии фермента лактатдегидрогеназы (ЛДГ) пируват преобразуется в лактат, забирая обратно протон водорода, а NADH окисляется до NAD+. Таким образом, с одной стороны, буферируются свободные протоны водорода (реальная причина утомления), а с другой - гликолиз может продолжать синтезировать АТФ.
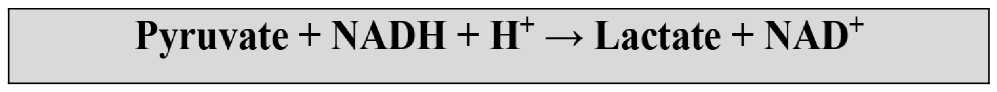
В некоторых клетках, например, эритроцитах, кислород не может использоваться в принципе, и возможен только гликолиз. В условиях покоя вклад гликолиза в общий ресинтез АТФ невысокий. А весь пируват, который образуется в процессе, транспортируется в митохондрии и там окисляется по аэробному пути. В условиях напряженной мышечной работы мощность гликолиза растет пропорционально энергозапросу. При околопредельной интенсивности гликолиз дает много энергии, но работает короткое время (от 30-90 с), а далее останавливается из-за накопления протонов водорода. [3].
Аэробная система производит гораздо больше энергии из каждой единицы «топлива», но процесс протекает медленнее. Кроме того, для аэробной системы требуется кислород, а в моменте в клетке его не всегда достаточно. Реакция окислительного фосфорилирования происходит в специализированных органеллах – митохондриях . При наличии кислорода митохондрии в качестве субстрата могут использовать и жирные кислоты, и пируват, образовавшийся в результате гликолиза, и даже аминокислоты. Все эти субстраты преобразуются в единую форму (ацетил-кофермент) и проходят через процесс, называемый циклом Кребса (также цикл лимонной кислоты, цикл трикарбоновых кислот). Аэробные процессы завершаются окислительным фосфорилированием .
В процессе окислительного фосфорилирования одна молекула пирувата (половина глюкозы) дает 30-38 АТФ. Одна молекула жирной кислоты - 120-130 АТФ. Эта энергия используется для восстановления креатинфосфата в меж- мембранном пространстве митохондрий, а также транспортируется из митохондрий в цитозоль (жидкое содержимое клетки) и далее по месту требования. КФ способен пересекать мембраны митохондрий гораздо свободнее и быстрее, чем АТФ, потому одна из задач аэробной системы - обеспечить энергией ресинтез КФ [2].
В условиях работы на предельной интенсивности энергозапрос такой, что совокупной мощности работы окислительной и гликолитической системы не хватает для энергообеспечения (рис. 1). По этой причине, как только запасы креатинфосфата истощаются, интенсивность работы неконтролируемо падает.
Иногда встречаются мнения, что системы ресинтеза АТФ включаются последовательно: сначала фосфагенная, при исчерпании запасов креатинфосфата гликолити-ческая и еще позже аэробная. И если, например, поработать всего 23 секунды, это будет «алактатная» работа, то есть без участия гликолитической системы.
При детальном изучении становится очевидно, что все три системы работают одновременно, просто с разной скоростью. И даже если вклад окислительной и гликолитической систем во время предельно мощной короткой работы не очень существенный, картина меняется при повторении усилий и в интервалах отдыха.
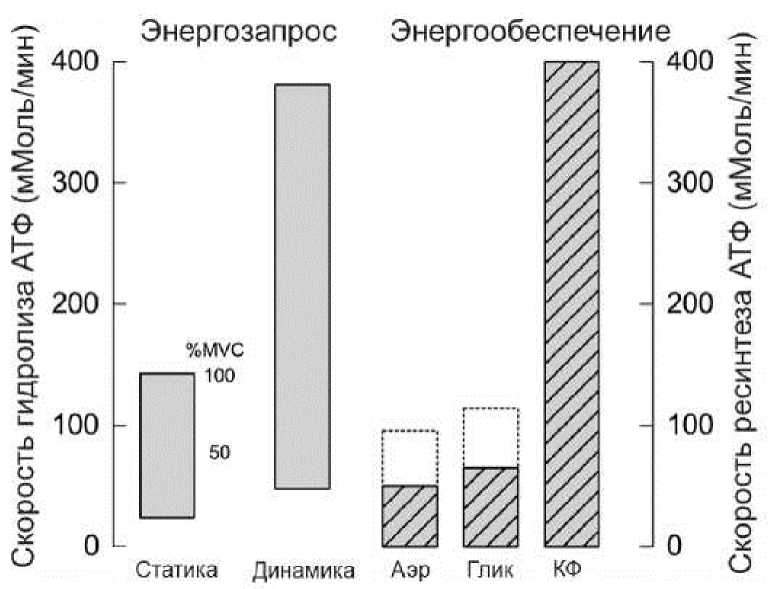
Рис 1. Энергозапрос и способность разных систем к ресинтезу АТФ
Можно подумать, например, что спринтерский бег на короткие дистанции - это алактатная работа (то есть без участия гликолитической системы и образования лактата). Но расчетный вклад гликолиза в энергообеспечение может составить около 50%ii. А после спринта, длящегося 10-11 секунд, уровень лактата продолжает расти в течение нескольких минут, достигая значений более 10 ммоль [2, 3].
Одновременная работа всех систем биологически целесообразна с учетом непрерывных затрат энергии на процессы жизнедеятельности и постоянной необходимости ее восполнять (рис. 2).
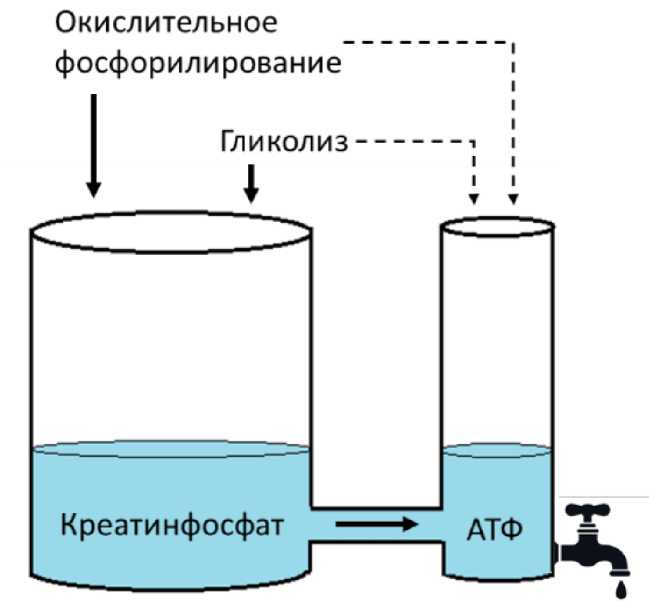
АТФ
Окислительное фосфорилирование
Креатинфосфат
Гликолиз
Рис 2. Работа всех систем энергообеспечения
Для ресинтеза АТФ нужны энергоресурсы, которые поступают в виде питательных веществ извне. Чтобы добыть питательные вещества - необходимо прикладывать физические усилия. Поэтому способность выполнять физическую работу является важным, биологически целесообразным приоритетом для организма.
Любая физическая работа управляется нервной системой, исполняется скелетно-мышечной и обслуживается кардиореспираторной. Бессмысленно рассуждать, что важнее: сердце, мышцы, легкие или извилины. Без любого из этих элементов невозможно выполнить сколько-нибудь длительную работу. А вот понимать, как каждая из систем участвует в работе и как влияет на выносливость - важно и полезно. На рис. 3 представлена схема происхождения сопряжения управляющей, исполнительной и обслуживающей систем при выполнении произвольного движения [3].
Двигательная задача

Биологическая целесообразность движения

Центральное
Управляющий аппарат
Центральная мереная система
Исполнительная система
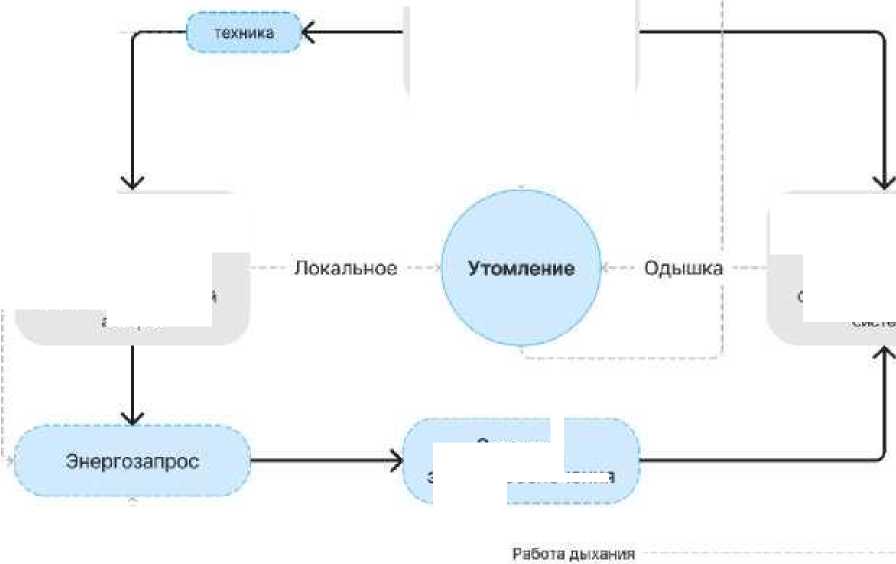
Обслуживающий аппарат
Опорно-двигательный аппарат
Сердечно-сосудистая система
Системы энергообеспечения
Рис 3. Сопряжение управляющей, исполнительной и обсуживающей системы
Техника и снижение энергозапроса
Движение является для организма приоритетной задачей, пока оно биологически целесообразно. Но любая двигательная задача сопряжена с тратой энергии, а последняя исторически всегда была в дефиците. И если расхода энергии нельзя избежать, можно хотя бы научиться ее экономить.
Человеческий организм – прекрасный пример адаптации к физической работе, в первую очередь, за счет экономизации усилий. Задолго до того, как в клетках станет больше митохондрий и прорастут новые капилляры, а сердце станет способно прокачивать большие объемы крови, организм научится снижать расход энергии, требуемый для выполнения того же объема работы. Снижение энергозатрат происходит за счет нескольких взаимодополняющих процессов [1].
-
1. Отсечение лишних мышечных напряжений . При выполнении новых
-
2. Расслабление внутри цикла движения . Оно позволяет добавить в каждое повторение микрофазу восстановления после фазы напряжения, что оказывает существенное влияние на предельную продолжительность работы.
-
3. Использование эластичных свойств мышц и соединительных тканей . Мышцы и сухожилия обладают разными характеристиками, в том числе вязкоупругими. При быстром растяжении мягких тканей в соединительных тканях и структурных белках запасается энергия упругой деформации. Если после быстрого растяжения незамедлительно следует быстрое сокращение, эта энергия упругой деформации трансформируется в кинетическую энергию. Такой механизм называется «цикл растяжения-сокращения» (ЦРС) и позволяет снизить потребности в метаболической энергии при фиксированной мощности. ЦРС также помогает повысить выходную мощность при суммации метаболической и упругой энергии. В любых циклических движениях (выполняемых более 1 раза) будет возможность для использования упругих свойств мягких тканей. В некоторых ациклических движениях тоже может присутствовать ЦРС (вертикальный прыжок, приседание).
-
4. Использование внешних сил . В движении на тело действуют разные силы: тяжести, инерции, трения, реакции опоры, упругости, аэродинамического сопротивления. Использование этих сил также позволяет экономить метаболическую энергию. Например, лучшее сцепление со снарядом за счет обработки магнезией позволяет снизить нагрузку на мышцы предплечья. Небольшой отбив штанги в многоповторной становой тяге позволяет использовать силу реакции опоры и упругие свойства грифа и дисков, что существенно облегчает первую треть амплитуды движения. В каждом виде спорта или двигательной задаче есть свои параметры внешних сил, которые можно научиться использовать для экономии собственной метабо-лической энергии.
-
5. Эффективность кинематических цепей . Кинематическая цепь - это совокупность вовлеченных в движение подвижных звеньев. Любое комплексное движение требует приложения силы в определенной точке и передачи усилия к другим подвижным звеньям или снаряду. Например, становая тяга: мышцы-разгибатели ног генерируют усилие, которое спортсмен прикладывает к помосту. Действующая в обратном направлении сила реакции опоры передается через туловище, плечевой пояс и руки к снаряду. Снаряд поднимается вверх. Все тело выступает как единая кинематическая цепь передачи усилия. Усилие может передаваться эффективно, с минимальными потерями энергии
-
6. Экономичность внешнего дыхания . Работа дыхания обеспечивается респираторными мышцами. В покое для вдоха хватает работы диафрагмы, а выдох и вовсе происходит пассивно за счет эластичности легких, мышц живота и соединительных тканей грудной клетки. При более активном дыхании подключаются дополнительные респираторные мышцы, в том числе вторичные. Дыхание регулируется непроизвольно или произвольно в зависимости от ситуации и навыков человека. Если человек начинает дышать слишком интенсивно для текущего энергозапроса, то растет работа дыхания по преодолению аэродинамического сопротивления дыхательных путей и упругости мягких тканей. В процессе тренировок дыхание становится более экономичным, а значит, дыхательные мышцы меньше устают и не создают конкуренции с локомоторными мышцами за кровоснабжение.
-
7. Вариативность техники . Если человек владеет только одним способом выполнения упражнения, то при утомлении задействованных мышц придется снижать темп или делать перерывы в работе. В некоторых случаях вариантов техники выполнения движения немного. Но часто одну и ту же двигательную задачу можно решить несколькими техническими способами, особенно если это многосуставные упражнения, выполняемые циклически (как в функциональном многоборье). Это касается как мышц в рамках одной мышечной группы, так и мышечных групп. Например, угол отведения плеча в отжиманиях в упоре лежа смещает акценты работы между волокнами большой грудной мышцы, передних дельтовидных и трех-главой мышцы плеча. А если речь о том, чтобы быстро принять положение лежа на полу и подняться - это можно выполнить десятком способов, в том числе без использования рук.
-
8. Вариативность дыхания . Поскольку дыхание - один из главных приоритетов, функции респираторных мышц могут обеспечивать почти все мышцы туловища. На вдохе могут помогать передние и задние верхние зубчатые, малые грудные, верхние волокна трапециевидной мышцы, лестничные, грудино-ключично-сосцевидные мышцы. В зависимости от условий (фиксированный плечевой пояс, пережатая брюшная стенка и т. д.) в соответствии с принципом нейромеханического сопряжения включаются те дыхательные мышцы, которые имеют наилучшее плечо силы в данный момент.
упражнений возникает генерализация нервного возбуждения. Преобладает торможение, движения закрепощенные, напрягается много лишних мышечных групп. При многократном выполнении подобной работы достаточно быстро нервная система разбирается, напряжение каких мышц реально важно для результативности двигательной задачи, а какие сокращать не нужно. Этот процесс происходит в любом случае, во многом бессознательно. Но может быть ускорен за счет осознанного и запланированного совершенствования техники.
в подвижных звеньях (поясничный и грудной отделы позвоночника, лопаточногрудной сустав, суставы кисти и пальцев). Но могут быть и серьезные потери: штанга может выскальзывать из рук, а спина может выгнуться дугой даже без отрыва штанги от пола.
Вариативность техники хоть и не снижает абсолютный энергозапрос, но позволяет распределить его во времени более равномерно для разных мышечных групп, без концентрации нагрузки только на один регион.
Этот процесс можно во многом сделать сознательным, произвольно варьируя дыхательные мышцы. Так можно обеспечить восстановление некоторых дыхательных мышц и отсрочить накопление локального утомления [2].
Даже в относительно координационно простых циклических видах спорта доказана многолетняя экономизация техники и за счет этого снижение потребления кислорода на конкретном уровне скорости бега. Примером может послужить Пола Рэдклифф, одна из лучших бегуний на длинные дистанции, долгое время удерживавшая рекорд в марафонской дистанции 2 часа 15 мин 25 сек. Несмотря на то, что МПК у нее в течение многолетней карьеры не особенно менялся, снижение кисло-родной стоимости бега на фиксированной скорости и повышение скорости бега на МПК росло, что напрямую свидетельствует о росте экономичности бега. Тем более, нет потолка совершенствования техники и снижения энергозапроса в более координационно сложных видах двигательной активности.
Резерв силы, скорости и скоростной силы
Любое упражнение обладает динамическими (сила) и пространственновременными (ускорение, скорость) характеристиками в разных сочетаниях. В становой тяге требуется проявить высокий уровень силы, но нет необходимости делать это быстро. В теннисной подаче - наоборот. В фазе ускорения спринта требуется и то, и другое в высокой степени. Требуемый уровень силы и скорости определяет то, какие двигательные единицы и в каком количестве будут рекрутированы.
При чем тут выносливость? Зависимость выносливости от силы очень проста: чем сильнее каждая отдельная двигательная единица, тем меньше двигательных единиц надо включать для выполнения каждого цикла работы . Это означает, что возникает запас двигательных единиц для рекрутирования. Вначале всегда вклю-чаются самые низкопороговые двигательные единицы, силы которых достаточно для выполнения работы. Они же и наиболее устойчивые к утомлению. Но они все равно в определенный момент устают. И тогда подключаются резервные, более высокопороговые двигательные единицы. Чем больше резерв двигательных единиц, тем дольше можно продлить работу. Это тем более важно, что во время работы одних ДЕ другие имеют возможность восстановиться и вновь включиться в работу позже.
Для понимания рассмотрим упрощенный пример: четырехглавая мышца бедра до гипертрофийного цикла и после. И задача - разгибание голени в тренажере до отказа с фиксированным весом 30 кг. До адаптации 1 повтор разгибания требует рекрутирования большого количества имеющихся двигательных единиц, чтобы в сумме они выдавали тягу для подъема 30 кг (предположим, 60%). Из-за этого предельное количество повторений до отказа не очень большое - всего около 20.
Но после тренировок на гипертрофию (возьмем 16 недельный цикл), поперечник волокон увеличился. Гипертрофия затронула и низкопороговые, и высокопороговые двигательные единицы. Это значит, что каждое волокно при сокращении проявляет больше силы. И там, где раньше требовалось включать сразу 60% доступных двигательных единиц, теперь достаточно всего лишь 40%.
При прочих равных, после гипертрофийной работы появился запас двигательных единиц, которые можно подключать по мере утомления и выключения из работы первоначально рекрутированных волокон. Это создает и запас работоспособности на несколько лишних повторений, даже если локальная мышечная выносливость не изменится.
Рассмотрим еще два примера. Первый: 50 становых тяг 100 кг на время. 3 спортсмена с разными силовыми показателями: спортсмен А имеет 1ПМ 105 кг, спортсмен Б имеет 1ПМ 160 кг, спортсмен В обладает 1ПМ 250 кг.
Для первого атлета вопрос выносливости тут вообще не стоит. От него требуют выполнить 50 становых тяг на 95%1ПМ. Каждый повтор будет требовать околопредельного напряжения нервной системы, рекрутирования всех двигательных единиц и создавать максимальный относительный энергозапрос. Задача нереалистична.
Второй спортсмен будет работать на 62,5%. При правильной разбивке объема на серии с коротким отдыхом после нескольких подъемов можно выполнить задание эффективно.
При прочих равных (одинаковый уровень локальных и центральных факторов выносливости), третьему будет проще всего. Просто потому, что ему надо будет работать на 40% от 1ПМ. Это значит, что требуемый уровень напряжения систем организма будет ниже. Вначале будут рекрутироваться относительно низкопороговые волокна, более устойчивые к утомлению. От нервной системы не требуется высокой интенсивности работы. Третий спортсмен сможет работать более длинны-ми сериями с меньшим отдыхом.
И другой пример: футбол, где требуется способность к повторным субмаксимальным ускорениям. Предположим, от игроков требуется за матч часто проявлять усилия для бега со скоростью 6-7 м/с. Один игрок обладает максимальной скоростью бега 10 м/с. Второй - 8 м/с. Каждый эпизод, где надо развивать скорость 6 м/с, для первого требует работы на 60% максимальной скорости, и для второго - на 75%. Это, как и в примерах выше, означает разное напряжение и управляющей, и исполнительной, и обслуживающей систем. А значит, и разный запас по вос-произведению таких усилий.
Понятно, что степень переноса силовой работы на выносливость ограничена и зависит от специализации спортсмена. В циклических видах спорта на длинные дистанции перенос будет наименьший, и в зоне интереса будут в основном сила окислительных волокон.
Очевидно, что существует и некая верхняя планка силовых показателей, достаточных для оптимальной выносливости. Для понимания необходимого уровня силы в каждом популярном виде спорта можно использовать нормативные данные по силе, накопленные на основе анализа большого количества спортсменов.
Также надо учитывать потенциальное ограничение: рост поперечника волокна может ухудшить его кровоснабжение, так как количество капилляров не увеличивается в обязательном порядке пропорционально поперечнику волокна, это две разных адаптации. В этой связи имеет смысл даже в силовых циклах на гипертрофию выполнять поддерживающую работу на выносливость, направленную, в том числе, на развитие капиллярной сети соответственно росту мышечной массы.
Любая дополнительная мышечная масса увеличивает затраты энергии просто на движение конечностей и противодействие внешним силам. По этой причине к вопросу повышения работоспособности за счет силовых показателей и гипертрофии следует подходить осторожно, анализируя потенциальные плюсы и минусы в каждом отдельном случае.
Наилучший перенос силовой работы на выносливость будет наблюдаться в видах спорта с переменной интенсивностью (единоборства, игровые виды), а также смешанных двигательных задачах с проявлением локальной силовой выносливости разных мышечных групп [2, 3].
Аэробные возможности мышц
Вне зависимости от уровня мощности работы, чем лучше окислительные возможности задействованных двигательных единиц, тем медленнее будут развиваться процессы утомления, и тем быстрее будет происходить восстановление по окончании работы.
Аэробные возможности мышц складываются из нескольких компонентов:
-
- количество и размер митохондрий;
-
- плотность капиллярной сети;
-
- активность окислительных ферментов.
Волокна I типа обладают наилучшими окислительными возможностями изначально, по своему строению. Промежуточные волокна IIа без тренировок на вынос-ливость утомляются достаточно быстро, так как за ненадобностью не обладают ни большой митохондриальной массой, ни плотной сетью капилляров. Однако эти волокна очень хорошо реагируют на тренировку выносливости. Промежуточные волокна не могут стать настолько же устойчивыми к утомлению, как окислительные, но аэробный потенциал все равно существенный.
Что касается волокон IIb, они характеризуются большими запасами гликогена и концентрацией гликолитических ферментов, но низкой капилляризацией и митохондриальной массой [3].
От суммарной способности мышечных волокон в составе мышцы потреблять кислород зависит ее показатель пикового потребления кислорода. Пиковое потребление кислорода - это максимальный объем кислорода, который может потреблять мышца, мышечная группа или группа мышц при высокоинтенсивной работе.
Разные по размеру и типу двигательные единицы требуют разных протоколов для повышения аэробных способностей. Волокна I типа изначально очень аэробные. Однако при объемной работе низкой и умеренной интенсивности можно по-высить эффективность утилизации жиров в качестве энергосубстрата, а также эффек-тивность окисления лактата, который приходит через кровоток от высокопороговых двигательных единиц. Но работа низкой интенсивности не задействует ВПДЕ. Нужна более высокая мощность работы. Такую работу необходимо организовывать в интервальном формате, после интервала работы давая требуемый для восстановления отдых.
Капиллярная сеть очень хорошо адаптируется к работе на выносливость. Капилляры растут в ответ на биохимические и механические стимулы. Биохимические стимулы связаны в основном с активацией фактора роста эндотелия сосудов и оксидом азота. Механический стимул вызывается напряжением сдвига, возникающем при усилении кровообращения [3].
У тренированных спортсменов плотность капиллярной сети может на 40% и более превосходить такой показатель у нетренированных людей. Есть данные, что ангиогенез (рост новых капилляров) может быть специфичен интенсивности работы. Низкоинтенсивная работа приводит к росту капиллярной сети в окислительных волокнах, но не в ВПДЕ. Для капилляризации ВПДЕ необходимо использовать высокоинтенсивные интервальные протоколы, в которых эти волокна рекрутируются.
Важно помнить: локальная мышечная выносливость развивается в конкретных двигательных единицах конкретных мышц . Один и тот же человек может иметь невероятно аэробные волокна I типа в мышцах ног, посредственную выносливость промежуточных волокон в ногах и очень низкую - в мышцах рук. Профессиональный массажист будет обладать очень приличной локальной выносливостью мышц рук, но может испытывать одышку и забитость мышц ног при банальном подъеме по лестнице на 9 этаж. Прежде, чем выбирать протокол тренировки, необходимо четко понять не только то, какой регион, но и какие двигательные единицы надо натренировать.
Лактатный челнок
Еще один механизм локальной мышечной выносливости - лактатный челнок (рис. 4).
Его роль двояка. Если энергозапрос требует б о льшего вклада гликолиза и образованный пируват не получается полностью окислить, для продолжения реакции гликолиза необходимо восстанавливать NAD+, а образовавшийся при этом лактат вывести из клетки.
После того, как лактат выведен из мышечного волокна в кровоток, он может быть утилизирован сердцем, печенью, почками, мозгом и окислительными мышеч-ными волокнами работающих мышц (в том числе дыхательных).
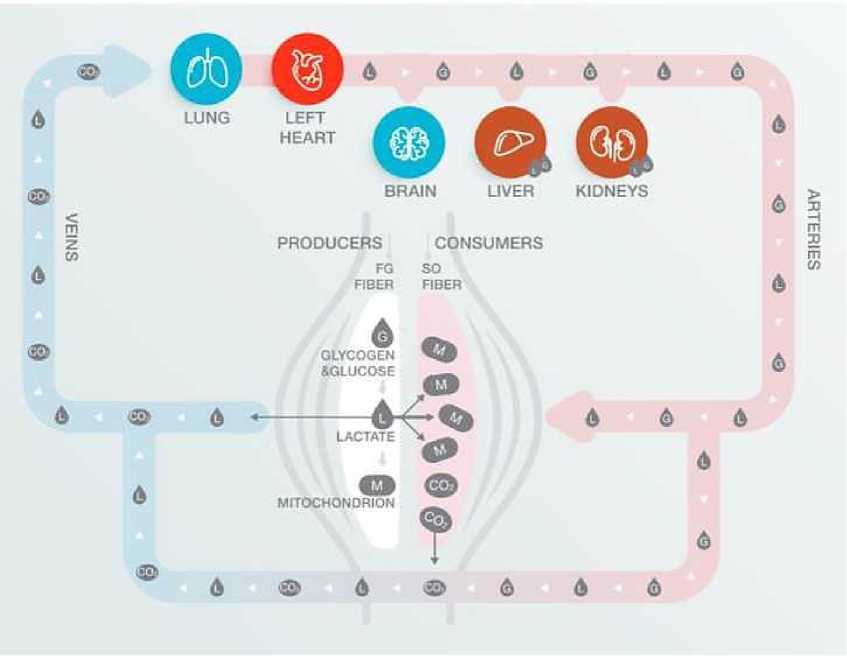
Рис. 4. Лактатный челнок
Тут раскрывается вторая роль лактатного челнока: доставка энергосубстратов по месту требования. Например, окислительные волокна не содержат больших запасов гликогена. Если интенсивность работы требует окисления именно углеводов, то окислительные волокна начинают зависеть от доставки глюкозы с кровью. В клетке глюкоза должна сначала ферментироваться до пирувата, который дальше будет ис-пользован в митохондриях. Лактат в этом смысле гораздо лучше глюкозы, т. к. является ферментированной формой глюкозы и может сразу использоваться в мито-хондриях [1, 2, 3].
Таким образом, лактатный челнок делает возможным продолжение гликолиза в волокнах IIa и IIb, а также доставку энергосубстратов к важным органам и тканям.
Выведение лактата из клетки зависит от плотности капиллярной сети и действия транспортных белков MCT (монокарбоксилат транспортер) и в существенной степени поддается тренировке.
В зависимости от задач, для повышения эффективности механизма лактатного челнока чаще всего используют методы на интенсивности от 1 до 2 лактатного порогов. Но применимы и смешанные методы из 2 упражнений, где лак-татобразующее упражнение чередуется с лактатутилизирующим, варьируя работу разных типов двигательных единиц и мышечных групп.
Буферные системы
Для предотвращения ацидоза в организме работает несколько буферных систем. Часть из них действует непосредственно внутри мышечных волокон.
Это физико-химическое буферирование, метаболическое буферирование и выведение про-тонов Н + из клетки.

Физико-химическое буферирование протекает с использованием белковых соединений (дипептидов карнозина), а также связывания с неорганическим фосфатом в дигидроген фосфат и угольной кислотой в бикарбонат. Эти процессы буфери-руют до 60% всех образуемых протонов Н + .
Еще 40% приходится на метаболическое буферирование , где главный процесс - это креатинфосфатная реакция ресинтеза АТФ. Внутримышечные буферные системы гораздо лучше развиты в высокопороговых двигательных единицах, активно задей-ствующих гликолиз в энергообеспечении [1, 2].
Важнейшее значение играет и буферизация протонов Н+ в крови . Главный механизм - буферирование через бикарбонат, в результате которой образуется Н2О и СО2 (рис. 5).

Рис. 5. Физико-химическое буферирование протонов Н+
Емкость буферных систем ограничена, то есть в какой-то момент при интенсивной работе количество протонов H+ начинает быстро нарастать. Буферные системы показывают адаптацию через 4-8 недель тренировок с существенным вкладом гликолитической системы. Емкость этих систем можно повысить за счет употребления пищевых добавок: креатина, бета-аланина (влияет на карнозин) и пищевой соды (влияет на буферирование через бикарбонат в системе крови).
Список литературы Пути повышения выносливости в силовых видах спорта
- Баркалов С. Н., Махов С. Ю. Особенности функционального состояния кардио - респираторной системы у сотрудников ОВД, занимающихся рукопашным боем // Современное состояние и тенденции развития физической культуры и спорта: сборник научных статей по итогам международной научно - практической конференции (г. Белгород, 26 ноября 2021 г.) / под общей редакцией И. Н. Никулина. Белгород: ИД "Белгород" НИУ "БелГУ", 2021. С. 744-751. ISBN: 978-5-9571-3020-8
- Махов С. Ю., Федин П. Е. Характеристика понятий специальной выносливости как физического качества // Автономия личности. 2021. № 3 (26). С. 110-114. URL: http://avtonomialichnosti.ru/ALM_3_26_2021.pdf (дата обращения: 20.09.2024).
- Махов С. Ю. Физиологические адаптационные изменения в боевых единоборствах при различных уровнях интенсивности нагрузки // Наука - 2020. 2020. № 7 (43). С. 26-35. URL: http://www.nauka-2020.ru/MN_7(43)2020.pdf (дата обращения: 20.09.2024). EDN: PNETRO
