Пыльцевой анализ линий селекционно-генетической коллекции подсолнечника

Автор: Демурин Я.Н., Рубанова О.А., Мелейчук А.В.
Рубрика: Селекция, семеноводство и биотехнология сельскохозяйственных растений
Статья в выпуске: 1 (197), 2024 года.
Бесплатный доступ
Настоящее исследование посвящено изучению продуктивности пыльцы и её морфологической гетерогенности у линий селекционно-генетической коллекции подсолнечника. Исследования проводили во ФГБНУ ФНЦ ВНИИМК, г. Краснодар, в 2022-2023 гг. Использовали линии коллекции: ВК934, ВК1-клп, ВК678, ВК876, In Крупняк, ВК195, МВГ-8, ВК416, Л2138, К1587, КГ49 и RIL200. Для оценки качества пыльцы использовали параметр морфологической гетерогенности пыльцевых зёрен, т.е. долю нетипичных зёрен. Пыльцевую продуктивность оценивали по двум параметрам: масса пыльцы и число пыльцевых зёрен. Использовали микроскоп Биолаб TS-2000 LCD МС 20 при увеличении 40×. Максимальной массой (1,17 мг/пыльник) и числом пыльцевых зёрен (45239 шт/пыльник) характеризовалась линия Л2138. Линия ВК876 имела минимальные значения данных признаков - 0,25 мг/пыльник и 16448 шт/пыльник соответственно. Между признаками числа и массы пыльцевых зёрен отмечена высокая положительная корреляция - r = 0,90. В 2022 г. средний показатель диаметра пыльцевых зёрен у 12 изученных линий варьировал от 30 до 32 мкм при среднем значении 31 мкм. В 2023 г. минимальный показатель диаметра пыльцевых зёрен был ниже, чем в 2022 г., и составил 27 мкм, максимальный находился на уровне 2022 г. - 32 мкм при среднем значении 30 мкм. За два года исследований у линии КГ49 отмечена значительная морфологическая гетерогенность пыльцы, что выражалось в высокой доле дефектных пыльцевых зёрен - 58 %. Линия ВК195 характеризовалась низкой морфологической гетерогенностью с долей дефектных пыльцевых зёрен 5 %.
Подсолнечник, масса пыльцы, гетерогенность, диаметр пыльцевого зерна
Короткий адрес: https://sciup.org/142241298
IDR: 142241298 | УДК: 633.854.78:575 | DOI: 10.25230/2412-608X-2024-1-197-3-9
Analysis of pollen in sunflower lines of a breeding and genetic collection
We studied pollen productivity and its morphological heterogeneity in the lines of the breeding and genetic sunflower collection. The research was carried out at the V.S. Pustovoit All-Russian Research Institute of Oil Crops, Krasnodar, in 2022-2023. Collection lines VK934, VK1-klp, VK678, VK876, In Krupnyak, VK195, MVG-8, VK416, L2138, K1587, KG49 and RIL200 were used. To assess the quality of pollen, we used the parameter of morphological heterogeneity of pollen grains, i.e. the proportion of atypical grains. The pollen productivity was assessed by two parameters: pollen weight and the number of pollen grains. A Biolab TS-2000 LCD MS 20 microscope was used at a magnification of 40×. A line L2138 was characterized by a maximum weight of 1.17 mg/anther and a number of pollen grains of 45,239 pieces/anther. A line VK876 had the minimal values of these traits - 0.25 mg/anther and 16448 pcs/anther, respectively. A high positive correlation of r = 0.90 was noted between these traits. In 2022, the average diameter of pollen grains in 12 studied lines varied from 30 to 32 µm, with an average value of 31 µm. In 2023, the minimal diameter of pollen grains was lower than in 2022 and amounted to 27 µm, the maximum was at the level of 2022 - 32 µm with an average value of 30 µm. Over two years of research, a line KG49 showed significant morphological heterogeneity of pollen, which was expressed in a high proportion of defective pollen grains (58%). A line VK195 was characterized by low morphological heterogeneity with a proportion of defective pollen grains of 5%.
Текст научной статьи Пыльцевой анализ линий селекционно-генетической коллекции подсолнечника
Введение. Изучение мужской генеративной сферы растений применяется в селекции многих сельскохозяйственных культур [1; 2; 3]. В первую очередь это связано с тем, что количество и качество 3
продуцируемой пыльцы во многом определяет формирование нормального урожая [4].
Мужской гаметофит подсолнечника представлен пыльцевым зерном. Его формирование происходит в результате двух следующих друг за другом процессов: микроспорогенеза и микрогаметогенеза. По данным H.T. Horner [5], микроспорогенез у подсолнечника включает 11 стадий. За 2–3 недели до начала цветения клетки археоспориальной ткани, выстилающей полости гнёзд пыльниковой трубки, претерпевают несколько митотических делений. Сформированные в результате митоза микроспороциты делятся мейотически и образуют тетрады микроспор. Далее тетрады распадаются, и микроспоры переходят на этап микрогаметогенеза. Результатом последнего являются двуклеточные пыльцевые зёрна, состоящие из сифоногенной клетки и генеративного ядра. За 2–3 дня до цветения генеративное ядро делится митотически и даёт начало двум спермиям, за счёт которых пыльцевое зерно становится трёхклеточным [6; 7].
Микроспорогенез, особенно на стадии мейоза, довольно уязвим к воздействию внешних и внутренних факторов [8; 9; 10]. А.А. Бабро и О.Н. Воронова [11] выявили, что у Helianthus ciliaris и H. tuberosus дефектные пыльцевые зёрна наблюдаются начиная со стадии тетрад. В сравнении с нормальными пыльцевыми зёрнами они имеют недифференцируемую структуру, нехарактерные толстые стенки, а также отличаются по размеру.
Известно, что на пыльцевую продуктивность у подсолнечника влияют температура воздуха в период микроспорогенеза и его относительная влажность. С ростом относительной влажности воздуха увеличивается и пыльцевая продуктивность [12]. Кроме того, в условиях теплового стресса возрастает доля нежизнеспособных пыльцевых зёрен [13].
Нормальные пыльцевые зёрна имеют сферическую либо эллиптическую форму с тремя порами и бороздами, поверхность покрыта узко коническими шипами длиной до 10 мкм [14]. Диаметр, по разным данным, может изменяться от 28 [15] до
45 мкм [16]. В среднем вес пыльцы в корзинке изменяется от 0,11 до 0,50 г [17], а число пыльцевых зёрен в цветке варьирует от 28 000 до 40 000 шт. [18].
Настоящее исследование было посвящено изучению продуктивности пыльцы и её морфологической гетерогенности у линий селекционно-генетической коллекции подсолнечника.
Материалы и методы. Исследования проводили на втором отделении ЦЭБ ВНИИМК, х. Октябрьский, г. Краснодар, в 2022–2023 гг. Использовали линии селекционно-генетической коллекции: ВК934, ВК1-клп, ВК678, ВК876, I n Крупняк, ВК195, МВГ-8, ВК416, Л2138, К1587, КГ49 и RIL200. Растения выращивали на четырёхрядных (в 2022 г.) и однорядных (в 2023 г.) делянках при густоте стояния 40 тыс. шт/га.
Для оценки качества пыльцы использовали параметр морфологической гетерогенности пыльцевых зёрен, т.е. долю нетипичных зёрен. Собранную пыльцу в небольшом количестве при помощи шпателя помещали на предметное стекло с каплей ацетоорсеина и тщательно перемешивали, чтобы добиться равномерного распределения в растворе. Препарат окрашивали в течение 5–10 минут, затем накрывали покровным стеклом и рассматривали под микроскопом Биолаб TS-2000 LCD МС 20 при увеличении 40×. Диаметр измеряли в 2022 г. на 200 пыльцевых зёрнах, собранных с трех растений одного генотипа, а в 2023 г. – на 100 пыльцевых зёрнах с каждого из трех растений разных генотипов при помощи встроенной в микроскоп шкалы. Параллельно с замерами отмечали дефектные пыльцевые зёрна. Морфологическую гетерогенность пыльцы рассчитывали, как процент дефектных пыльцевых зёрен в общей массе измеренных пыльцевых зёрен.
Пыльцевую продуктивность оценивали по двум параметрам: масса пыльцы и число пыльцевых зёрен. Предварительно изолированные корзинки трёх растений каждого генотипа на 2–3-й день цветения срезали и доставляли в лабораторию. На следующее утро с каждой корзинки до выхода рылец брали 10 пыльниковых трубок для определения массы и 3–4 пыльниковые трубки – для подсчёта числа пыльцевых зёрен.
Для определения массы пыльцы содержимое 10 пыльниковых трубок взвешивали на небольшом листе бумаги, а полученное число пересчитывали на одну пыльниковую трубку. Для подсчёта числа пыльцевых зёрен каждый пыльник помещали в эппендорф, заполненный одним миллилитром воды, вскрывали и тщательно встряхивали. Из полученного коллоидного раствора пипеткой отбирали пробу известного объёма и под микроскопом подсчитывали в ней число пыльцевых зёрен. Полученное значение пересчитывали на объём всего раствора.
Результаты и обсуждение. В 2023 г. максимальное число пыльцевых зёрен отмечено у линии Л2138: в среднем на пыльник приходится 45239 шт. Минимальное число отмечено у линии ВК876 – 16448 шт/пыльник при среднем значении 29471 шт/пыльник и коэффициенте вариации 29 %. Семь линий из 12 имели больше 30000 пыльцевых зёрен в пыльнике (рис. 1).
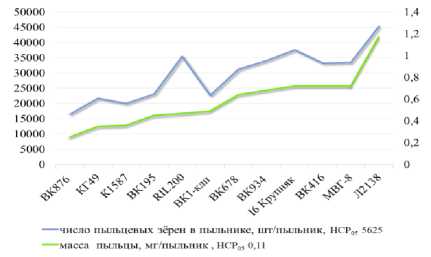
Рисунок 1 – Число и масса пыльцевых зёрен в пыльнике, 2023 г.
Наибольшая масса пыльцы выявлена у линии Л2138 (1,17 мг), наименьшая – у ВК876 (0,25 мг) при среднем значении 0,59 мг/пыльник и коэффициенте вариации 32 % (рис. 1). Между признаками числа и массы пыльцевых зёрен отмечена высокая положительная корреляция – 0,90.
В 2022 г. показатель диаметра пыльцевых зёрен у изученных линий варьировал от 30 мкм у КГ49 до 32 мкм у ВК195, К1587, ВК934, ВК1-клп, Л2138 и ВК416
при среднем значении 31 мкм. По данному признаку пять линий (ВК195, К1587, МВГ-8, ВК934 и ВК1-клп) имели невысокие значения коэффициента вариации – до 10 %. Семь линий: RIL200, I 5 Крупняк, Л2138, ВК678, ВК876, КГ49 и ВК416 характеризовались средним коэффициентом вариации (табл. 1).
Таблица 1
Диаметр пыльцевых зёрен у линий подсолнечника
2-е отделение, ЦЭБ ВНИИМК, г. Краснодар, 2022 г.
|
Генотип |
d, мкм |
CV, % |
||
|
среднее |
min |
max |
||
|
ВК195 |
32 |
10 |
35 |
9 |
|
К1587 |
32 |
10 |
35 |
8 |
|
МВГ-8 |
31 |
23 |
35 |
8 |
|
ВК934 |
32 |
25 |
38 |
8 |
|
RIL200 |
31 |
8 |
38 |
11 |
|
ВК1-клп |
32 |
10 |
38 |
10 |
|
I 5 Крупняк |
31 |
10 |
38 |
19 |
|
Л2138 |
32 |
13 |
38 |
13 |
|
ВК678 |
31 |
10 |
38 |
12 |
|
ВК876 |
31 |
10 |
38 |
16 |
|
КГ49 |
30 |
10 |
38 |
19 |
|
ВК416 |
32 |
10 |
40 |
13 |
НСР 05 1
Нормальные пыльцевые зёрна имели округлую форму и диаметр около 31 мкм. При этом в пыльце присутствовали зёрна неправильной формы и меньшего диаметра. Нетипичные пыльцевые зёрна можно классифицировать по размерам на два вида: среднеразмерные деформированные (диаметр 20–30 мкм) и микропыльцевые зёрна (диаметр 8–18 мкм) (рис. 2).
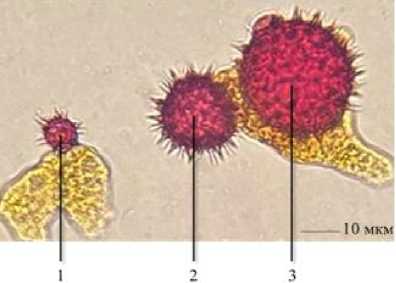
Рисунок 2 – Пыльцевые зёрна подсолнечника (1 – микропыльцевое зерно; 2 – среднеразмерное зерно; 3 – нормальное пыльцевое зерно), 2022 г.
Минимальное количество дефектных пыльцевых зёрен отмечено у линии ВК195 – 11 шт. (6 %), из них девять были среднеразмерными и два – микропыльце-выми. Максимальное количество дефектных пыльцевых зёрен имели две линии КГ49 и ВК416 – 89 шт. (45 %). У линии КГ49 из 89 деформированных пыльцевых зёрен 79 среднеразмерных и 10 микро-пыльцевых. Для линии ВК416 было отмечено 88 среднеразмерных и одно микропыльцевое зерно. У линий МВГ-8 и ВК934 микропыльцевые зёрна отсутствовали (табл. 2).
Таблица 2
Типы дефектных пыльцевых зёрен
2-е отделение, ЦЭБ ВНИИМК, г. Краснодар, 2022 г.
|
Генотип |
Дефектные пыльцевые зёрна |
|||||
|
всего |
среднеразмерные (диаметр 20–30 мкм) |
микропыльце-вые (диаметр 8–18 мкм) |
||||
|
штук |
% |
штук |
% |
штук |
% |
|
|
ВК195 |
11 |
6 |
9 |
82 |
2 |
18 |
|
К1587 |
15 |
8 |
14 |
97 |
1 |
7 |
|
МВГ-8 |
16 |
9 |
16 |
100 |
0 |
0 |
|
ВК934 |
20 |
10 |
20 |
100 |
0 |
0 |
|
RIL200 |
22 |
11 |
19 |
86 |
3 |
14 |
|
ВК1-клп |
28 |
14 |
27 |
96 |
1 |
4 |
|
I 5 Крупняк |
47 |
24 |
36 |
77 |
11 |
23 |
|
Л2138 |
55 |
26 |
50 |
91 |
5 |
9 |
|
ВК678 |
56 |
29 |
54 |
96 |
2 |
4 |
|
ВК876 |
60 |
30 |
54 |
90 |
6 |
10 |
|
КГ49 |
89 |
45 |
79 |
89 |
10 |
11 |
|
ВК416 |
89 |
45 |
88 |
99 |
1 |
1 |
НСР 05 4
В 2023 г. минимальное значение диаметра пыльцевых зёрен отмечено для линии RIL200 – 27 мкм, максимальный показатель был у ВК1-клп и Л2138 – 32 мкм при среднем значении 30 мкм. По данному признаку семь линий (ВК195, ВК934, Л2138, МВГ-8, К1587, ВК1-клп и ВК416) имели невысокие значения коэффициента вариации – до 10 %. Пять линий: RIL200, I 6 Крупняк, ВК678, ВК876 и КГ49, характеризовались средним коэффициентом вариации (табл. 3).
Линия ВК195 характеризовалась минимальным количеством дефектных пыльцевых зёрен – 8 шт. (3 %), все они были среднеразмерные. У линии КГ49 отмечено максимальное количество дефектных пыльцевых зёрен – 204 шт.
(68 %), из них 201 шт. (99 %) среднеразмерных и 3 шт. (1 %) микропыльцевых.
Таблица 3
Диаметр пыльцевых зёрен у линий подсолнечника
2-е отделение, ЦЭБ ВНИИМК, г. Краснодар, 2023 г.
|
Генотип |
d, мкм |
CV, % |
||
|
среднее |
min |
max |
||
|
ВК195 |
31 |
27 |
35 |
5 |
|
ВК934 |
30 |
24 |
33 |
5 |
|
МВГ-8 |
31 |
23 |
35 |
7 |
|
Л2138 |
32 |
24 |
37 |
8 |
|
I 6 Крупняк |
30 |
12 |
35 |
15 |
|
ВК1-клп |
32 |
25 |
37 |
8 |
|
К1587 |
30 |
25 |
33 |
7 |
|
ВК678 |
29 |
18 |
34 |
11 |
|
ВК876 |
28 |
14 |
33 |
14 |
|
ВК416 |
30 |
22 |
35 |
10 |
|
RIL200 |
27 |
17 |
33 |
11 |
|
КГ49 |
28 |
14 |
35 |
13 |
НСР 05 1
У линий Л2138, К1587 и ВК678 наблюдали дефектные крупные пыльцевые зёрна. Линии ВК934, Л2138, К1587, ВК1-клп не имели микропыльцевых зёрен (табл. 4).
Таблица 4
Типы дефектных пыльцевых зёрен
2-е отделение, ЦЭБ ВНИИМК, г. Краснодар, 2023 г.
|
Генотип |
Дефектные пыльцевые зёрна |
|||||||
|
всего |
среднеразмерные (диаметр 20–30 мкм) |
микро-пыльцевые (диаметр 8–18 мкм) |
крупные (диаметр 33–35 мкм) |
|||||
|
штук |
% |
штук |
% |
штук |
% |
штук |
% |
|
|
ВК195 |
8 |
3 |
8 |
100 |
0 |
0 |
0 |
0 |
|
ВК934 |
18 |
6 |
18 |
100 |
0 |
0 |
0 |
0 |
|
МВГ-8 |
23 |
8 |
22 |
96 |
1 |
4 |
0 |
0 |
|
Л2138 |
41 |
14 |
39 |
95 |
0 |
0 |
2 |
5 |
|
К1587 |
58 |
19 |
57 |
98 |
0 |
0 |
1 |
2 |
|
I 6 Крупняк |
61 |
20 |
47 |
77 |
14 |
23 |
0 |
0 |
|
ВК678 |
86 |
29 |
82 |
95 |
2 |
3 |
2 |
2 |
|
ВК876 |
88 |
29 |
71 |
81 |
17 |
19 |
0 |
0 |
|
ВК1-клп |
89 |
30 |
79 |
89 |
0 |
0 |
0 |
0 |
|
ВК416 |
102 |
34 |
101 |
99 |
1 |
1 |
0 |
0 |
|
RIL200 |
172 |
57 |
162 |
94 |
10 |
6 |
0 |
0 |
|
КГ49 |
204 |
68 |
201 |
99 |
3 |
1 |
0 |
0 |
НСР 05 5
За два года исследований линия ВК195 характеризовалась минимальной гетерогенностью пыльцы – 5 %, а линия КГ49 имела максимальные показатели данного признака – 58 % (рис. 3). Остальные линии показали долю дефектных пыльцевых зёрен в интервале от 9 до 42 %.
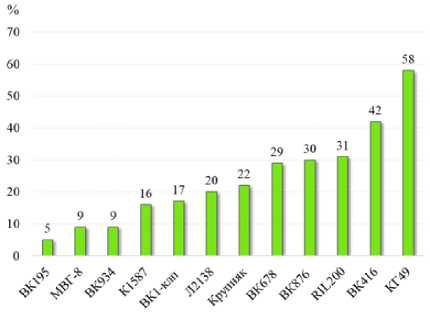
Рисунок 3 – Гетерогенность пыльцевых зёрен, 2022–2023 гг.
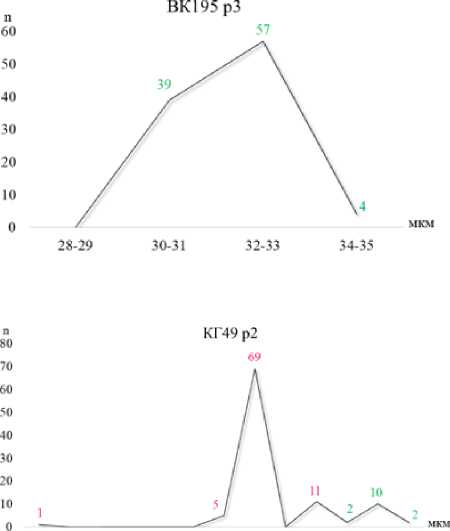
Для линии ВК195 (растение 3) на графике вариационной кривой отмечен один пик для диаметра 32–33 мкм, при этом гетерогенность пыльцы отсутствовала – 0 %, т. е. все пыльцевые зёрна были морфологически однородны, без деформаций (рис. 4). Кроме того, для данного растения отмечен высокий уровень автофертильности – 57 %.
Напротив, для линии КГ49 (растение 2) наблюдали значительную гетерогенность диаметра пыльцевых зёрен – 89 %, что сопровождается трехпиковым распределением значений признака. В частности, отмечен основной пик среднеразмерных деформированных пыльцевых зёрен (24–25 мкм) и малый пик (28– 29 мкм) также деформированных пыльцевых зёрен. Нормальные пыльцевые зёрна (32–33 мкм) образовали малый пик из 10 шт. (рис. 4). Автофертильность данного растения была очень низкой – 6 %.
10-H12-1314-1516-1718-1920-2122-2324-2526-2728-2930-3152-3334-35
Рисунок 4 – Вариационная кривая диаметра пыльцевых зёрен у подсолнечника (n = 100), 2023 г.
Выводы. В ходе исследований было установлено, что максимальной массой (1,17 мг/пыльник) и числом пыльцевых зёрен (45239 шт/пыльник) характеризовалась линия Л2138. Линия ВК876 имела минимальные значения данных признаков – 0,25 мг/пыльник и 16448 шт/пыльник соответственно. Между признаками числа и массы пыльцевых зёрен отмечена высокая положительная корреляция – r = 0,90. В 2022 г. средний показатель диаметра пыльцевых зёрен у 12 изученных ли ний варьировал от 30 до 32 мкм при среднем значении 31 мкм. В 2023 г. минимальный показатель диаметра пыльцевых зёрен был ниже, чем в 2022 г., – 27 мкм, максимальный находился на уровне 2022 г. – 32 мкм при среднем значении 30 мкм. За два года исследований у линии КГ49 отмечена значительная морфологическая гетерогенность пыльцы, что выражалось в высокой доле дефектных пыльцевых зёрен – 58 %. Линия ВК195 характеризовалась низкой морфологиче- ской гетерогенностью с долей дефектных пыльцевых зёрен 5 %.
Список литературы Пыльцевой анализ линий селекционно-генетической коллекции подсолнечника
- Киселёва Н.С. Оценка качества пыльцы генотипов груши для использования в селекции // Новые технологии. – 2019. – № 4 (50). – С. 153–165.
- Корзин В.В., Горина В.М., Месяц Н.В. Качество пыльцы сортов и форм абрикоса и перспективность ее использования в селекционном процессе // Вестник КрасГАУ. – 2019. – № 8. – С. 10–16.
- Цаценко Л.В., Назаров А.Л., Леденева А.Л. Пыльцевой анализ в селекции пшеницы // Научный журнал КубГАУ. – 2021. – № 165 (01). – С. 1–11.
- Chamer A.M., Medan D., Mantese A.I., Bartoloni N.J. Impact of pollination on sunflow-er yield: Is pollen amount or pollen quality what matters? // Field Crops Research. – 2015. – Vol. 176. – P. 61–70.
- Horner H.T. A comparative light and elec-tron microscopic study of microsporogenesis in male-fertile and cytoplasmic male-sterile Sunflower (Helianthus annuus) // Amer. J. Bot. – 1977. – No 64 (6). – P. 745–759.
- Устинова Е.И. Цитоэмбриологическое исследование подсолнечника и кукурузы: ав-тореферат дис. ... д-ра биол. наук / Елена Ивановна Устинова. – М., 1967. – 40 с.
- Gotelli M.M., Galati B.G., Medan D. Em-bryology of Helianthus annuus (Asteraceae) // Ann. Bot. Fennici. – 2008. – No 45. – P. 81–96.
- Бессонова В.П. Состояние пыльцы как показатель загрязнения среды тяжелыми ме-таллами // Экология. – 1994. – № 4. – С. 45–50.
- Rezanejad F., Maid A., Shariatzade S.M. Effect of air pollution on water proteins, struc-ture and cellular material release in Lagerstroe-mia indica L. (Lytraceae) // XVII Intern. Congr. «Sexual Plant Reproduction in Nature and the Laboratory»: Progr., Abstr. – Lublin, 2002. – P. 128.
- Lalonde, S., Beebe D.U., Saini H.S. Early signs of disruption of wheat anther development associated with the induction of male sterility by meiotic-stage water deficit // Sexual Plant Re-production. – 1997. – No 10. – Р. 40–48.
- Бабро А.А., Воронова О.Н. Развитие мужских репродуктивных структур у Helianthus ciliaris и H. tuberosus (Asteraceae) // Ботанический журнал. – 2018. – № 103 (9). – С. 1093–1108.
- Astiz V., Hernández L.F. Pollen produc-tion in sunflower (Helianthus annuus L.) is af-fected by air temperature and relative humidity during early reproductive growth // Fyton. – 2013. – No 82. – Р. 297–302.
- Kalyar T., Rauf S., Teixeira da Silva J.A., Shahzad M. Handling sunflower (Helianthus annuus L.) populations under heat stress // Ar-chives of Agronomy and Soil Science. – 2014. – Vol. 60. – P. 655–672.
- Мейер-Меликян Н.Р., Бовина И.Ю., Косенко Я.В. [и др.]. Атлас пыльцевых зёрен Астровых (Asteraceae) – М.: Товарищество научных изданий КМК, 2004. – 236 с.
- Воронова О.Н., Гаврилова В.А. Количественный и качественный анализ пыльцы подсолнечника (Helianthus L.) и его использование в селекционной работе // Труды по прикладной ботанике, генетике и селекции. – 2019. – Вып. 180 (1). – С. 95–104.
- Bonciu E. Aspects of the pollen grains di-ameter variability and the pollen viability to some sunflower genotypes // Journal of Horticul-ture, Forestry and Biotechnology. – 2013. – Vol. 17 (1). – P. 161–165.
- Andrei E., Jitareanu C.D. Selection of sunflower restorer inbred lines according to the pollen amount // Proc. of 15th Intern. Sunflower Conf., Toulouse, France, 2000. – V. 2. – P. 150–155.
- Vear F., Pham-Delegue M., Tourvieille de Labrouhe D. [et al.]. Genetical studies of nec-tar and pollen production in sunflower // Agron-omie. – 1990. – No 10. – Р. 219–231.
