Quorum Sensing и нодуляционная конкурентоспособность ризобий при инфицировании бобовых растений

Автор: Воробьев Н.И., Проворов Н.А.
Журнал: Сельскохозяйственная биология @agrobiology
Рубрика: Молекулярные и клеточные механизмы симбиоза
Статья в выпуске: 3 т.50, 2015 года.
Бесплатный доступ
Один из основных симбиотических признаков ризобий - способность конкурировать за образование клубеньков у бобовых растений, проявляемая при совместной инокуляции хозяев несколькими штаммами (нодуляционная конкурентоспособность - НКС). Закономерности регуляции НКС остаются малоизученными, что затрудняет использование полученных данных в биотехнологических исследованиях, направленных на повышение эффективности биопрепаратов ризобий. Нами создана математическая модель нодуляционной конкурентоспособности ризобий (НКС) для описания мультиштаммовой инокуляции бобовых растений (НКС-модель). В НКС-модели мы учли, что в соответствии с положением о Quorum Sensing регулировании (QS-регулировании) в почвенных нишах ограничивается не только число бактерий в нише, но и миграция клеток в соседние ниши. НКС-модель представляет собой нелинейную степенную зависимость миграции клеток из ниши от пространственной плотности клеток в нише (величина степени меньше, чем 1). Согласно формуле НКС-модели, относительная доля мигрирующих клеток из ниши обитания уменьшается с ростом плотности клеток в нише. Мы предполагаем, что после мультиштаммовой инокуляции в ризосфере растений в большем количестве (с большей НКС) будет представлен тот бактериальный штамм, который формирует больший миграционный поток клеток из ниши обитания. Верификация и параметрическая идентификация (миграционная активность штаммов, индекс частотно-зависимого отбора) НКС-модели проводились с использованием опубликованных ранее экспериментальных данных. Анализ вычисленных по этим данным параметров НКС-модели показал, что штаммы ризобий с большей миграционной активностью и малым индексом частотно-зависимого отбора образуют больший миграционный поток клеток из ниш обитания и демонстрируют большую НКС. Генетическая модификация ризобий с целью повышения НКС может вызвать интенсификацию миграционных потоков клеток штамма из ниши обитания и к ускоренному расходованию ресурсов ниши. Интенсивная миграция клеток из почвенной ниши может привести к преждевременному расходованию ресурса ниши и вызвать гибель клеток ризобий раньше, чем они образуют симбиоз с бобовыми растениями. Рассмотренные особенности выживания и миграции ризобий в естественных почвенных условиях необходимо учитывать, особенно, при разработке микробных препаратов длительного действия.
Математическое моделирование, клубеньковые бактерии, нодуляционная конкурентоспособность, регуляция quorum sensing, миграция, частотно-за-висимый отбор
Короткий адрес: https://sciup.org/142133590
IDR: 142133590 | УДК: 635.65:573.22 | DOI: 10.15389/agrobiology.2015.3.298rus
The Quorum Sensing and the nodulation competitiveness of rhizobia during infection of leguminous plants
A basic property of Rhizobia is the ability to compete for nodule formation under a multi strain inoculation. Its regulation remains poorly understood, thus limiting advances in biotechnological studies to improve the effectiveness of Rhizobia microbial preparations. The mathematical model of Rhizobia Nodulation Competitiveness (RNC) for describing a multi strains inoculation of leguminous plants is created (RNC-model). In the RNC-model we took into account that under Quorum Sensing regulation (QS-regulation) in soil niches not only the bacteria number in the niche, but a cell’s migration into adjacent niches is limited. The RNC-model represents a power-law nonlinear dependence of cell’s migration from a niche from the cell’s spatial density in the niche (the power value is less than 1). According to the formula of the RNC-model, the relative proportion of migratory cells from niche habitat decreases under increasing cell’s density. We suggest that after a multi strains inoculation in the rhizosphere of plants the bacterial strains with a large cells migration flow from the niche habitat (with big RNC) will be presented in large amounts. The verification and parametric identification (migration activity of strains, the index of frequency-dependent selection) of the RNC-model were carried out with using the previously published experimental data. The analysis of RNC-model’s parameters, calculated with using these data, showed that rizobia strains with a large migratory activity and a small index of frequency-dependent selection form a large migratory flow of cells from the niche habitat and demonstrate large RNC. The genetic modification of rhizobia with the aim of improving the RNC may cause the intensification of migration flows of strain cells from the niche habitat and the accelerated spending of niche resources. The intensive migration of cells from soil niche may lead to premature consumption of niche’s resource and cause a cell death of rhizobia before they form symbiosis with legume plants. The considered features of survival and migration of rhizobia in natural soil conditions must be considered, especially, in the development of microbial preparations with prolonged action.
Текст научной статьи Quorum Sensing и нодуляционная конкурентоспособность ризобий при инфицировании бобовых растений
Один из основных симбиотических признаков ризобий ― их способность конкурировать за образование клубеньков у бобовых растений, проявляемая при совместной инокуляции хозяев несколькими штаммами (нодуляционная конкурентоспособность ― НКС). Молекулярно-генетические исследования выявили полигенный контроль НКС, показали его связь с различными культурально-биохимическими и симбиотическими признаками (структурой клеточной стенки, устойчивостью к антибиотикам и фагам, способностью адсорбироваться на поверхности корней, скоростью образования клубеньков при моноинокуляции растений) и позволили приступить к разработке методов конструирования штаммов ризобий с повышенной НКС (1-5). Вместе с тем закономерности регуляции НКС остаются малоизученными, что затрудняет использование полученных данных в биотехнологических исследованиях, направленных на повышение эффективности биопрепаратов ризобий.
Ранее была предложена гипотеза о том, что скорость миграции бактерий между разными нишами, определяющая их конкурентоспособность, может регулироваться с участием механизма Quorum Sensing (QS-
∗ Работа поддержана Российским научным фондом, грант 14-26-00094.
регуляция) (6-8). На это указывают экспериментальные данные о колонизации ризобиями клубеньков растений (9, 10), демонстрирующие нелинейную зависимость НКС от общего числа (следовательно, и от пространственной плотности) бактерий в инокулюме. QS-регуляция осуществляется посредством низкомолекулярных сигнальных молекул (аутоиндукторов), которые выделяются бактериями во внешнюю среду и воздействуют на регуляторные белки, вызывая блокировку либо запуск клеточного деления (11-16). В результате популяция может разделиться на группы клеток с разным физиологическим состоянием, в которых процессы размножения либо активированы, либо заблокированы (17). Ранее такая дифференциация была показана для клубеньковых бактерий люцерны ( Si-norhizobium meliloti ), у которых QS-регуляция обеспечивает различие изогенных клеток по активности генов биосинтеза экзополисахаридов, определяющей скорость развития колонии (18).
Цель представляемого исследования ― построение математической модели нодуляционной конкурентоспособности бактериальных штаммов (НКС-модели), учитывающей QS-регуляцию миграционных потоков клеток из инокулюма в клубеньковые ниши бобовых растений.
Построение модели. Ранее было установлено (9), что отношение долей клубеньков, колонизированных двумя конкурирующими штаммами ( k 1: k 2) связано с отношением их численностей в единице объема инокулюма ( N 1: N 2) уравнением:
где k 1 + k 2 = 1; с12 > 0 ― парный индекс НКС, равный отношению числа клубеньков, колонизированных изученными штаммами, при их одинаковой представленности в инокулюме (с12 = k 1: k 2, N 1 = N 2); 0 < а < 1 ― индекс частотно-зависимого отбора (ЧЗО), определяющий селективные преимущества наиболее редкого штамма при с12 = 1. С уменьшением индекса ЧЗО представленность этого штамма в клубеньках возрастает (11).
Мы полагаем, что в ответ на повышение пространственной плотности каждого штамма QS-регуляция, поддерживая соотношение деля- щихся и покоящихся клеток, смещает его в пользу покоящихся клеток, что ведет к замедлению скорости роста штамма. При моделировании этого процесса мы исходили из того, что миграционные потоки клеток из ниши обитания подвергаются QS-регуляции и нелинейно зависят от пространственной плотности популяций. Эти допущения представляются логичными, поскольку делящиеся клетки расходуют значительно больше получаемых извне энергетических ресурсов, чем покоящиеся клетки, которым эти ресурсы нужны только на поддержание жизнеспособности. Очевидно, что для экономии ресурсов и повышения выживаемости бактериального штамма доля активных клеток в нем должна быть минимальной. Возможно, что QS-регуляция сформировалась в бактериальных популяциях для длительного выживания в условиях дефицита внешних ресурсов.
Интенсивность QS-регулируемого миграционного потока бактериального штамма из инокулюма определяет масштабы накопления клеток на поверхности корней. В свою очередь, различие в количествах клеток на поверхности корней предопределяет распределение штаммов в клубеньках при их колонизации. Следовательно, первопричиной НКС бактериальных штаммов служит QS-регуляция клеточных миграционных потоков. Поэтому при построении НКС-модели мы будем учитывать QS-регуляцию миграционных потоков клеток, функциональный характер которой пред- ставлен в уравнении [1]. Поскольку в ответ на возрастание пространственной плотности каждого штамма QS-регуляция снижает долю мигрирующих клеток, в НКС-модели следует использовать обратно пропорциональную зависимость миграционного индекса (m) от числа клеток каждого штамма:
n m = —M =
N
r
1+
V
N
1-a
N M ,
-1
где N ― суммарное число делящихся и покоящихся клеток штамма в единице объема инокулюма; nM < N — численность способных мигрировать клеток; NМ ― миграционная активность бактериального штам- ма, выраженная плотностью клеток в нише, при которой половина клеток способна мигрировать в соседние ниши (2-nM = N = Nm); 0 < а < 1 — индекс ЧЗО из уравнения [1].
При мультиштаммовой конкуренции для вычисления доли клу- беньков (kJ), колонизированных J-штаммом, необходимо определить долю клеток этого штамма (nMJ) среди мигрирующих клеток всех штаммов:
k J = n M J ^ n M L ’
L =1
где kJ ― доля клубеньков, колонизированных J-штаммом; nMJ, nML ― число клеток J- и L-штаммов в единице объема инокулюма, рассчитываемое по НКС-модели [2]; S ― число штаммов, участвующих в колонизации клубеньков. Для конкуренции между двумя штаммами из [3] можно полу- чить равенство:
k 1 : k 2 = n Ml : n M2 . [4]
Подставляя [2] в [4] и учитывая, что N 1 и N 2 принимаются значительно большими, чем NM1 и NM2, получим уравнение, асимптотически близкое к [1]:
al I ЛУа1 I log I /'1 = log I A' I = log М1 l + log 1 I, [5] Vk2 J VnA2 J V NМ-2 J V N2a2 J, 1 J где NM1, NM2 — миграционная активность популяций конкурирующих штаммов; 0 < а1, а2 < 1 — индексы ЧЗО для этих штаммов. Асимптотическая близость [1] и [5] доказывает корректность НКС-модели [2] и допускает использование [2] и [3] для описания мультиштаммовой колонизации корней бобовых растений. В результате для двух конкурирующих штаммов можно рассчитать индивидуальные (с1 = N 1M- a11, с2 = N M1-2a2) и парный (c12 = c1:c2) индексы НКС.
Верификация модели. Используя экспериментальные данные (9, 10), мы определили уровни миграционной активности популяций, а также индивидуальные индексы НКС и ЧЗО для конкурирующих штаммов ризобий (табл.). В качестве показателя адекватности НКС-модели использовался коэффициент детерминации (r2) экспериментальных и расчетных (вычисленных по модели) данных. Близость коэффициента детерминации к единице подтверждает высокую сходимость НКС-модели и экспериментальных данных (см. табл., рис.).
Аналитические зависимости, построенные с помощью НКС-модели для значений параметров из таблицы, показали, что с возрастанием плотности бактерий в инокулюме число мигрирующих клеток растет (см. рис., А, сплошные линии), однако доля мигрирующих клеток (миграционный индекс) уменьшается (см. рис., А, пунктирные линии). Для штамма KIM5s
(см. рис., Б) при плотностях клеток в инокулюме 108 кл/мл и 1010 кл/мл QS-регуляция потока мигрирующих клеток определяется значениями соответственно 2600 и 9300 кл/мл.
Параметры НКС - модели колонизации клубеньков бобовых растений штаммами Rhizobium leguminosarum
|
Штамм ризобий |
Миграционная активность бактерий NM, кл/мл |
Индивидуальный индекс НКС, сM |
Индекс ЧЗО, аM |
Коэффициент детерминации модели, r2 |
|
FSS1 (9) |
10±1 |
3,8±0,4 |
0,42±0,03 |
0,98 |
|
FH20S1 (9) |
9±1 |
4,2±0,5 |
0,35±0,03 |
0,93 |
|
FS163S (9) |
12±1 |
4,0±0,5 |
0,45±0,03 |
0,98 |
|
KIM5s (10) |
42±2 |
14,8±1,6 |
0,28±0,02 |
0,98 |
|
CE3 (10) |
6±1 |
3,5±0,6 |
0,30±0,02 |
0,98 |
|
Примечание. НКС — нодуляционная конкурентоспособность, ЧЗО — частотно-зависимый отбор. |
||||
У родственных штаммов KIM5s и CE3 (см. табл.) индексы ЧЗО близки по величине, и поэтому в уравнении [5] можно принять их равенство a1 = a2. В этом случае [5] совпадает с [1], что подтверждает корректность НКС-модели [2]. Если инокулюм штамма FSS1 (см. табл.) содержит 107 кл/мл, а штамма FH20S1 — 105 кл/мл ( N 1: N 2 = 100:1), то в клубеньках эти штаммы будут представлены в соотношении k 1: k 2 = 93:7, то есть в клубеньках представленность редкого штамма FH20S1 возрастает в 7 раз. Дальнейшее снижение индексов ЧЗО (a1 = 0,21, a2 = 0,175) еще более изменит соотношение клубеньков в пользу штамма FSS1 ( k 1: k 2 = 80:20).
Как видно из представленных в таблице данных, штамм KIM5s имеет наибольший индивидуальный индекс НКС (с4 = 14,8). Можно предположить, что QS-регуляция численности этого штамма поддерживает б о льшую интенсивность потока мигрирующих клеток по сравнению с таковой у штамма CE3 (с5 = 3,5). Поэтому при равном соотношении штаммов KIM5s и CE3 в инокулюме в клубеньках будет преимущественно представлен штамм KIM5s: k 4: k 5 = с45 = с4:с5 = 14,8:3,5 = 4,2 > 1. Таким образом, повышенный индивидуальный индекс НКС и уровень миграционной активности могут быть объяснены ослабленной QS-регуляцией миграционного потока бактерий из инокулюма.
Анализ выявленных закономерностей позволяет предположить, что бактериальные штаммы с высокими значениями индекса миграционной активности и индивидуального индекса НКС тратят на миграцию клеток большое количество энергетических ресурсов, что сокращает время выживания бактерий. Поэтому повышение НКС у штаммов с помощью генетических методов может повлечь за собой снижение жизнеспособности ри-зобий вне растений и их последующую гибель.
Известно, что низкой выживаемостью вне растений часто обладают штаммы ризобий с повышенной эффективностью симбиотической N2-фиксации (19). Поэтому ЧЗО в пользу редких штаммов может играть важную роль в поддержании эффективного взаимодействия бактерий и растений. Благодаря этому отбору в клубеньках осуществляется преимущественное размножение активных азотфиксаторов, обеспечивающее максимальную продуктивность бобово-ризобиальной системы.
Ранее при изучении фитопаразитарных систем было показано, что растения могут влиять на QS-регуляцию генов вирулентности патогенных бактерий, подавляя инфекционный процесс (20). Возможно, что аналогичный механизм действует и при образовании бобово-ризобиального симбиоза, что позволяет растению регулировать потоки бактерий из окружающей среды в эндосимбиотические ниши и таким образом осуществлять отбор штаммов ризобий по признакам симбиотической эффективности. Проверка этой гипотезы входит в задачи нашей дальнейшей работы.
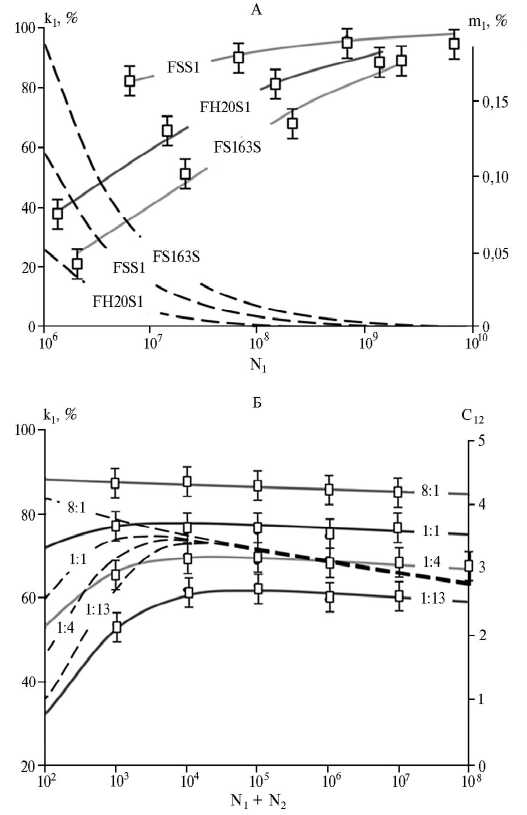
Распределение конкурирующих штаммов ризобий по клубенькам бобовых растений в зависимости от плотности клеток в инокулюмах (N1, N2): k 1 ― доля клубеньков, образованных штаммом FSS1 (9). Левые оси соответствуют экспериментальным данным ( ^ ) и сплошным линиям, построенным по НКС-модели [2], правые оси ― пунктирным линиям, которые соответствуют значениям миграционного индекса m 1 (А) и парного индекса НКС с12 (Б), вычисленным по НКС-модели для штамма FSS1.
А. Результаты конкуренции штаммов Rhizo-bium leguminosarum bv. vi-ciae (FSS1, FH20S1, FS163S) с местной почвенной популяцией за нодуляцию конских бобов Vicia faba сорта Ascott (9).
Б. Результаты конкуренции мутантного штамма KIM5s с родительским штаммом CE3 R. legumino-sarum bv. phaseoli за ноду-ляцию фасоли обыкновенной Phaseolus vul garis сорта Puebla 152 при разных соотношениях KIM5s:CE3 в инокулюме (10).
Итак, нодуляционная конкурентоспособность (НКС) ризобий определяется не только преимуществами штаммов при освоении питательных ресурсов в ризосфере бобовых растений, но и миграционной активностью клеток бактерий из ниш обитания. Повышение НКС методами генной инженерии может приводить к возрастанию миграционной активности клеток ризобий и преждевременному расходованию ресурсов почвенной ниши, в которой популяция ризобий обитает вне симбиоза. Поэтому при длительном выживании бактерий в почвенных условиях повышение энергетических затрат на миграцию клеток может повысить риски гибели популяции. Особенности выживания и миграции ризобий необходимо учитывать, в частности, при разработке микробных препаратов длительного действия.
Список литературы Quorum Sensing и нодуляционная конкурентоспособность ризобий при инфицировании бобовых растений
- Курчак О.Н., Проворов Н.А., Симаров Б.В. Плазмида pSym1-32 Rhizobium leguminosarum bv.viceae, контролирующая азотфиксирующую активность, эффективность симбиоза, конкурентоспособность и кислотоустойчивость. Генетика, 2001, 37(9): 1225-1232.
- Онищук О.П., Курчак О.Н., Шарыпова Л.A., Проворов Н.А., Симаров Б.В. Анализ различных типов конкурентоспособности у Тп5-мутантов клубеньковых бактерий люцерны (Sinorhizobium meliloti). Генетика, 2001, 37(11): 1507-1512.
- Онищук О.П., Симаров Б.В. Гены, контролирующие нодуляционную конкурентоспособность клубеньковых бактерий. Генетика 1996, 32: 1157-1166.
- Онищук О.П., Симаров Б.В. Генетическая изменчивость нодуляционной конкурентоспособности у клубеньковых бактерий и ее использование в селекции. Генетика, 1995, 31(3): 293-303.
- Sessitsch A., Jjemba P.K., Hardarson G., Akkermans A.D.L., Wilson K.J. Measurement of the competitiveness index of Rhisobium tropici strain CIAT899 derivatives marked with the gusA gene. Soil Biol. Biochem., 1997, 29: 1099-1110.
- Pérez-Montaño F., Jiménez-Guerrero I., Del Cerro P., Baena-Ropero I., López-Baena F.J., Ollero F.J., Bellogín R., Lloret J., Espuny R. The symbiotic biofilm of Sinorhizobium fredii SMH12, necessary for successful colonization and symbiosis of Glycine max cv. Osumi, is regulated by Quorum Sensing systems and inducing flavonoids via NodD1. PLoS ONE, 2014, 9(8): e105901 ( ) DOI: 10.1371/journal.pone.0105901
- Jitacksorn S., Sadowsky M.J. Nodulation gene regulation and Quorum Sensing control density-dependent suppression and restriction of nodulation in the Bradyrhizobium japonicum-soybean symbiosis. Appl. Environ. Microbiol., 2008, 74: 3749-3756 ( ) DOI: 10.1128/AEM.02939-07
- Zheng H., Zhong Z., Lai X., Chen W.X., Li S., Zhu J. LuxR/LuxI-type Quorum-Sensing system in plant bacterium Mesorhizobium tianshanense controls symbiotic nodulation. J. Bacteriol., 2006, 188: 1943-1949 ( ) DOI: 10.1128/JB.188.5.1943-1949.2006
- Amarger N., Lobreau J.P. Quantitative study of nodulation competitiveness in Rhizobium strains. Appl. Environ. Microbiol., 1982, 44(3): 583-588.
- Beattie G.A., Clayton M.C., Handelsman J. Quantitative Comparison of the laboratory and field competitiveness of Rhizobium leguminosarum biovar phaseoli. Appl. Environ. Microbiol., 1989, 55(11): 2755-2761.
- Проворов Н.А., Воробьев Н.И. Генетические основы эволюции растительно-микробного симбиоза. СПб, 2012.
- Хмель И.А. Quorum-Sensing регуляция экспрессии генов: фундаментальные и прикладные аспекты, роль в коммуникации бактерий. Микробиология, 2006, 75(4): 457-464.
- Шпаков А.О. Сигнальные молекулы бактерий непептидной природы QS-типа. Микробиология, 2009, 78(2): 163-175.
- Conway B., Greenberg E. Quorum-Sensing signals and Quorum-Sensing genes in Burkholderia vietnamiensis. J. Bacteriol., 2002, 184: 1187-1191 ( ) DOI: 10.1128/jb.184.4.1187-1191.2002
- Daniels R., De Vos D.E., Desair J., Raedschelders G., Luyten E., Rosemeyer V., Verreth C., Schoeters E., Vanderleyden J., Michiels J. The cin Quorum-Sensing locus of Rhizobium etli CNPAF 512 affects growth and symbiotic nitrogen fixation. J. Biol. Chem., 2002, 277: 462-468 ( ) DOI: 10.1074/jbc.M106655200
- Koutsoudis M.D., Tsaltas D., Minogue T.D., Von Bodman S.B. Quorum-Sensing regulation governs bacterial adhesion, biofilm development, and host colonization in Pantoea stewartii subspecies stewartii. PNAS, 2006, 103: 5983-5988 ( ) DOI: 10.1073/pnas.0509860103
- Воробьев Н.И., Семенов A.M., Шаталов А.А., Ван Бругген A.Х.К., Свиридова О.В. Нелинейное моделирование автоколебаний численности Pseudomonas fluorescens в субстрат-бактериальной системе. Изв. Самарского НЦ РАН, 2009, 11(7): 1620-1624.
- Schluter J.P., Czuppon P., Schauer O., Pfaffelhuber P., McIntosh M., Becker A. Classification of phenotypic subpopulations in isogenic bacterial cultures by triple promoter probing at single cell level. J. Biotechnol., 2015, 198: 3-14 ( ) DOI: 10.1016/j.jbiotec.2015.01.021
- Проворов Н.А., Онищук О.П., Юргель С.Н., Курчак О.Н., Чижевская Е.П., Воробьев Н.И., Затовская Т.В., Симаров Б.В. Конструирование высокоэффективных симбиотических штаммов бактерий: эволюционные модели и генетические подходы. Генетика, 2014, 50(11): 1273-1285.
- Braeken K., Daniels R., Ndayizeye M., Vanderleyden J., Michiels J. Quorum Sensing in bacteria-plant interactions. In: Molecular mechanisms of plant and microbe coexistence. Berlin, Heidelberg, Springer-Verlag, 2008: 265-289 (ISBN 978-3-540-75574-6) ( ) DOI: 10.1007/978-3-540-75575-3
