Радиоуглеродное датирование костных остатков сурка из пещеры Страшной (Северо-Западный Алтай)
")
Автор: Кривошапкин А.И., Сердюк Н.В., Зенин В.Н., Панов В.С., Васильев С.К., Пархомчук В.В., Рудая Н.А., Шалагина А.В.
Журнал: Проблемы археологии, этнографии, антропологии Сибири и сопредельных территорий @paeas
Рубрика: Археология каменного века палеоэкология
Статья в выпуске: XXII, 2016 года.
Бесплатный доступ
В рамках современного этапа археологических исследований в пещере Страшной (Северо-Западный Алтай) были взяты образцы для проведения радиоуглеродного датирования костей сурка из всех подразделений стратиграфического разреза пещеры. В результате датирования было получено 21 определение радиоуглеродного возраста остатков. Большая часть дат соотносится с периодом МИС 2 (сартанский стадиал) и попадает во временной интервал 17-19 тыс. л.н., остальные даты соотносятся с МИС 3 (каргинский интерстадиал). Ископаемая фауна и данные палинологического анализа подтверждают существование благоприятных условий для жизнедеятельности популяций сурка во время формирования верхней пачки позднеплейстоценовых отложений пещеры. В это время в окрестностях пещеры Страшной преобладали горные степи, полупустыни, лесотундры, каменистые россыпи с доминированием открытых биотопов. Проведенные исследования позволяют соотносить массовое присутствие костей сурка в отложениях пещеры именно с этим периодом.
Северо-западный алтай, радиоуглеродный метод датирования, поздний плейстоцен, сурок, морские изотопные стадии
Короткий адрес: https://sciup.org/14522325
IDR: 14522325 | УДК: 902.652:(571)
Radiocarbon dating of the marmot fossil remains from Strashnaya cave (Northwest Altai)
During the current phase of archaeological research at the Strashnaya cave (North-Western Altai) marmot bones samples from all stratigraphic layers of the cave were taken for radiocarbon dating. As a result, 21 radiocarbon dates were obtained. Most of the dates correlate with MIS 2, and fit the time interval 17-19 kyr BP. The other dates correspond to the MIS 3. The paleontological and palynological data confirm the existence of favorable conditions for the marmot populations during the formation of top part of Late Pleistocene sediments. At this time, mountain steppes, semi-desert, forest-tundra landscapes in the vicinity of the cave dominated. It allows us to correlate the presence of numerous marmot bones in cave deposits with this period.
Текст научной статьи Радиоуглеродное датирование костных остатков сурка из пещеры Страшной (Северо-Западный Алтай)
Пещера Страшная – многослойный археологический объект, расположенный в Краснощековском р-не Алтайского края (Северо-Западный Алтай). Многолетние исследования, проводимые на объекте специалистами из Института археологии и этнографии СО РАН, показали, что отложения пещеры подразделяются на 13 литологических слоев, большинство из которых содержит богатый археологический и палеонтологический материал. Исключение составляют слои 11, 12 и 13, в которых находок не обнаружено, весь найденный материал из этих слоев происходят из кротовин грызунов. Проведенное ранее радиоуглеродное датирование, выполненное по образцам костей и угля из слоев 3, 5 и 6, показало, что хронологические рамки накопления данных слоев укладываются в интервал от 19 до 40 тысяч л.н., хотя некоторые из этих дат оказались запредельными для радиоглеродного метода [Криво-шапкин, Зенин, Шалагина, 2014].
Более 40 % определимого фаунистического материала пещеры принадлежит суркам, чьи остатки и следы жизнедеятельности (глубокие кротовины) прослеживаются во всех слоях. Численность остатков сурка постепенно убывает по направлению от предвходовой площадки в сторону центральной камеры. Ископаемые сурки обнаружены не только в виде разрозненных частей скелета, но и в виде целых костяков, что может свидетельствовать о гибели животных во время анабиоза, а следовательно и об использовании сурками пещеры под гнездовые норы. До настоящего момента специальных исследований (генетических, морфометрических) по видовой диагностике ископаемого сурка из Страшной пещеры не проводилось, хотя в фаунистических списках он фигурировал как серый сурок Marmota baibacina (Kastschenko, 1899) [Васильев, Зенин, 2009].
Последние исследования современных популяций сурков Старого и Нового Света показали генетическое однообразие палеарктической ассоциации этих наземных беличьих [Брандлер, Ляпунова, Банникова и др., 2010, Steppan, Akhverdyan, Lyapunova et al., 1999]. Несмотря на это, внутри группы bobak близкие виды M. bobak (Müller, 1776), M. kastschenkoi (Stroganov et Yudin, 1956) и M. baibacina имеют достоверные морфологические отличия в строении черепа, нижней челюсти и посткраниального скелета [Гасилин, Косинцев, 2011; Громов, Бибиков, Калабухов и др., 1965; Громов, Ерабаева, 1995; Огнев, 1947; Потапова, Пузачен-ко, 1998; Слудский, Варшавский, Исмагилов и др., 1969; Тараненко, 2005, 2011; Cardini, Slice, 2004]. Следовательно, для понимания этапов формирования древней популяции сурков необходима их видовая идентификация. Учитывая прежние фауни- стические наработки в этой области по материалам пещеры и комплекс морфологических характеристик черепа, обнаруженного при полевых работах в 2016 г., на данном этапе исследований ископаемых сурков Страшной пещеры следует относить к алтайскому (или серому) сурку. Последующее углубленное изучение позволит уточнить детали систематики и отследить становление вида.
С целью выяснения, в какой период времени обитала колония сурков в пещере Страшная и насколько сильно могут быть повреждены стратиграфические подразделения пещеры, было проведено радиоуглеродное датирование по костям сурков. Чтобы исключить повторное датирование одного и того же индивида были использованы только тазовые кости.
Навески костной ткани по 500–700 мг обрабатывались по стандартной ABA-методике [Brock, Higham, Ditchfield et al., 2010], выделенные индивидуальные образцы коллагена после высушивания были количественно переведены в углекислый газ с помощью системы EuroEA 3028 HT и далее в образцы графита по модифицированной методике [Bronk Ramsey, Humm, 2000]. Остаточная концентрация радиоуглерода определялась с помощью экспериментальной установки УМС СО РАН.
В результате датирования была получена 21 радиоуглеродная дата (см. таблицу ). В слоях 1–6 из 15 дат 13 укладываются в возрастной диапазон 17–20 тыс. л.н. Согласно шкале SPECMAP [Imbrie, Hays, Martinson et al., 1984] и уточненной шкале по стеку из низкоширотных океанических скважин ODP 677+MD900963 [Bassinot, Labeyrie, Vincent et al., 1994] этот период относится к МИС 2 (сар-танский стадиал). В слоях 7–12 из 11 дат 7 располагаются в диапазоне 41,5–52 тыс. л.н. (большинство из них, по-видимому, запредельные), что соответствует МИС 3 (каргинский интерстадиал). Две из дат приходятся на середину сартанского стадиала, две – на заключительную треть каргинского времени (см. рисунок ).
Проведенное исследование показало, что пик норной деятельности сурков пришелся на середину сартанского времени. Их кротовины уходили на глубину свыше 3 м. Датировка > 50 тыс. л.н из слоя 3.3 показывает, что сурки поднимали на дневную поверхность все включения в грунте, мешающие работе, в том числе и кости своих предшественников.
Современные сурки предпочитают открытые пространства с разреженным травостоем; основу их питания составляют сочные растительные корма, под норы грызуны стараются использовать рыхлые мелкоземы. Результаты изучения фауны современных мелких млекопитающих указывают 101
Радиоуглеродные датировки по тазовой кости сурка Marmota baibacina из Страшной пещеры
|
Номер |
Код |
Возраст |
Слои пещеры |
|
NSKA 01010 |
109-С14 |
1698±466,12 |
Сл. 1 |
|
NSKA 01011 |
110-С14 |
20097±674,11 |
Сл. 2 |
|
NSKA 01012 |
111-С14 |
18415±742,64 |
Сл. 3.1.А |
|
NSKA 01013 |
112-С14 |
17394±505,03 |
Сл. 3.1.Б |
|
NSKA 01014 |
113-С14 |
50000±0 |
Сл. 3.3 |
|
NSKA 01016 |
115-С14 |
19170±512,16 |
Сл. 4 |
|
NSKA 01017 |
116-С14 |
19078±483,53 |
Сл. 5 |
|
NSKA 01018 |
117-С14 |
18924±378,26 |
Сл. 5.1 |
|
NSKA 01019 |
118-С14 |
18216±374,54 |
Сл. 5.2 |
|
NSKA 01020 |
119-С14 |
18084±386,93 |
Сл. 5.3 |
|
NSKA 01021 |
120-С14 |
17323±607,23 |
Сл. 6 |
|
NSKA 01022 |
121-С14 |
18625±294,7 |
Сл. 6.1 |
|
NSKA 01023 |
122-С14 |
17984±346,32 |
Сл. 6.2 |
|
NSKA 01024 |
123-С14 |
25561±940,18 |
Сл. 6.3 |
|
NSKA 01025 |
124-С14 |
17134±431,23 |
Сл. 6.4 |
|
NSKA 01028 |
127-С14 |
32556±629,19 |
Сл. 7.1 |
|
NSKA 01029 |
128-С14 |
41468±2074,1 |
Сл. 7.2 |
|
NSKA 01032 |
131-С14 |
18426±264,69 |
Сл. 9 |
|
NSKA 01033 |
132-С14 |
24188±396,72 |
Сл. 10.1 |
|
NSKA 01034 |
133-С14 |
33097±892,55 |
Сл. 10.2 |
|
NSKA 01037 |
136-С14 |
52007±1893,4 |
Сл. 11.2 |
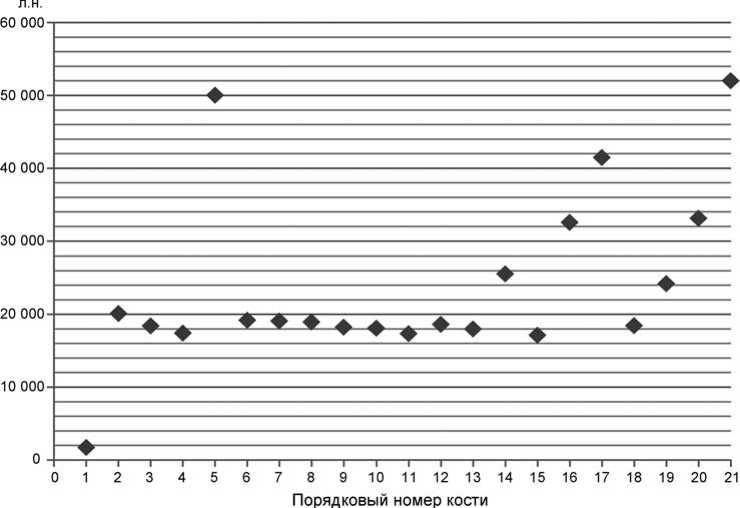
Радиоуглеродный возраст тазовых костей сурка Marmota baibacina из пещеры Страшной.
на преобладание таежного растительного комплекса в окрестностях Страшной пещеры в настоящее время [Пожидаева, 2005; Serdyuk, Zenin, 2016]. Ископаемая же фауна мелких млекопитающих демонстрирует яркую мозаичность ландшафтов во времена позднего плейстоцена в районе пещеры: горные степи, полупустыни, лесотундры, каменистые 102
россыпи с доминированием открытых биотопов [Serdyuk, Zenin, 2016]. Существование степных открытых фитоценозов во время накопления позднеплейстоценовых слоев подтверждается и палинологическими данными. В палинологических спектрах доминируют астровые, полыни и маревые [Рудая, Кривошапкин, Шалагина, 2016, в печати]. В таких условиях существование популяции сурков было успешным и продолжительным, что необходимо учитывать при последующих археологических работах в пещере.
Список литературы Радиоуглеродное датирование костных остатков сурка из пещеры Страшной (Северо-Западный Алтай)
- Брандлер О.В., Ляпунова Е.А., Банникова А.А., Крамеров Д. А. Филогения и систематика сурков (Marmota, Sciuridae, Rodentia), основанные на данных интер^ШЕ-ПЦР//Генетика. -2010. -Т. 46, № 3. -С. 321-331.
- Васильев С.К., Зенин А.Н. Фаунистические остатки из пещеры Страшная (Северо-Западный Алтай) по материалам раскопок в 1988-2008 годах//Проблемы археологии, этнографии, антропологии Сибири и сопредельных территорий. -Новосибирск: Изд-во ИАЭТ СО РАН, 2009. -Т. XV. -С. 56-62.
- Гасилин В.В., Косинцев П.А. Видовая диагностика степного (Marmota bobak) и серого (Marmota baibacina) сурков по краниологическим признакам//Зоол. журн. -2011. -Т. 90, № 12. -С. 1509-1521.
- Громов И.М., Бибиков Д.И., Калабухов Н.И., Мейер М.Н. Наземные беличьи (Marmotinae). -М.; Л.: Наука, 1965. -467 с. -(Фауна СССР. Млекопитающие; т. 3, вып. 2).
- Громов И.М., Ербаева М. А. Зайцеобразные и грызуны. Млекопитающие фауны России и сопредельных территорий. -СПб.: Изд-во СПб. гос. ун-та, 1995. -524 с.
- Кривошапкин А.И., Зенин В.Н., Шалагина А.В. Результаты полевых исследований пещеры Страшная в 2014 году//Проблемы археологии, этнографии, антропологии Сибири и сопредельных территорий. Новосибирск: Изд-во ИАЭТ СО РАН, 2014. -Т. XX. -С. 54-56.
- Огнев С.И. Звери СССР и прилежащих стран: звери Восточной Европы и Средней Азии. -М.; Л.: Изд-во АН СССР, 1947. -Т. 5. -810 с.
- Пожидаева Л.В. Население мелких млекопитающих некоторых биотопов территории Тигирекского заповедника//Горные экосистемы Южной Сибири: изучение, охрана и рациональное природопользование: мат-лы междунар. конф., посвящ. 5-летию организации Тигирекского заповедника. -Барнаул: Алтайские страницы, 2005. -С. 48-52.
- Потапова Е.Г., Пузаченко А.Ю. Морфологическая дифференциация серого сурка (Marmota baibacina) и тарбагана (M. sibirica) в Южной Монголии//Зоол. журн. -1998. -Т. 77, № 10. -С. 1177-1190.
- Рудая Н.А., Кривошапкин А.И., Шалагина А.В. Итоги палинологического изучения пещеры Страшная (Алтайский край) в 2014-2015 годах//Проблемы археологии, этнографии, антропологии Сибири и сопредельных территорий. -Новосибирск: Изд-во ИАЭТ СО РАН, 2016. -Т. XXII (в печати).
- Слудский А. А., Варшавский С.Н., Исмагилов М.И., Капитонов В.И., Шубин И.Г. Млекопитающие Казахстана. -Алма-Ата: Наука, 1969. -Т. 1. -456 с.
- Тараненко Д.Е. Сравнительный анализ морфологической изменчивости и эколого-географических характеристик лесостепного (Marmota kastschenkoi Stroganov et Yudin, 1956) и серого (Marmota baibacina Kastschenko, 1899) сурков: автореф. дис.... канд. биол. наук. -Новосибирск, 2005. -35 с.
- Тараненко Д.Е. Пространственные взаимоотношения двух видов сурков: Marmota kastschenkoi и Marmota baibacina (Rodentia, Sciuridae)//Журн. Сиб. федер. ун-та. Биология. -2011. -Т. 4, № 3. -С. 220-228.
- Bassinot F.C., Labeyrie L.D., Vincent E., Quidelleur X., Shackleton N.J., Lancelot Y. The astronomical theory of climate and the age of the Brunhes-Matuyama magnetic reversal//Earth and Planetary Science Letters. -1994. -Vol. 126 (1/3). -Р. 91-108.
- Brock F., Higham T., Ditch field P., Bronk Ramsey C. Current pretreatment methods for AMS Radiocarbon dating at the Oxford Radiocarbon Accelerator Unit (ORAU)//Radiocarbon. -2010. -Vol. 1. -P. 103-112.
- Bronk Ramsey C., Humm M.J. On-line combustion of samples for AMS and ion source developments at ORAU//Nuclear Instruments and Methods in Physics Research. Section B. -2000. -Vol. 172, iss. 1/4. -P. 242-246.
- Cardini A., Slice D.E. Mandibular shape in the genus Marmota (Rodentia, Sciuridae): a preliminary analysis using outlines//Ital. J. Zool. -2004. -Vol. 71. -P. 17-25.
- Imbrie J., Hays J.D., Martinson D.G., McIntyre A., Mix A.C., Morley J.J., Pisias N.G., Prell W.L., Shackleton N.J. The orbital theory of Pleistocene climate: support from a revised chronology of the marine δ18O record//Milankovitch and climate. Pt. 1. NATO ASI Series C. Mathematical and Physical Sciences. -1984. -Vol. 126. -P. 269-305.
- Serdyuk N., Zenin A. Small mammals from the Strashnaya cave (Northwest Altai, West Siberia, Russia)//Quaternary Intern. -2016. -Vol. 406, pt. B. -P. 162-168.
- Steppan S.C., Akhverdyan M.R., Lyapunova E.A., Fraser D.G., Vorontsov N.N., Hoffmann R.S., Braun M.J. Molecular phylogeny of the marmots (Rodentia, Sciuridae): tests of evolutionary and biogeographic hypotheses//Syst. Biol. -1999. -Vol. 48. -P. 715-734.
