Распространение и вирулентность заразихи (Orobanche cumana Wallr.) на подсолнечнике в Ростовской области
 на подсолнечнике в Ростовской области")
Автор: Антонова Т.С., Ситало Г.М., Арасланова Н.М., Гучетль С.З., Рамазанова С.А., Челюстникова Т.А.
Статья в выпуске: 1 (140), 2009 года.
Бесплатный доступ
Обследованы посевы подсолнечника в 43 районах Ростовской области на предмет выявления степени засоренности их заразихой (Orobanche cumana Wallr.). Лишь в 6 районах не обнаружена заразиха на полях подсолнечника. Остальные имели разную степень засоренности, в том числе, 14 районов - сильную и среднюю степень. Выполнена оценка вирулентности 18 популяций заразихи из районов Ростовской области и других регионов по отношению к европейским линиям дифференциаторам подсолнечника: 7, 202, LC1002, LC1003, LC1093, 16А×25, и P 96. Имеются популяции, где ещѐ преобладает раса D (в Сальском, Целинском, Шолоховском, Родионовонесветайском районах Ростовской области и Ейском Краснодарского края). Выявлена раса Е (в районах Константиновском, Цимлянском Ростовской области и Каневском Краснодарского края). Широко представлена раса F (в Константиновском, Миллеровском, Морозовском, 154 Цимлянском, районах Ростовской области, а также в обеих Волгоградских популяциях и Светлоградской Ставропольского края). Особенно неблагополучны районы Ростовской области: Белокалитвинский, Егорлыкский, Константиновский и Тацинский наличием высокой доли биотипа, преодолевающего устойчивость дифференциатора к расе G, что не позволяет однозначно указать на присутствие там и других рас. Этот биотип присутствует и в других изученных популяциях, за исключением Сальской и Целинской из Ростовской области.
Подсолнечник, линии-дифференциаторы, заразиха, расы, вирулентность, районы, степень засорения
Короткий адрес: https://sciup.org/142150880
IDR: 142150880 | УДК: 632.9:633.854.78
Spreading and virulence of broomrape in Rostov region
The sunflower crops in 43 districts of Rostov region were inspected for the purpose of exposure of their weeding degree by broomrape (Orobanche cumana Wallr.). It was shown 14 districts have a high and a mid degree of fields weeding by broomrape. It was carried out the test of virulence 18 broomrape populations from Rostov region and some other areas with respect to sunflower line differentiator: 7, 202, LC1002, LC1003, LC1093, 16Ах25, и P 96. There are populations where race D prevails (in districts Salsky, Tselinsky, Sholochovsky, Rodionovonesvetaisky from Rostov region and Eisky - Krasnodar region). It was revealed race E (in districts Konstantinovsky, Tsimlyansky from Rostov region and Kanevskoy - Krasnodar region). Widely race F is presented (in districts Konstantinovsky, Millerovsky, Morozovsky, Tsimlyansky from Rostov region, in both populations from Volgograd region and Svetlogradskaya - Stavropol region). Especially districts Belokalitvinsky, Egorliksky, Konstantinovsky, Tatsinsky from Rostov region are not satisfactory owing to the presence a big portion of biotype overcoming the differential resistance to race G. This fact is not let to show there the presence of other races. This biotype presents in other studied populations except ones from Salsky and Tselinsky districts of Rostov region.
Текст научной статьи Распространение и вирулентность заразихи (Orobanche cumana Wallr.) на подсолнечнике в Ростовской области
Введение. Облигатный паразит подсолнечника Orobanche cumana Wallr. относится к высшим цветковым растениям и способен погубить весь урожай. До введения подсолнечника в культуру этот вид заразихи был распространён в девственной степи, где она паразитировала на полынях морской (Artemisia maritima incana Schm.) и австрийской (A. austriaca Jacq.) [1]. В середине прошлого века O. cumana стала массовым явлением во всех районах Средней Азии, распространилась в Средиземноморье, на Балканах, в Малой Азии, Украине, на Кавказе, Поволжье, в отдельных районах Западной и Восточной Сибири [2].
Растение заразихи чрезвычайно плодовито, может образовать до 500 тысяч пылевидных семян, которые легко переносятся ветром и, находясь в почве в природных условиях, могут сохранять всхожесть до 20 лет [3].
В настоящее время в Европейских странах, возделывающих подсолнечник, O. cumana расценивается как одно из главных ограничений в производстве этой культуры. За последние пятнадцать лет усилия учёных и селекционеров представить генетические источники устойчивости к этому паразиту в гибридах подсолнечника сопровождались возникновением новых вирулентных рас, которые быстро преодолели все известные гены резистентности [4].
В Испании частый возврат подсолнечника на прежнее место ещё в 1995-1996 гг. привёл к образованию новых биотипов паразита, поразивших все устойчивые гибриды. Эти биотипы были названы расами D, E, F [5]. В настоящее время раса F распространена там повсеместно в провинциях Севилья, Кордоба, Куэнка [6]. Наиболее же засорены заразихой области Есиха и Кармона, а в Андалусии имеют место большие потери в производстве подсолнечника. Там также обнаружена и более вирулентная раса G, которая поражает гибриды, устойчивые ко всем предшествующим расам [7].
В Румынии раса F была идентифицирована в 1997 г. Новый вирулентный биотип заразихи, преодолевший устойчивость к расе F, появился там к 2005 г. (и позднее ещё один). Они были названы расами G и H [8]. Появление новых рас заразихи отмечено также в Турции [9] и Болгарии [10].
Общеизвестно, что вирулентные физиологические расы паразитов возникают в ходе сопряжённой эволюции паразита и хозяина. С “уходом” сортов и гибридов их питающих исчезают и старые расы. В ходе сортообновления появляются расы, приспосабливающиеся к новым генотипам хозяина.
На протяжении истории возделывания подсолнечника в России было три периода, когда сильное поражение посевов заразихой ставило эту культуру под угрозу исчезновения. Сопряженная эволюция паразита и хозяина приводила к появлению новых рас заразихи, способных преодолевать иммунитет устойчивых сортов и гибридов. Последняя эпифитотийная обстановка сложилась в СССР в начале 70-х годов, когда появившийся впервые в Молдавии биотип заразихи, названный молдавской расой, начал поражать прежде бывшие устойчивыми сорта и быстро распространился во всех регионах возделывания подсолнечника, особенно на Северном Кавказе. Успешная селекция новых сортов подсолнечника на устойчивость к этой расе в течение десяти последующих лет позволила решить проблему. Повсеместное возделывание этих устойчивых сортов привело к постепенному уничтожению основных запасов семян заразихи в почве.
Примерно до конца 90-х годов прошлого века особых проблем с заразихой на подсолнечнике в России не возникало. Однако в последние годы из разных мест Ростовской области, Ставропольского и Краснодарского краёв стали поступать сведения о сильной засорённости посевов подсолнечника заразихой.
Целью данного исследования было проведение сравнительного анализа вирулентности заразихи из районов Ростовской области и других регионов Российской Федерации.
Материалы и методы . В период созревания подсолнечника в 2008 г. были обследованы посевы подсолнечника в 43 районах Ростовской области на предмет их засорения заразихой (табл. 1). Обследование проводилось совместно с сотрудниками отдела семеноводства группы компаний Агроземинвест и Россельхознадзора Ростовской области. Учёт засоренности проводили следующим образом: подсчитывали число цветоносов заразихи на 50 площадках площадью 1 м2, произвольно выбранных через равные промежутки по диагонали поля. Усреднённое число цветоносов на 1 м2 использовали как показатель степени засоренности отдельных полей. Аналогичным образом вычисляли средний показатель по каждому конкретному району.
Семена заразихи были собраны в Ростовской и Волгоградской областях (2008 г.), Краснодарском (2003, 2006 гг.) и Ставропольском (2005 г.) краях. Всхожесть семян заразихи более ранних лет сбора поддерживалась хранением в морозильной камере. Использовали также семена, полученные от Y. Kaya из Турции (2008 г.), и представляющие собой смесь рас, в том числе, F, G, H. Для установления расового состава популяций паразита использовали линии дифференциаторы: 7 (восприимчив ко всем расам) и 220 устойчив к расе С, LC1002 – к расе D, LC1003 –к расе Е, LC1093 – к расе F, 16×25 – к расе G (получены из Румынии от M. Pãcureanu-Joita) и P 96 – раса F (получена из Испании от J. Melero-Vara).
Семена линий-дифференциаторов подсолнечника высевали в пластиковые цветочные ящики размером 50 x 20 x 20, заполненные почвенно-песчаной смесью (3:1), смешанной с семенами паразита. Семена каждой популяции заразихи вносили в отдельный ящик из расчёта 200 мг на 1 кг почвенной смеси. Растения подсолнечника выращивали в камере искусственного климата при 16-часовом фотопериоде и температуре 22-25 оС (рис. 1). Полив осуществляли при подсыхании верхнего слоя почвы. Через 30 дней после появления всходов растения выкапывали, отмывали корни водой и учитывали на них количество проростков клубеньков и зачатков стеблей заразихи.
Таблица 1 – Степень засорения заразихой полей подсолнечника в районах Ростовской области
2008 г.
|
№ п/п |
Район |
Посев подсолнечника, га |
Доля засоренных посевов, % |
Степень поражения заразихой*, % |
|
|
всего |
в том числе засоренных заразихой |
||||
|
1 |
Азовский |
42910 |
1042 |
2,4 |
с лабая |
|
2 |
Аксайский |
10236 |
812 |
7,9 |
Слабая |
|
3 |
Багаевский |
7813 |
4716 |
60,4 |
Слабая |
|
4 |
Белокалитвинский |
35108 |
1053 |
3,0 |
Сильная |
|
5 |
Боковский |
32476 |
998 |
3,1 |
Средняя |
|
6 |
Веселовский |
22699 |
6433 |
28,3 |
Слабая |
|
7 |
Верхнедонской |
35124 |
7326 |
20,9 |
Слабая |
|
8 |
Волгодонской |
12389 |
409 |
3,3 |
Слабая |
|
9 |
Дубовский |
12433 |
0,00 |
0,0 |
0 |
|
10 |
Егорлыкский |
52278 |
901 |
1,7 |
Слабая |
|
11 |
Заветинский |
1205 |
0,00 |
0,0 |
0 |
|
12 |
Зерноградский |
59657 |
0,00 |
0,0 |
0 |
|
13 |
Зимовниковский |
16091 |
0,00 |
0,0 |
0 |
|
14 |
Кагальницкий |
28074 |
6642 |
23,7 |
Слабая |
|
15 |
Каменский |
30285 |
685 |
2,3 |
Средняя |
|
16 |
Кашарский |
65117 |
359 |
0,6 |
Средняя |
|
17 |
Константиновский |
36759 |
3321 |
9,0 |
Сильная |
|
18 |
Красносулинский |
27554 |
289 |
1,0 |
Слабая |
|
19 |
Куйбышевский |
14567 |
748 |
5,1 |
Слабая |
|
20 |
Мартыновский |
29624 |
830 |
2,8 |
Слабая |
|
21 |
М-Курганский |
39323 |
694 |
1,8 |
Слабая |
|
22 |
Миллеровский |
56684 |
3058 |
5,4 |
Средняя |
|
23 |
Милютинский |
40244 |
366 |
0,9 |
Слабая |
|
24 |
Морозовский |
32338 |
2141 |
6,6 |
Сильная |
|
25 |
Мясниковский |
9910 |
2160 |
21,8 |
Слабая |
|
26 |
Неклиновский |
32747 |
0,00 |
0,0 |
0 |
|
27 |
Обливский |
26579 |
675 |
2,5 |
Сильная |
|
28 |
Октябрьский |
23946 |
5414 |
22,6 |
Слабая |
|
29 |
Орловский |
30230 |
4894 |
16,2 |
Слабая |
|
30 |
Песчанокопский |
47219 |
5807 |
12,3 |
Слабая |
|
31 |
Пролетарский |
24714 |
279 |
1,1 |
Слабая |
|
32 |
Ремонтненский |
2178 |
0,00 |
0,0 |
0 |
|
33 |
Р-Несветайский |
29391 |
260 |
0.9 |
Слабая |
|
34 |
Сальский |
53352 |
9082 |
17,0 |
Слабая |
|
35 |
Семикаракорский |
16006 |
7158 |
44,7 |
Средняя |
|
36 |
Советский |
18794 |
224 |
1,2 |
Средняя |
|
37 |
Тарасовский |
47076 |
2325 |
4,9 |
Средняя |
|
38 |
Тацинский |
31226 |
1987 |
6,4 |
Сильная |
|
39 |
Усть-Донецкий |
11905 |
2313 |
19,4 |
Средняя |
|
40 |
Целинский |
40162 |
5285 |
13,2 |
Слабая |
|
41 |
Цимлянский |
20737 |
519 |
2,5 |
Слабая |
|
42 |
Чертковский |
39298 |
1163 |
3,0 |
Слабая |
|
43 |
Шолоховский |
28909 |
3687 |
12,8 |
Слабая |
|
* Цветоносов заразихи на 1 м 2 при степени: слабая - < 1; средняя - 1-10; сильная - > 10 |
|||||

Рисунок 1 – Линии дифференциаторы устойчивости подсолнечника к заразихе в цветочных ящиках в камере искусственного климата, 2009 г.
При оценке вирулентност и заразихи, собранн ой на трёх полях в Морозовском районе Ростовской области, использовали самоопыленные лини и подсолнечника ВК 623 и ВК 680 селекции ВНИИМК, а также сорт ВНИИМК 8883.
Таблица 2 – Степень поражения линий подсолнечника селекции ВНИИМК заразихой (Orobanche cumana Wallr.)
из Морозовского района Ростовской области при тестировании их устойчивости в климатической камере
ВНИИМК 2008
|
Место сбора семян заразихи |
ВК 623 |
ВК 680 |
ВНИИМК 8883 |
|||
|
% поражения |
степень поражения, шт./раст. |
% поражения |
степень поражения, шт./раст. |
% поражения |
степень поражения, шт./раст. |
|
|
Краснодарский край, 2003 г. |
0 |
0 |
30 |
3 |
100 |
42 |
|
ЗАО «Вишневское» Морозовского района Ростовской области |
100 |
22 |
100 |
40 |
100 |
39 |
|
СПК «Победа» Морозовского района Ростовской области |
100 |
43 |
100 |
52 |
100 |
73 |
Результаты и обсуждение. Данные таблицы 1 показывают, что в 6 из 43 обследованных районов Ростовской области поля, занятые подсолнечником в 2008 г., были свободны от заразихи. Эти районы: Ду-бовский, Заветинский, Зерноградский, Зимовниковский, Неклиновский, Ремонтненский.
Средняя степень засорения заразихой посевов подсолнечника наблюдалась в Боковском, Каменском, Кашарском, Миллеровском, Семикаракорском, Советском, Тарасовском и Усть-Донецком районах. Здесь количество цветоносов заразихи на 1 м2 площади посева колебалось в пределах 1-10 штук. В пяти районах: Белокалитвинском, Константиновском, Морозовском, Обливском и Тацинском степень засорения посевов заразихой была сильной, свыше 10 цветоносов на 1 м2. В остальных 22 районах посевы были засорены в слабой степени (до 1 цветоноса на 1 м2), что всё-таки не позволяет считать благополучными эти районы ввиду чрезвычайной плодовитости паразита. Последующее возделывание подсолнечника на тех же полях приведёт к быстрому размножению заразихи.
Семена паразита с трёх полей из двух хозяйств Морозовского района были использованы для определения их вирулентности по отношению к инцухт-линиям селекции ВНИИМК, бывшим ранее устойчивыми к заразихе. Данные опыта, представленные в таблице 2, показывают, что к заразихе, собранной в 2003 г. в Краснодарском крае, линия ВК 623 сохраняла иммунитет. Другая линия ВК 680 поразилась ею, но c очень незначительной степенью, тогда как сорт ВНИИМК 8883 был поражен на 100 % с высокой степенью (42 клубенька на одно поражённое растение).
Другая картина наблюдалась при заражении указанных линий заразихой с полей Морозовского района. Эта популяция оказалась высоко вирулентной для обеих линий. Все растения каждой линии были поражены с высокой степенью, сопоставимой с таковой контрольного сорта ВНИИМК 8883 (см. табл. 2).
Данные таблицы 2 свидетельствуют о том, что в популяции заразихи из Морозовского района Ростовской области присутствует значительная доля биотипа высокой степени вирулентности, который преодолевает иммунитет изученных линий подсолнечника селекции ВНИИМК. Учитывая, что по данным таблицы 1 уже свыше 6 % площадей, занятых подсолнечником в Морозовском районе, засорены заразихой в сильной степени, можно прогнозировать быстрое распространение этого опасного биотипа на больших площадях при дальнейшем возделывании здесь подсолнечника.
Для идентификации рас семена заразихи были взяты с отдельных полей двенадцати районов Ростовской области (рис. 2). В наших опытах все использованные линии-дифференциаторы были поражены заразихой из Белокалитвинского, Егорлыкского, Константиновского и Тацинского районов (табл. 3). При этом линия 16 x 25, которая устойчива к расе G в Румынии, оказалась поражена с высокой степенью: 31-36 клубеньков на одно поражённое растение. Это свидетельствует, что в указанных районах имеются поля, где накоплен опасный биотип заразихи, к которому нет пока источников устойчивости. Заразиха Белокалитвинского и Тацинского районов поразила также со значительной степенью линии, устойчивые к расе F: LC 1093А (12-21 клубенек) и Р 96 (12-15 клубеньков), несущие разные гены устойчивости к ней.
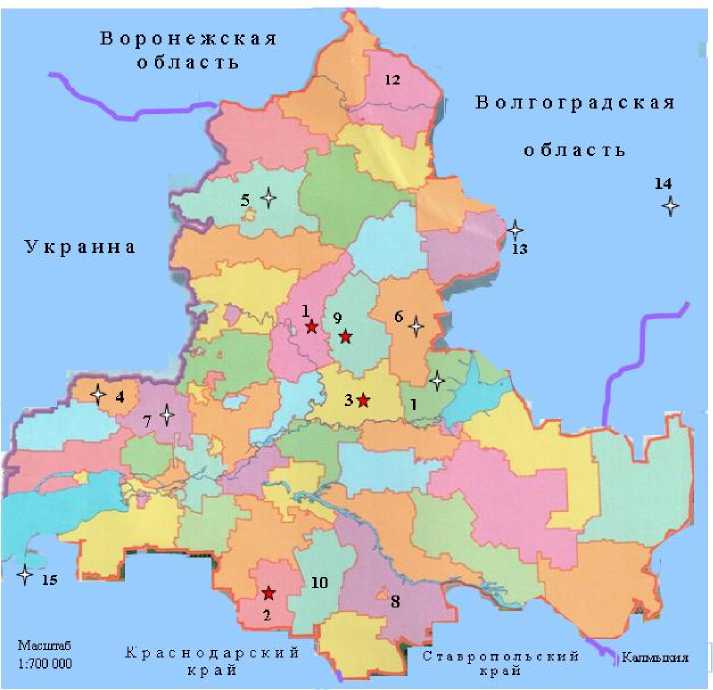
Рисунок 2 – Наличие опасного биотипа заразихи, поражающего линию подсолнечника, устойчивую к расе G , со степенью: сильной ( ), слабой ( ) в районах Ростовской области:
1 – Белокалитвинский, 2 – Егорлыкский, 3 – Константиновский, 4 – Куйбышевский,
5 – Миллеровский, 6 – Морозовский, 7 – Родионовонесветайский, 8 – Сальский,
9 – Тацинский, 10 – Целинский, 11 – Цимлянский, 12 – Шолоховский
Поражение этих и остальных линий дифференциаторов могло быть вызвано самым вирулентным биотипом, так как степень их поражения значительно меньше, чем линии 16 x 25. Поэтому дифференцировать здесь другие расы по приведенным в таблице 3 цифрам не представляется возможным. Наличие здесь в большом количестве указанного выше биотипа, преодолевающего устойчивость к расе G, превосходит значимость присутствия других, более слабых рас.
Очевидно, что в Константиновской популяции, кроме большой доли самого опасного биотипа заразихи, присутствуют расы E и F, так как имеется значительная разница в поражении линий LC1002 и LC1003 (соответственно 57 и 36 клубеньков). Поражение же обеих линий, устойчивых к расе F, может быть вызвано как наличием расы G, так и более вирулентного биотипа.
В других районах Ростовской области, биотип заразихи, поражающий линию 16 x 25, также уже имеется, хотя и в меньшем количестве (табл. 3). Популяция из Миллеровского района представлена, в основном, расой F, так как дифференциаторы LC1002 и LC1003, устойчивые, соответственно, к расам D и E, поражены, примерно, с одинаковой степенью (29 и 34 клубенька). О наличии возможной примеси расы G может свидетельствовать поражение линии LC 1093, устойчивой к расе F. Однако это поражение могло быть вызвано и примесью более вирулентного биотипа, который поразил линию 16 x 25. В популяции из Морозовского района также выделяется раса F с примесью наиболее вирулентного биотипа. В Цимлянской популяции (Мар-тинская) обнаруживаются расы E, F и упомянутый особо вирулентный биотип.
Таблица 3 – Степень поражения* линий-дифференциаторов подсолнечника заразихой из Ростовской области и других регионов
|
№ |
Район сбора заразихи в Ростовской области |
7 восприимчив |
202 (С)** Or3 |
LC1002 (D)** Or4 |
LC1003 (E)** Or5 |
ВК 623 (E)** Or5 |
LC1093 (F)** Or6 |
Р 96 (F)** or6, or7 |
16A x 25 (G)** ? |
|
1 |
Белокалитвинский |
26 |
28 |
18 |
21 |
13 |
12 |
15 |
36 |
|
2 |
Егорлыкский |
7 |
- |
18 |
13 |
4 |
3 |
1 |
34 |
|
3 |
Константиновский |
19 |
6 |
57 |
36 |
23 |
15 |
7 |
31 |
|
4 |
Куйбышевский |
- |
- |
10 |
13 |
4 |
4 |
1 |
4 |
|
5 |
Миллеровский |
- |
- |
29 |
34 |
3 |
12 |
1 |
6 |
|
6 |
Морозовский |
- |
- |
17 |
15 |
21 |
3 |
0 |
3 |
|
7 |
Родионовонесветайский |
- |
- |
6 |
23 |
8 |
1 |
2 |
6 |
|
8 |
Сальский |
- |
12 |
0 |
0 |
1 |
0 |
7 |
0 |
|
9 |
Тацинский |
11 |
- |
30 |
35 |
43 |
21 |
12 |
3 5 |
|
10 |
Целинский |
5 |
14 |
0 |
0 |
0 |
2 |
0 |
0 |
|
11 |
Цимлянский (Мартинская) |
6 |
- |
22 |
9 |
16 |
2 |
3 |
8 |
|
12 |
Шолоховский |
5 |
6 |
3 |
0 |
1 |
1 |
0 |
2 |
|
Районы сбора заразихи в других регионах |
|||||||||
|
13 |
Волгоград (окрестности) |
- |
5 |
24 |
20 |
5 |
7 |
2 |
11 |
|
14 |
Суровикино, Волгоградская область |
- |
5 |
4 |
10 |
0 |
0 |
1 |
3 |
|
15 |
Ейский, Копанская, Краснодарского края |
12 |
23 |
1 |
2 |
0 |
1 |
0 |
3 |
|
16 |
Каневской, Привольная Краснодарского края |
9 |
14 |
5 |
4 |
0 |
1 |
0 |
1 |
|
17 |
Светлоградский, Ставропольского края |
25 |
5 |
11 |
5 |
14 |
7 |
13 |
5 |
|
18 |
Европейская часть Турции (смесь F, G, H) |
- |
2 |
4 |
5 |
2 |
3 |
0 |
4 |
|
* Степень поражения – среднее число здоровых клубеньков на 1 поражённое растение через 30 дней после всходов; (выборка из пяти растений), прочерки означают, что семена линий не взошли; * *В скобках указано, к какой расе устойчива линия-дифференциатор; |
|||||||||
Более слабым по вирулентности выглядит образец заразихи из КФХ «Спартак» Сальского района. Он не поразил не только линию 16 x 25, но и большинство других дифференциаторов (см. табл. 3). Аналогично проявила себя и заразиха из Целинского района. Поражение линии 202, устойчивой к расе С, и отсутствие поражения линий LC 1002 и LC1003 свидетельствует, что эти популяции представлены преимущественно расой D с незначительной примесью двух других более вирулентных биотипов, преодолевающих: первый – ген устойчивости Or6 (в популяции Целинского района) и второй – совместное действие двух генов or6 и or7 (в популяции Сальского района). Заразиха из районов Родионовонесветайского и Шолоховского сходна с таковой из Сальского и Целинского районов.
Заразиха из популяции, собранной в окрестностях Волгограда содержит преимущественно расу F, так как поражены примерно с одинаковой степенью (24 и 20 клубеньков) линии LC1002 и LC 1003, каждая из которых устойчива, как уже упоминалось, соответственно к расам D и E. Степень же поражения других дифференциаторов не превышает таковую у линии 16 x 25 (11 клубеньков), и это не позволяет дифференцировать другие расы (см. табл. 3). Можно сказать о преимущественном содержании расы F и в другой популяции из Волгоградской области (окрестности Суровикино). Она отличается лишь меньшей долей самого вирулентного биотипа, поражающего линию 16 x 25.
Популяция заразихи из окрестностей станицы Копанской Краснодарского края содержит преимущественно расу D, так как значительную степень поражения (23 клубенька) имеет лишь линия 202, устойчивая к расе С. Поражение других дифференциаторов со степенью 1-3 не позволяет выделить в этой популяции другие расы, кроме биотипа, поражающего линию 16 x 25.
Популяция заразихи из окрестностей станицы Привольная Каневского района Краснодарского края содержит преимущественно расы D и E с незначительной примесью расы F. Сочетанием большого набора рас (C, D, E, F, G и самого вирулентного биотипа) отличается популяция заразихи из окрестностей г. Светло-града Ставропольского края.
В сравнении с большинством изученных популяций заразихи, представленных в таблице 3, образец из Турции слабее поражает использованные линии-дифференциаторы. Очевидно, что концентрация самых вирулентных биотипов заразихи в этом образце ниже. Данные таблицы 3 свидетельствуют о наличии в отечественных популяциях особо вирулентного биотипа, преодолевающего резистентность дифференциатора, который устойчив к европейской расе G заразихи. Остаётся открытым вопрос о том, аналогичен ли этот биотип расе Н, выявленной сравнительно недавно в некоторых странах: Румынии, Турции, Испании. Следует отметить, что дальнейшее возделывание подсолнечника на полях, где он выявлен, может быстро привести к опасной концентрации его семян в почве и увеличению ареала распространения.
Таким образом, из 43 обследованных районов Ростовской области лишь в 6 не обнаружена заразиха на полях подсолнечника. Остальные имели разную степень засоренности, в том числе, 14 районов – сильную и среднюю степень. Оценка вирулентности популяций заразихи из районов Ростовской области и других регионов Российской Федерации по отношению к европейским линиям дифференциаторам подсолнечника показала их расовую неоднородность. Имеются популяции, где ещё преобладает раса D (в Сальском, Целинском, Шолоховском, Родионовонесветайском районах Ростовской области и окрестностях станицы Копанской Краснодарского края). Выявлена раса Е (в Константиновском, Цимлянском районах Ростовской области и Каневском районе Краснодарского края). Широко представлена раса F (в Константиновском, Миллеровском, Морозовском и Цимлянском районах Ростовской области, а также в обеих Волгоградских популяциях и Светлоградской Ставропольского края). Особенно неблагополучны районы Ростовской области: Белокалитвинский, Егорлыкский, Константиновский, Тацинский наличием высокой доли биотипа, преодолевающего устойчивость дифференциатора к расе G, что не позволяет однозначно указать на присутствие там и других рас E, F, G. Этот биотип присутствует и в других изученных популяциях, за исключением Сальской и Целинской Ростовской области.
