Разнообразие бактерий, выделенных из района разработок месторождения калийных солей Верхнекамья

Автор: Ястребова О.В., Ананьина Л.Н., Пастухова Е.С., Плотникова Е.Г.
Журнал: Вестник Пермского университета. Серия: Биология @vestnik-psu-bio
Рубрика: Микробиология
Статья в выпуске: 10, 2009 года.
Бесплатный доступ
Из почвы, воды и галитовых отходов района солеразработок (г. Соликамск, Пермский край) методом накопительных культур выделено 12 различных по грампринадлежности штаммов бактерий. По результатам генотипирования методами REP-ПЦР и ARDRA выделенные штаммы были объединены в пять различных геномогрупп. У представителей геномогрупп определены нуклеотидные последовательности гена 16S рРНК, сравнительный анализ которых показал принадлежность штаммов к родам Bacillus, Virgibacillus, Pseudomonas, Chromohalobacter и Idiomarina. Из образца породы калийно-магниевых солей выделены два штамма, идентифицированные как представители родов Bacillus и Streptomyces. В накопительной культуре из данного образца на полноценной среде c 20% NaCl обнаружены бактерии, близкородственные бактериям родов Ralstonia и Stenotrophomonas (сходство 98%). Большинство выделенных штаммов растет при 15-20% NaCl в среде культивирования, штаммы С2 и К52 способны к росту в щелочных условиях (pH 9.0). Штамм Pseudomonas sp. К513 использует в качестве субстрата бензойную кислоту в высокой концентрации (5 г/л) при содержании в среде культивирования 3% хлорида натрия.
Короткий адрес: https://sciup.org/147204449
IDR: 147204449 | УДК: 579.26:579.222.2
The study of bacteria, isolated from the salt mining of Upper-Kama potassiummagnesium salt deposit
Using a method of enrichment, twelve strains of bacteria belonging to different taxonomic groups were isolated from soil, water and salt mine wastes from the salt mining area, of Solikmsk in the Perm region. The analysis of REP-PCR and ARDRA showed that isolated strains formed five genomic groups. Phylogenetic analysis based on 16S rDNA sequences indicated that isolates belonged to the genera Bacillus, Virgibacillus, Pseudomonas, Chromohalobacter and Idiomarina. Two strains, identified as belonging to the genera Bacillus and Streptomices, were isolated from a rock sample originating from a potassium-magnesium salt deposit. Microorganisms, closely related to the genus Ralstonia and Stenotrophomonas (98% identity) were found in the enrichment culture, obtained from the rock specimen with the use of medium supplemented with 20% NaCl. The majority of the strains grew at 15-20% NaCl in cultural medium, two strains, designated С2 and К52 grew at alkaline conditions (pH 9.0). Strain Pseudomonas sp. К513 used benzoic acid at high concentration (5 g/l) as a growth substrate at 3 % of NaCl.
Текст научной статьи Разнообразие бактерий, выделенных из района разработок месторождения калийных солей Верхнекамья
-
aИнститут экологии и генетики микроорганизмов УрО РАН, 614081, Пермь, ул. Голева, 13
-
bПермский государственный университет, 614990, Пермь, ул. Букирева, 15
Из почвы, воды и галитовых отходов района солеразработок (г. Соликамск, Пермский край) методом накопительных культур выделено 12 различных по грампринадлежности штаммов бактерий. По результатам генотипирования методами REP-ПЦР и ARDRA выделенные штаммы были объединены в пять различных геномогрупп. У представителей геномогрупп определены нуклеотидные последовательности гена 16S рРНК, сравнительный анализ которых показал принадлежность штаммов к родам Bacillus, Virgibacillus , Pseudomonas, Chromohalobacter и Idiomarina . Из образца породы калийно-магниевых солей выделены два штамма, идентифицированные как представители родов Bacillus и Streptomyces. В накопительной культуре из данного образца на полноценной среде c 20% NaCl обнаружены бактерии, близкородственные бактериям родов Ralstonia и Stenotrophomonas (сходство 98%) . Большинство выделенных штаммов растет при 15-20% NaCl в среде культивирования, штаммы С2 и К52 способны к росту в щелочных условиях (pH 9.0). Штамм Pseudomonas sp. К513 использует в качестве субстрата бензойную кислоту в высокой концентрации (5 г/л) при содержании в среде культивирования 3% хлорида натрия.
В последние десятилетия интенсивное исследование микрофлоры экосистем с высоким засолением среды позволило выделить и охарактеризовать умеренно и экстремально галофильные бактерии, различные по систематическому положению и отличающиеся разнообразием ферментных систем (Ventosa et al., 1998). Данные бактерии представляют интерес в связи с высоким биотехнологическим потенциалом, в частности они могут быть использованы для получения осмопротектерных соединений, ферментов, активных при высоком осмотическом давлении среды, а также для создания биопрепаратов очистки почв и стоков со сложным, полихимическим характером загрязнения (Margesin, Schinner, 2001).
Цель работы – исследование разнообразия микроорганизмов почвы, воды, образца породы и галитовых отходов района солеразработок.
Методы исследования
Образцы почвы, воды, галитовых отходов, а также образец породы для постановки накопительных культур были отобраны на территории Верхнекамского месторождения калийно-магниевых солей (ОАО ”Сильвинит” г. Соликамск). Отбор образца почвы производился на глубине 5 см на расстоянии 2 м от солеотвала в осеннее время года. Проба воды была взята из прилегающего к солеотвалу водоема с глубины 5 см от поверхности и на расстоянии 70 см от береговой линии. Образец галитовых отходов отобран с глубины 5 см непосредственно с солеотвала.
Метод накопительного культивирования
Для выделения штаммов микроорганизмов использовали метод накопительного культивирования. Для постановки накопительных культур (НК) использовалась жидкая минеральная среда Раймонда (Розанова, Назина, 1982) с разными концентрациями NaCl (5, 10, 15, 20, 25%). В качестве субстратов использовались: бензоат (1 г/л) или триптон (0.5 г/л) и дрожжевой экстракт (0.25 г). Образцы почвы, воды или галитовых отходов (1 г) были помещены в 250 мл колбы с 100 мл ростовой среды. Инкубация проводилась в течение 2 недель на термокачалке (100 об/мин.) при температуре 28оС. Из накопительных культур путем высева на селективные агаризованные среды выделяли чистые культуры микроорганизмов. Чистота культур контролировалась путем высева на агаризованную полноценную среду Раймонда, содержащую 3% хлорида натрия.
Изучение физиологических свойств
Устойчивость бактерий к NaCl (концентрация соли от 3 до 25%) определяли на полноценной агаризованной среде Раймонда. Рост бактерий учитывали на седьмой день.
Рост бактерий при разных значениях рН определяли при концентрации Na+ 0.8-0.85 М в буферных системах (для рН 5 - ацетатный буфер, для рН 6-8 - фосфатный буфер, для рН 810 - трис-HCl буфер (“Методы общей бактериологии”, 1983), приготовленных на основе минеральной среды Раймонда, содержащей глюкозу в качестве источника углерода и энергии. Штаммы культивировались на агаризованной среде при рН 5, 7, 8, 9, 10. Рост учитывали на седьмой день.
Идентификация бактерий
Идентификацию бактерий проводили на основе изучения их культуральных, морфологических и биохимических свойств (Определитель бактерий Берджи, 1997; Методы общей бактериологии, 1983). Морфологию и жизненный цикл бактерий изучали у выращенных на агаризованной среде 12-72часовых культур с использованием световой микроскопии (фазовый контраст).
Амплификация гена 16S рРНК. Для амплификации использовали бактериальные праймеры 27F и 1492R. Амплификацию проводили так, как описано у Гавриш и др. (2004).
REP-ПЦР (полимеразная цепная реакция повторяющихся экстрагенетических палиндром-ных последовательностей ДНК) проводили по методу Versalovic (1994). Продукты реакции разделяли электрофорезом в 1.5% агарозном геле, приготовленном на 1 х ТВЕ буфере (Маниатис и др., 1984).
Расщепление ДНК эндонуклеазами рестрикции. 5 мкл ПЦР продукта были обработаны 1 ед. рестрикционных ферментов Hha I и Mse I (Fermentas, Литва), используя для каждой эндонуклеазы рекомендованный буфер и соответствующий температурный режим. Фрагменты рестрикции разделяли электрофорезом в 1.2% агарозном геле (Scortichini et al., 2002).
Денатурирующий градиентный гель-электрофорез ампликонов 16S рРНК был выполнен в 6% (в/об.) полиакриламидном геле, содержащем линейный денатурирующий химический градиент от 35 до 50%, где 100% составляет 7М мочевина и 40% - формамид, согласно протоколу Muyzer et al. (1993).
Определение нуклеотидной последовательности гена 16S рРНК. Секвенирование гена 16S рРНК, амплифицированного с использованием универсальных бактериальных праймеров (Гавриш и др., 2004), проводили с применением набора реактивов CEQ Dye Terminator Cycle Sequencing Kit на автоматическом секвенаторе MegaBASE 1000 (JSC GE “Healthcare”, США).
Филогенетический анализ .
Нуклеотидную последовательность 16S рДНК изучаемого изолята сравнивали с нуклеотидными последовательностями типовых штаммов близкородственных видов из базы данных GenBank с помощью программы CLUSTAL W и корректировали вручную (Thompson et al., 1994). Эволюционное расстояние, выраженное как число замен на 100 нуклеотидов, рассчитывали согласно методу Jukes, Cantor (1969). Построение филогенетического древа производили с помощью пакета программ TREECON c использованием метода “neighbor-joining” (NEIGHBOR) (Van de Peer, DeWachter, 1994). Оценку статистической достоверности ветвления (“bootstrap-анализ”) проводили с использованием соответствующей функции программы TREECON на основе 1000 альтернативных деревьев.
Результаты и обсуждение
Из образцов почвы, воды и галитовых отходов района солеразработок ОАО “Сильвинит” (г. Соликамск, Пермский край) методом накопительного культивирования (НК) на полноценной и минеральной среде Раймонда с разным содержанием NaCl были выделены 14 штаммов бактерий, различных по грампринадлежности, морфологии клеток и колоний.
Из НК на полноценной среде с содержанием NaCl 5 %, 10 и 15% выделено два грамположительных (С32, С41) и шесть грамотрицательных (В151, В152, В153, В154, В201, В202) штаммов бактерий. Выделенные штаммы обладают каталазной активностью, оксидазоотрицательны. Из НК на минеральной среде с бензоатом в качестве субстрата и содержанием NaCl 5% выделено четыре бактериальных штамма (К51, К52, К511 и К513), различных по грампринадлежности, каталазной и оксидазной активностям.
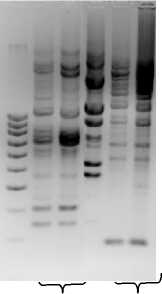
Проведено сравнение выделенных штаммов с использованием методов REP-ПЦР и ARDRA. Анализ полученных REP-ПЦР профилей фрагментов геномной ДНК исследуемых штаммов показал, что два штамма - К511 и К513 входят в одну геномогруппу (I тип) (рис. 1). Штаммы В201 и В202 также составляют одну геномогруппу (II тип). Штамм В153 отличается от представителей данных групп и выделен в отдельную геномогруппу (III тип).
м 1 2 3 4 5
м 1 2 3 4 5 6 7 8 1 2 3 4 5 6 7 8
IV V I II III IV V I II III
I III II
А
Mbo I Hha I
B
Рис. 1. Электрофореграмма продуктов амплификации REP-ПЦР штаммов-деструкторов (A) и 16S рДНК, обработанных рестриктазами Mbo I и Hha I (B). На электрофореграмма А: м - маркер 1 kb (“Силекс”, Россия), 1 - К511, 2 - К513, 3 - В153, 4 - В201, 5 - В202. На электрофореграмма В: м - маркер 1 kb (“Силекс”, Россия), 1 - В151, 2 - В152, 3 - К513, 4 - К511, 5 - В201, 6 - Paenibacillus sp. SN501, 7 - Bacillus sp. 2508, 8 - В153.
0.02
I----------------------------------1
I B. sabtilis subsp. suhriiis DSM101 (AJ2 76351)
B. mojavensis IFO157181 (AB021191)
B. subtilis subsp. spiziaeui NRRLB-23049T(AFD749 7D)i
50 I B. vaiiismortis DSM110311
B. omrZatiy!Me/aciezisATCC23350T(X60605)
B. atrophaeus JCM90701 (ABO21181)
B. soHorensis NRRL B-231541 (EU138473)
В. lichMformis DSM131 (X68416)
63 -C2
B. as^us 24 KT fAJ831843^
B. tequileKsis 10b1 (AY197613)
98J B. pumiius DSMZ271 (AY456263)
1 B. safensis FO-OMb1 (AF234854)
B. uhtotdims 41KF2bT (AJS31842)
B. aerophihisl'SK1 (AJ831844)
B. stratosphericiis 41KF2aT (AJ831841)
B. idrienss S MC 4352-2T (AY904033)
100 I B. in&cus Sdtt1 (AJ583158)
B. cibi JG-3O1 (AY55O276)
----Б. taeanensis BH0300171 (AY6039^)
В. «igicola KMM3 7371 (AY22S462)
— В. hwajinpoensis SW-721 (AF541966) B. decoleradouis LMG 19507T(AJ315075)
AlicydobacUlus addocaMarius NBRC 15б52т (AB271754)
Рис. 2. Филогенетическое древо, построенное с использованием метода “neighbor-joining”, отображающее положение штамма С2 в системе рода Bacillus . Масштаб соответствует 2 нуклеотидным заменам на каждые 100 нуклеотидов. Цифрами показана статистическая достоверность порядка ветвления, определенная с помощью “bootstrap”-анализа 1000 альтернативных деревьев
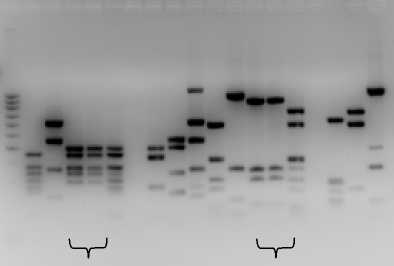
Результаты рестрикционного анализа амплифицированных 16S рДНК (ARDRA) с использованием рестриктаз Mbo I и Hha I подтвердили принадлежность штаммов К511 и К513 к одной геномогруппе (I тип). Штаммы В201, В153, В151 и В152 отличаются от представителей данной группы и различаются между собой. Они выделены в отдельные геномогруппы II, III, IV и V типа, соответственно (рис. 2).
У представителей геномогрупп были определены нуклеотидные последовательности гена 16S рРНК размером около 1400 п.н.
Сравнительный анализ нуклеотидной последовательности гена 16S рРНК показал 99.5% сходство штамма К51 с типовым штаммом вида Bacillus alсalophilus . Штамм К513 имеет наибольший уровень гомологии с типовым штаммом вида Pseudomonas putida (99%), штамм К52 - с типовым штаммом вида Virgibacillus picturae (99,5%), штаммы В201 и В151 – c Chromohalobacter canadensis (99.4%), штаммы B152 и В153 – с типовым штаммом вида Idiomarina loihensis (99.5%).
Из образца породы калийно-магниевых солей методом НК на полноценной среде Раймонда, содержащей 5% NaCl, выделено два штамма: С31 и С2. Филогенетический анализ нуклеотидной последовательности гена 16S рРНК штамма С2 выявил его принадлежность к роду Bacillus . Наибольшее сходство (100%) изолят С2 имеет с B. licheniformis DSM13T
(X68416) (рис. 3). Штамм С31 имеет наибольший уровень гомологии с типовым штаммом вида Streptomyces ambofaciens (99.4%).
Грамположительные штаммы способны к росту как в отсутствие соли, так и при концентрации NaCl до 3%. Они являются галотолерантными микроорганизмами по классификации Кашнера (Кашнер, 1981). Грамотрицательные штаммы не растут в отсутствие соли в среде, способны расти при концентрации NaCl 10%–15%, а штамм В201 – до 20%. Данные штаммы относятся к галофильным микроорганизмам (таблица 1).
Все выделенные бактерии растут при значении рН 7.0, штаммы Bacillus sp. C2 и Virgibacillus sp. К52 являются алкалофильными и способны к росту в щелочных условиях среды - рН 9 (табл. 1). Способность спорообразующих бактерий к росту в условиях повышенной солености и высоких значений pH среды описана для ряда галоалкалофильнх штаммов рода Bacillus, выделенных из содовых озер и солончаков (Vargas et al., 2005; Sorokin et al., 2008).
Два штамма - Bacillus sp. К51 и Virgibacillus sp. К52 - способны к слабому росту на бензойной кислоте в среде, содержащей NaCl в концентрации до 4 %.
Грамотрицательный штамм Pseudomonas sp. К513 эффективно растет на бензоате (1 г/л) в среде без добавления соли и в среде, содержащей до 6% хлорида натрия.
Таблица 1
Рост штаммов бактерий в полноценной среде Раймонда при различных значениях рН среды и в присутствии различных концентраций хлорида натрия
|
Штамм |
Концентрация NaCl (%) |
Значение pH |
||||||
|
Без NaCl |
3 |
15 |
20 |
6.0 |
7.0 |
8.0 |
9.0 |
|
|
Bacillus sp. C2 |
+ |
+ |
+ |
+ |
+ |
+ |
± |
|
|
Streptomyces sp. C31 |
+ |
+ |
+ |
+ |
+ |
|||
|
Chromohalobacter sp. B151 |
+ |
+ |
||||||
|
Idiomarina sp. B152 |
+ |
+ |
||||||
|
Idiomarina sp. B153 |
+ |
+ |
+ |
|||||
|
Chromohalobacter sp. B201 |
+ |
+ |
+ |
+ |
||||
|
Chromohalobacter sp. B202 |
+ |
+ |
+ |
|||||
|
Pseudomonas sp. К511 |
+ |
+ |
+ |
|||||
|
Pseudomonas sp. К513 |
+ |
+ |
+ |
|||||
|
Bacillus sp. К51 |
+ |
+ |
+ |
|||||
|
Virgibacillus sp. К52 |
+ |
+ |
± |
+ |
+ |
+ |
||
Примечание: “+” – хороший рост (колонии размером более 3 мм); “±” - средний рост (колонии размером 1-2 мм); “–“ - отсутствие роста бактерий.
Известна способность ряда грамотрицательных галофильных штаммов, в частности Halomonas campisalis и Chromohalobacter sp. HS-2, к утилизации бензоата в концентрации 1.6 г/л и 0.6 г/л соответственно (Kim et al., 2008; Oie et al., 2007). Выделенный нами штамм К513 растет на данном субстрате в высокой концентрации - до 5 г/л (рис. 3).
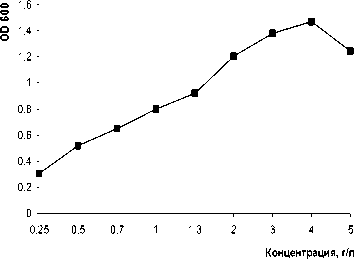
Рис. 3. Максимальное значение оптической плотности штамма К513 в минеральной среде Раймонда (3% NaCl) при разных концентрациях бензойной кислоты
I Uncultured bacterium
II « Ralstonia pickettii »
III « Stenotrophomonas maltophilia »
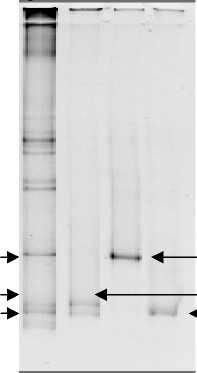
Рис. 4. ДГГЭ продуктов амплификации фрагмента гена 16S рРНК, полученных с матрицы суммарной бактериальной ДНК накопительной культуры, культивируемой на 20% NaCl ( 1 ); 2, 3, 4 – реамплифицированные элюаты
1 2 3 4
Из образца породы калийно-магниевых солей получена накопительная культура на полноценной среде Раймонда, содержащей 20% NaCl. В НК преобладали палочковидные прямые и искривленные клетки, одиночные, в парах и собранные в цепочки. Размер клеток варьировал от 0.4 х 0.9 мкм до 0.5 х 2.8 мкм. Однако при высеве на агаризованные среды с разным содержанием хлорида натрия (от 3% до 20%) бактериального роста не наблюдалось. Для дальнейшего исследования НК был использован метод денатурирующего градиентного гель-электрофореза (рис. 4).
У фрагментов 16S рДНК, отличающихся электрофоретической подвижностью и элюированных из агарозного геля, определены нуклеотидные последовательности. Показано, что в составе НК присутствуют бактерии разных классов, близкородственные бактериям рода Ralstonia (сходство с типовым штаммом Ralstonia pickettii HPC578T 95%) и рода Stenotrophomonas (сходство с типовым штаммом Stenotrophomonas maltophilia PSM-1T 99%).
Заключение
В исследуемых образцах почвы, воды и галитовых отходов района солеразработок обнаружены бактерии, отличающиеся по физиологическим характеристикам и таксономическому положению. Выделены бактерии класса Gammaproteobacteria (родов Pseudomonas, Chromohalobacter, Idiomarina, Stenotrophomonas), класса Betaproteobacteria (рода Ralstonia), класса Bacilli (родов Bacillus, Virgibacillus) и класса Actinobacteria (рода Streptomyces). Исследуемые штаммы представлены галофильными, галотолерантными и галоалкалофильными бактериями, ряд штаммов способен к деструкции бензоата при высокой солености среды. Изолированные штаммы бактерий являются перспективными для использования при биоремедиации почв и стоков с повышенным содержанием солей и решения ряда других биотехнологических задач.
Работа поддержана грантом РФФИ-Урал №07-04-96078_а., ФЦП “Научные и научнопедагогические кадры инновационной России”, тема ”Разработка биокаталитических технологий синтеза органических кислот и энантиомерно-чистых соединений для полимерной химии, медицины и экологической биотехнологии на основе микроорганизмов-продуцентов, селекционированных из природных и антропогенно-измененных почв” ГК№02.740.11.0078.
Список литературы Разнообразие бактерий, выделенных из района разработок месторождения калийных солей Верхнекамья
- Гавриш, Е.Ю. Три новых вида бревибактерий -Brevibacterium antiguum sp. nov., Brevibacterium aurantiacum sp. nov. и Brevibacterium permense sp. nov./Е.Ю. Гавриш, В.И. Краузова, Н.В. Потехина и др.//Микробиология. 2004. Т. 73., №2. С. 218-225.
- Кашнер Д. Жизнь микробов в экстремальных условиях. М.: Мир, 1981. 365 с.
- Маниатис Т., Фрич Э., Сэмбрук Дж. Методы генетической инженерии//Молекулярное клонирование. М.: Мир, 1984. 390 с.
- Методы общей бактериологии//пер. с англ.; под ред. Ф. Герхардт и др. М.: Мир, 1983. Т. 1, 2, 3.
- Определитель бактерий Берджи/пер. с англ.; под ред. Дж. Хоулта и др. М.: Мир, 1997. Т. 1, 2.
- Розанова Е.П. Углеводородокисляющие бактерии и их активность в нефтяных пластах/Е.П. Розанова, Т.Н. Назина//Микробиология. 1982. Т. 51. С. 324-348.
- Jukes T.H.//Mamallian protein metabolism/T.H. Jukes, C.R. Cantor. New York: Acad. Press. 1969. P. 21-132.
- Kim D. Molecular cloning and functional characterization of the genes encoding benzoate andp -hydroxybenzoate degradation by the halophilic Chromohalobacter sp. strain HS-2/D. Kim, S.W. Kim, K.Y. Choi//FEMS Microbiol. Lett. 2008. Vol. 280. P. 235-241.
- Margesin R. Potential of halotolerant and halophilic microorganisms for biotecnilogy/R. Margesin, Schinner F.//Extremophiles. 2001. Vol. 5. P. 73-83.
- Muyzer G. Profiling of complex microbial populations by denaturing gradient gel electrophoresis analysis of polymerase chain reaction-amplified genes coding for 16S rRNA/G. Muyzer, E.C. de Waal, A.G. Uitterlinden//Appl. Environ. Microbiol. 1993. Vol. 59. P. 695-700.
- Oie C.S.I. Benzoate and salicylate degradation by Halomonas campisalis, an alkaliphilic and moderately halophilic microorganism/C.S.I. Oie, C.E. Albaugh, B.M. Peyton//Water research. 2007. Vol. 41. № 6. P. 1235-1242.
- Scortichini М. Bacteria associated with hazelnut (Corylus avellana L.) decline are of two groups: Pseudomonas avellanae and strains resembling P. syringae pv. syringae/М. Scortichini, U. Marchesi, M.-P. Rossi//Appl. Environ. Microbiol. 2002. Vol. 68. P. 476-484.
- Sorokin I.D., Haloalkaliphilic diazotrophs in soda solonchak soils/I.D. Sorokin, I.K. Kravchenko, E.V. Doroshenko//FEMS Microbiol Ecol. 2008. V. 65. P. 425-433.
- Thompson J.D. CLUSTAL W: improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position specific gap penalties and weight matrix choice/J.D. Thompson, D.G. Higgins, T.J. Gibson//Nucleic. Acids. Res. 1994. Vol. 22. P. 4673-4680.
- Van de Peer Y. TREECON for Windows a software package for the construction and drawing of evolutionary trees for the Microsoft Windows environment/Y. Van de Peer, R. DeWachter//Comput. Appl. Biosci. 1994. Vol. 10. P. 569-570.
- Vargas A.V. Bacillus bogoriensis sp. nov., a novel alkalophilic, halotolerant bacterium isolated from Kenyan soda lake/A.V. Vargas, O.D. Delgado, R. Hatti-Kaul, B. Mattiasson//International Journal of Systematic and Evolutionar Microbiology. 2005. Vol. 55. P. 899-902.
- Ventosa A. Biology of halophilic aerobic bacteria/A. Ventosa, J.N. Joaquhn, A. Oren//Microbiol. and Molec.Biol. Rev. 1998. Vol.2. P. 504-544.
- Versalovic J. Genomic fingerprinting of bacteria using repetitive sequence-based polymerase chain reaction/J. Versalovic, M. Schneider, F.J. de Bruijn//Meth. Cell. Mol. Biol. 1994. Vol. 5. P. 25-40.
