Разработка способа индикации и идентификации Vibrio parahaemolyticus с помощью real-time ПЦР

Автор: Полеева Марина Владимировна, Чемисова Ольга Сергеевна, Трухачев Алексей Леонидович
Журнал: Вестник Пермского университета. Серия: Биология @vestnik-psu-bio
Рубрика: Микробиология
Статья в выпуске: 2, 2019 года.
Бесплатный доступ
Бактерии Vibrio parahaemolyticus могут быть причиной возникновения острого кишечного заболевания у людей, протекающего по типу пищевой токсикоинфекции. Успех лечебно-профилактических и санитарно-противоэпидемических мероприятий, направленных на предотвращение распространения «галофилезов», во многом зависит от своевременной точной идентификации штаммов вибрионов, выделяемых от больных, из морепродуктов и при мониторинге объектов окружающей среды. Представлена разработка способа идентификации штаммов вида V. parahaemolyticus методом ПЦР в режиме «реального времени» (Real-Time PCR), который включает выделение хромосомной ДНК из исследуемого материала, постановку ПЦР со сконструированными нами праймерами, специфичными к участку гена коллагеназы vppC, и зондом, позволяющим детектировать амплифицированный фрагмент и проводить учет полученных результатов в режиме реального времени. Показана возможность индикации и идентификации микроорганизмов вида V. parahaemolyticus при исследовании проб биологического материала (рыба, морепродукты) на основе метода Real-Time PCR.
Праймеры, металлопротеаза (коллагеназа)
Короткий адрес: https://sciup.org/147227079
IDR: 147227079 | УДК: 579.843:57.083.18:576.3:616-078 | DOI: 10.17072/1994-9952-2019-2-175-181
The development of real-time PCR method for indication and identification of Vibrio parahaemolyticus
Bacteria V. parahaemolyticus - gram-negative halophilic bacteria that can be the cause of acute intestinal disease in humans, occurring as food toxicoinfection. The success of the treatment-and-prophylactic and sanitary-anti-epidemic measures aimed at preventing of "halophiles" spreading largely depends on timely and accurate identification of Vibrio cholerae strains isolated from patients, seafood and under monitoring of environmental objects. This article presents the development of a method for identifying V. parahaemolyticus strains by Real-Time PCR, which includes the isolation of chromosomal DNA from the test material, setting up the PCR method with constructed primers specific to the site of the collagenase gene vppC and probe, allowing to detect the amplified fragment and to record the results in real time. The possibility of indication and identification of microorganisms of V. parahaemolyticus species under the study of samples of biological material (fish, seafood) based on the Real-Time PCR method is shown.
Текст научной статьи Разработка способа индикации и идентификации Vibrio parahaemolyticus с помощью real-time ПЦР
Vibrio parahaemolyticus – грамотрицательная галофильная бактерия, встречающаяся в природе в морских и эстуарных водах и часто изолируемая из различных морепродуктов – устриц, креветок, крабов, лобстеров, омаров, раков, рыбы (треска, сардина, скумбрия, камбала), осьминогов, гребешков и замороженных морепродуктов во многих странах мира [Spanggaard et al., 2000; Hara-Kudo et al., 2001; Deepanjali et al., 2005]. Этот возбудитель вызывает заболевание по типу острого гастроэнтерита, характеризующегося тошнотой, рвотой, спаз- мами в животе, диареей [Chakraborty, Nair, Shinoda, 1997; Daniels et al., 2000]. Выявлена взаимосвязь обсемененности вибриофлорой воды и рыбы с заболеваемостью людей. Заражение людей происходит при употреблении в пищу некачественно приготовленных продуктов моря. Так, установлено, что последние годы в Астраханской обл. острые кишечные инфекции, обусловленные вибрионами, составляют около 14% всех диарей. В районах промысла дельты р. Волги различные вибрионы были выделены из судака, осетра, белуги, стерляди. Заболевания, вызываемые парагемоли-тическими вибрионами и связанные с употребле-
нием в пищу морепродуктов, регистрируются на побережье Черного и Азовского морей [Домниц-кий, 2016]. В 2012 г. была зарегистрирована вспышка пищевой токсикоинфекции, вызванной V. parahaemolyticus в Хасанском р-не Приморского края.
Успех лечебно-профилактических и санитарноэпидемиологических мероприятий, направленных на предотвращение распространения «галофиле-зов», во многом зависит от своевременной точной идентификации штаммов вибрионов, выделяемых от больных, из морепродуктов, а также при мониторинге объектов окружающей среды.
В настоящее время существует большое количество микробиологических методов детекции, основанных на культивировании, выделении чистых культур и биохимической идентификации этих вибрионов [Лабораторная …, 2004]. Сложность идентификации и дифференциации парагемолити-ческих от других близкородственных видов вибрионов традиционными методами обусловлена их большим фенотипическим сходством, вариабельностью признаков и, как следствие, небольшой относительной диагностической ценностью отдельных таксономических тестов.
Молекулярно-генетические методы находят широкое применение, как для идентификации культивируемых микроорганизмов, так и для детекции специфических групп бактерий. Для быстрой идентификации V. parahaemolyticus успешно применяется метод MALDI-ToF масс-спектрометрии; однако, не все лаборатории оснащены необходимым оборудованием [Чемисова и др., 2013; Полеева, Чемисова, 2018].
Методы диагностики бактериальных инфекций на основе видоспецифичной полимеразной цепной реакции (ПЦР-диагностика) показали свое преимущество перед классическими микробиологическими методами [Белькова, Дзюба, Суханова, 2009] и могут стать основой молекулярногенетического мониторинга ассоциированной микрофлоры водных организмов. Они требуют меньшего времени и эффективны в детекции конкретных патогенов на всех стадиях заболевания. ПЦР-диагностика позволяет корректно идентифицировать микроорганизмы, культивирование и определение которых по классическим биохимическим тестам затруднительно. Эти методы активно используются в странах с развитой аквакультурой, постоянно совершенствуются и развиваются [Buller, 2004; Белькова и др., 2010].
В последние годы широкое распространение получил метод идентификации микроорганизмов с помощью полимеразной цепной реакции с детекцией продуктов амплификации в режиме реального времени (Real-Time PCR), преимуществами которого являются объединение этапов амплифика- ции и детекции, что упрощает процедуру анализа, снижает риск контаминации, позволяет проводить количественный учет результатов, обеспечивает высокую специфичность и чувствительность анализа [Ребриков и др., 2009].
Ранее зарубежными авторами, были выявлены видоспецифичные для V. parahaemolyticus участки ДНК и разработаны методы идентификации с помощью ПЦР с электрофоретическим способом учета результатов [Di Pinto, Ciccarese, Tantillo, 2005; Luan et al., 2007; Reham, Amani, 2012; Li et al., 2016]. Однако учет результатов реакции с помощью электрофореза требует дополнительного времени, организации отдельной рабочей зоны в связи с повышенным риском контаминации. Эти обстоятельства усложняют использование метода ПЦР с учетом результатов с помощью электрофореза в геле. Метод Real-Time ПЦР лишен этих недостатков. На сегодняшний день в Российской Федерации отсутствуют зарегистрированные олигонукле-отидные праймеры и зонды для видовой идентификации вида V. parahaemolyticus , а существующие зарубежные тест-системы являются дорогостоящими для широкого применения в лабораторной практике.
Цель данной работы – разработка способа индикации и идентификации микроорганизмов вида V. parahaemolyticus при исследовании проб биологического материала (рыба, морепродукты) на основе метода полимеразной цепной реакции с детекцией продуктов амплификации в режиме реального времени (Real-Time PCR).
Материалы и методы исследования
В работе были использованы штаммы V. parahaemolyticus (100 штаммов) из коллекции Музея живых культур ФКУЗ Ростовского-на-Дону противочумного института Роспотребнадзора, а также коллекционные штаммы других видов рода Vibrio – V. alginolyticus (25 штаммов), V. fluvialis (10 штаммов), V. hollisae (5 штаммов), V. vulnificus (10 штаммов), V. harvey (5 штаммов), V. mimicus (5 штаммов), V. furnisii (5 штаммов) и V. fortis (2 штамма).
Для анализа нуклеотидной последовательности ДНК V. parahaemolyticus была использована база данных GeneBank. Для поиска уникальных последовательностей специфического гена были использованы ресурсы GeneBank-on-line Blast. Для конструирования праймеров и зонда было применено программное обеспечение PrimerM (ФКУЗ Ростов-ский-на-Дону противочумный институт Роспотребнадзора) и BLAST NCBI.
Синтез сконструированных нами праймеров и зонда был осуществлен компанией «Синтол» (г. Москва). Для проведения амплификации использована коммерческая стандартизированная реак- ционная смесь для проведения ПЦР в режиме «реального времени» компании «Синтол» (г. Москва). Амплификацию и флуоресцентную детекцию проводили на автоматическом детектирующем термоциклере «ДТ-Lite» (ДНК-технология, Россия), в режиме «реального времени».
Подготовка материала для исследования
-
1. Штаммы микроорганизмов. Суточные агаровые культуры суспендировали в дистиллированной воде до 1×109 микробных клеток в 1 мл (м. кл./мл) и обеззараживали прогреванием при 100ºС в течение 30 мин., согласно МУ 1.3.2569-09 [МУ 1.3.2569-09, 2010]. Затем дебрис осаждали центрифугированием при 10 тыс. об/мин в течение 5 мин. и использовали супернатанты в качестве ДНК-матриц при ПЦР.
-
2. Образцы тканей рыбы (судака), искусственно контаминированной бактериями V. parahaemoly-ticus . В исследование брали кусочки кожи с чешуей и мышцами. Навеску массой 1 г измельчали и тщательно растирали в стерильной ступке с 2–3 г стерильного кварцевого песка. Добавляли 10 мл забуференного физиологического раствора, тщательно перемешивали. Затем к гомогенату добавляли 0.1мл микробной взвеси V. parahaemolyticus , содержащей 109–103 м. кл./мл согласно стандарту мутности ГИСК им. Тарасевича. Тщательно перемешивали и давали отстояться приготовленной смеси 20 мин. при комнатной температуре. Затем через стерильный тампон отбирали пробу (надосадочную жидкость) объемом 0.5 мл и использовали в качестве ДНК-матриц при ПЦР.
-
3. При использовании бактериальных взвесей с концентрацией 104–101 м. кл./мл проводили дополнительное обогащение в 1%-ной пептонной воде с 2%-ным NaCl в течение 8 ч. Контроль количества клеток проводили бактериологическим высевом на плотную питательную среду с последующим подсчетом колоний. ПЦР ставили через 2, 4,
-
6, 8 ч. выращивания в среде обогащения.
Выделение ДНК проводили с помощью комплекта реагентов для экстракции ДНК из клинического материала «АмплиПрайм ДНК-сорб-В», согласно прилагаемой инструкции. В качестве контроля реакции использовали бактериальные взвеси в концентрации 109–103 м. кл./мл согласно стандарту мутности ГИСК им. Тарасевича.
Приготовление реакционной смеси для ПЦР . Реакцию проводили в 25 мкл смеси, содержащей: буферный раствор для ПЦР в реальном времени; 0.25 мМ каждого из дезоксинуклеозидтрифосфатов; 1 ед. Taq-полимеразы с функцией «горячего старта»; 2.5 мМ хлористого магния; 50 пкМ каждого праймера; 50 пкМ зонда и 20 нг хромосомной ДНК одного из исследуемых образцов.
Амплификация . Постановку реакции амплификации в автоматическом детектирующем ам-плификаторе ДТ-Lite (ДНК-технология, Россия), в котором были заданы следующие условия: первичной денатурации и активация Taq-полимеразы (94°С, 15 мин.), 35 циклов, включающих этапы 94°С, 30 с.; 55°С, 30 с. (детекция) и 72°С, 30 с.
Детекция флуоресценции производится автоматически в ходе проведения ПЦР с помощью детектирующего амплификатора. При наличии в исследуемой пробе ДНК гена металлопротеазы (коллагеназы) вида V. parahaemolyticus с помощью используемой пары специфических праймеров гибридизируется зонд, так как является комплементарным ему (фрагменту), а последующее разрушение зонда Taq-полимеразой приводит к началу флуоресцентного свечения флуорофора ROX с длиной волны 605 нм, которое регистрируется прибором. Учет детекции флуоресцентного свечения по соответствующей длине волны в амплифи-каторе отражается на мониторе компьютера, связанного с прибором, в виде графиков, на которых представлены кривые и численные значения, отражающие уровни флуоресцентного свечения определенной длинны волны, соответствующие каждой пробе, взятой для исследования.
Результаты и их обсуждение
С помощью программного обеспечения PrimerM (ФКУЗ Ростовский-на-Дону противочумный институт Роспотребнадзора) и BLAST NCBI был проанализирован участок гена металлопротеа-зы (коллагеназы), в результате чего определен специфический фрагмент гена, который был использован в качестве мишени для конструирования специфического зонда, комплементарного соответствующей последовательности ДНК. Зонд имел следующую структуру: vppCProbaA (ROX-CG-TTC-ACA-ACC-ACC-AAC-AGC-AAC-GAC-TTG-BHQ2). Зонд комплементарен специфическим продуктам ПЦР, гибридизуясь с ними и впоследствии разрушаясь под воздействием ДНК-полимеразы во время ферментативной реакации, что приводит к началу флуоресцентного свечения флуорофора, связанного с зондом. Данный специфический зонд vppCProbaA содержит в своем составе флуоресцентную метку ROX, гаситель флуоресценции BHQ2.
Для амплификации в ПЦР фрагмента ДНК, содержащего мишень для зонда и в соответствии с нуклеотидной последовательностью зонда, были сконструированы и синтезированы специфичные для вида V. parahaemolyticus праймеры:
– праймер vppCFor (CGG-CAA-GCG-TGG-TTT-GTG-AC);
– праймер vppCRev (CGT-TGA-TGC-AAC-TTG-CAC-CTT-G).
Был получен патент на изобретение «Способ идентификации штаммов вида Vibrio parahaemolyticus методом ПЦР в режиме реального времени № 2644232 от 8 февр.2018 г.
При апробации предложенного способа на коллекционных штаммах V. parahaemolyticus с точно установленной видовой принадлежностью и штаммах видов V. alginolyticus , V. fluvialis , V. hollisae , V. vulnificus , V. harvey , V. mimicus , V. furnisii и V. fortis , разработанные праймеры и зонд показали 100%-ную специфичность по отношению к штаммам V. parahaemolyticus (таблица).
Результаты идентификации клинических штаммов V. parahaemolyticus методом Real-Time ПЦР
|
Вид микроорганизма |
Число ис-следованных штаммов |
Результаты ПЦР |
|
|
число положи-тельных проб |
число от-рица-тельных проб |
||
|
V. parahaemolyticus |
100 |
100 |
0 |
|
V. alginolyticus |
25 |
0 |
25 |
|
V. fluvialis |
10 |
0 |
10 |
|
V. hollisae |
5 |
0 |
5 |
|
V. vulnificus |
10 |
0 |
10 |
|
V. harvey |
5 |
0 |
5 |
|
V. mimicus |
5 |
0 |
5 |
|
V. furnisii |
5 |
0 |
5 |
|
V. fortis |
2 |
0 |
2 |
Флуоресцентный сигнал с длиной волны 605 нм, характерной для флуорофора Rox, детектировался в ПЦР выше порогового значения тогда, когда пробы содержали ДНК парагемолитических вибрионов, и не детектировался в случае содержания в пробах ДНК других представителей рода Vibrio (таблица). При исследовании чистых культур бактериальных суспензий положительный результат нами наблюдался при концентрации клеток 109–105 м. кл./мл. При этом наблюдалась зависимость количества клеток и номера цикла (начала реакции): 15-й цикл при 109 м. кл./мл и 29-й при 105 м. кл./мл. (рисунок).
Таким образом, в результате проведенной нами работы были сконструированы специфичные праймеры и зонд к ДНК гена металлопротеазы (коллагеназы) V. parahaemolyticus . Были подобраны оптимальные условия проведения полимеразной цепной реакции с выбранной парой праймеров и флуоресцентным зондом в режиме «реального времени». Использование предполагаемого способа выявления специфического участка ДНК гена металлопротеазы (коллагеназы) V. parahae-molyticus с помощью ПЦР в режиме «реального времени» позволит быстро, точно и эффективно проводить идентификацию представителей вида V.
parahaemolyticus и дифференцировать их от близкородственных видов.
Следующим этапом нашей работы было определение возможности использования сконструированных нами праймеров для индикации штаммов V. parahaemolyticus в биологическом материале. Для этого использовали пробы рыбы, искусственно контаминированные бактериями V. parahae-molyticus. В результате наблюдали наличие флуоресценции по каналу ROX в пробах, содержащих (с учетом разведений) 107–105 м. кл./мл. При этом реакция начиналась позже, чем при постановке ее с чистой культурой микроорганизмов: 107 – 27-й цикл, 105 – 31-й цикл (рисунок).
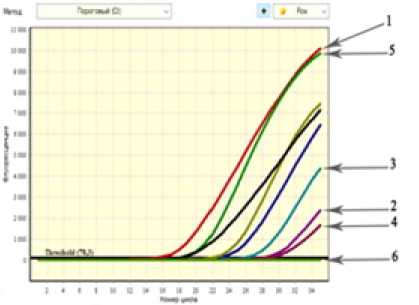
Зависимость флуоресценции канала ROX и номера цикла от состава пробы:
-
1 – концентрация клеток чистой культуры V. para-haemolyticus 109 м. кл./мл; 2 – концентрация клеток чистой культуры V. parahaemolyticus 105 м. кл./мл; 3 – концентрация клеток V. parahaemolyticus в контаминированной пробе рыбы, 107 м. кл./мл; 4 – концентрация клеток V. parahaemolyticus в контаминированной пробе рыбы, 105 м. кл./мл; 5 – положительный контроль; 6 – отрицательный контроль
Проведенные нами исследования показали возможность использования полученных праймеров, специфичных последовательности генов металло-протеазы (коллагеназы) – vppC V. parahae-molyticus, для индикации парагемолитических вибрионов в зараженных пробах рыбы. Чувствительность реакции составила 105 м. кл. в 1 г зараженного продукта.
При изучении возможности использования среды обогащения (1%-ная пептонная вода с 2%-ным NaCl) для накопления культуры в пробах, содержащих V. parahaemolyticus в количестве 101–104 м. кл./мл было показано, что после «подращивания» проб, изначально содержащих 104 м. кл./мл, в течение 4 ч. в среде обогащения количество колоний на плотной агаровой среде составило 105 м. кл./мл; пробы, изначально содержащие 101 м. кл./мл, достигли концентрации 105 м. кл./мл через 8 ч. выращивания в среде обогащения. После проведения ПЦР в режиме «реального времени» через 8 ч. выращивания наблюдали наличие флуоресценции по каналу ROX в пробах, изначально содержащих 101 м. кл./мл.
Использование метода Real-Тime ПЦР с предложенными нами праймерами, позволяет существенно повысить достоверность ПЦР-анализа, исключив возможность контаминации; сократить трудозатраты и время анализа, задействовав в аналитическом процессе минимум сотрудников; проводить ПЦР-анализ в одном помещении, что играет существенную роль для лабораторий, занимающихся исследованиями объектов рыбного промысла.
Выводы
Набор полученных нами олигонуклеотидов, включающий прямые, обратные праймеры и флуоресцентно-меченый зонд, сконструированные на основе фрагментов гена металлопротеазы (коллагеназы), специфичного V. parahaemolyticus , позволяет методом ПЦР с гибридизационно-флуоресцентным учетом результатов в режиме реального времени выявлять ДНК возбудителей V. pa-rahaemolyticus в исследуемом материале.
Применение разработанного набора реагентов для идентификации штаммов вида V. parahaemo-lyticus методом полимеразной цепной реакции в режиме реального времени (Real-Time PCR) при исследовании проб биологического материала и объектов окружающей среды позволяет обнаружить и дифференцировать ДНК возбудителей V. parahaemolyticus с аналитической чувствительностью от 1×105 м. к./мл и аналитической специфичностью – 100%.
Предварительное обогащение проб в 1%-ной пептонной воде с 2%-ным NaCl в течение 8 ч. позволяет выявлять бактерии V. parahaemolyticus при исходной концентрации их в пробе 101 м. кл./мл методом Real-Time PCR.
Список литературы Разработка способа индикации и идентификации Vibrio parahaemolyticus с помощью real-time ПЦР
- Белькова Н.Л., Дзюба Е.В., Суханова Е.В. Молекулярно-генетическая идентификация кишечной микрофлоры и протистов байкальских рыб // Аннотированный список фауны оз. Байкал и его водосборного бассейна. Новосибирск: Наука, 2009. С. 957-980.
- Белькова Н.Л. и др. Молекулярно-генетический мониторинг ассоциированной микрофлоры лососевидных рыб: разнообразие и физиологический статус // Известия Самарского научного центра РАН. 2010. Т. 12, № 1(4). С. 1108-1114.
- Домницкий И.Ю. Ихтиопатология: краткий курс лекций. Саратов, 2016. 120 с.
- Лабораторная диагностика заболеваний, вызываемых парагемолитическими и другими патогенными для человека вибрионами: МУК 4.2.1793-03. М.: Федеральный центр госсанэпиднадзора Минздрава России, 2004. 26 с.
- Организация работы лабораторий, использующих методы амплификации нуклеиновых кислот при работе с материалом, содержащим микроорганизмы I-IV групп патогенности: МУ 1.3.2569-09. М.: Федеральный центр гигиены и эпидемиологии Роспотребнадзора, 2010. 51 c.
- Полеева М.В., Чемисова О.С. Использование масс-спектрометрического анализа для детекции бактериальных токсинов // ЖМЭИ. 2018. № 1. С. 93-101.
- Ребриков Д.В. и др. ПЦР в реальном времени. М: Бином. Лаборатория знаний, 2019. 223 с.
- Способ идентификации штаммов вида Vibrio paraharmolyticus методом ПЦР в режиме реального времени: пат. № 2016146441 Рос. Федерации, 2016. № 2644232; заявл. 08.02.2018. Бюл. № 4 / Чемисова О.С., Трухачев А.Л., Рыковская О.А., Полеева М.В.
- Чемисова О.С. и др. Масс-спектрометрический анализ как метод идентификации и внутривидовой дифференциации Vibrio parahaemolyticus // Холера и патогенные для человека вибрионы: материалы проблемной комиссии. Ростов-н/Д, 2013. Вып. 26. С. 153-157.
- Buller N.B. Bacteria from fish and other aquatic animals: a practical identification manual. Oxfordshire: CABI publishing, 2004. 361 p.
- Chakraborty S., Nair G.B., Shinoda S. Pathogenic vibrios in the natural aquatic environment // Rev. Environ. Health. 1997. Vol. 12 (2). P. 63-80.
- Daniels N.A.et al. Emergence of a new Vibrio parahaemolyticus serotype in raw oysters: A prevention quandary // J. Am. Med. Assoc. 2000. Vol. 284 (12). P. 1541-1545.
- Deepanjali A. et al. Seasonal variation in abundance of total and pathogenic Vibrio parahaemolyticus bacteria in oysteralong the southwest coast of India // Appl. Environ. Microbiol. 2005. Vol. 71 (7). P. 3575-3580.
- Di Pinto A., Ciccarese G., Tantillo G. A collagenase-targeted multiplex PCR assay for identification of Vibrio alginolyticus, Vibrio cholerae, and Vibrio parahaemolyticus // J. of Food Protection. 2005. Vol. 68, № 1. P. 150-153.
- Hara-Kudo Y. et al. Improved method for detection of Vibrio parahaemolyticus in seafood // Appl. Environ Microbiol. 2001. Vol. 67 (12). P. 5819-5823.
- Li R. et al. A Novel PCR-Based Approach for Accurate Identification of Vibrio parahaemolyticus // Front Microbiol. 2016. Vol. 7. P. 44.
- Luan X.-Y. et al. Comparison of different primers for rapid detection of Vibrio parahaemolyticus using the polymerase chain reaction // Lett. Appl. Microbiol. 2007. Vol. 44 (3). P. 242-247.
- Reham A.A., Amani M.S. Specific Detection of Pathogenic Vibrio species in shellfish by using multiplex polymerase chain reaction // J. Global Veterinaria. 2012. Vol. 8, № 5. P. 525-531.
- Spanggaard B. et al. The microflora of rainbow trout intestine: a comparison of traditional and molecular identification // Aquaculture. 2000. Vol. 182. P. 1-15.
