Разработка стратегии индукции АОХ1 промотора при культивировании метилотрофных дрожжей Komagataella phaffii

Автор: Бытяк Д.С., Корнеева О.С., Мотина Е.А.
Журнал: Вестник Воронежского государственного университета инженерных технологий @vestnik-vsuet
Рубрика: Пищевая биотехнология
Статья в выпуске: 1 (87), 2021 года.
Бесплатный доступ
В настоящее время наблюдается значительное повышение интереса к промышленному производству ферментных препаратов (и других рекомбинантных белков) при помощи различных микроорганизмов, в том числе метилотрофных дрожжей, таких как Komagataellaphaffii. При этом наиболее значимая продуктивность целевых белков достигается при метанольной индукции гетерологичных генов, клонированных под контролем AOX1 промотора. Таким образом эффективность биосинтеза во многом определяется метаболизмом метанола. В связи с чем целью работы является разработка оптимальной стратегии метанольной индукции AOX1 промотора Komagataella phaffii.Объектом исследования является культура рекомбинантного штамма-продуцента фосфолипазы А2 Komagataellaphaffii. Исследования проводились в лабораторном ферментере InforsMinifors (Швейцария) на жидкой питательной среде BSM (BasalSaltMedium). В работе применяли общепринятые методы исследований характеристик метаболической активности, в том числе расчета удельных характеристик и продуктивности штамма. Результатом исследования является определение удельной скорости потребления метанола, используемого в качестве источника углерода, которая составила 19,2±1,8 мг/г*ч. Также в ходе выполнения работы определена удельная скорость роста Komagataella phaffii и составила 0,24 ч-1. На основании данных, полученных в ходе исследований разработана стратегия индукции AOX1 промотора при культивировании метилотрофных дрожжей Komagataellaphaffii, путем поддержания концентрации метанола в диапазоне от 0,6 до 2 % на основании концентрации растворенного кислорода в среде. Разработанная стратегия индукции AOX1 промотора позволила обеспечить получение не менее 1,87 г/л рекомбинантного белка (фосфолипаза А2) при культивировании Komagataella phaffii на протяжении 96 ч, что в 3,7 раз превосходит известные результаты.
Фосфолипаза а2, генная инженерия, биотехнология ферментов, экспрессия, aox промотор
Короткий адрес: https://sciup.org/140257311
IDR: 140257311 | УДК: 579.66 | DOI: 10.20914/2310-1202-2021-1-115-120
Development of a strategy induction AOX1 promoter methylotrophic yeast Komagataella phaffii
Currently, there is a significant increase in interest in the industrial production of enzyme preparations (and other recombinant proteins) using various microorganisms, including methylotrophic yeasts such as Komagataella phaffii. At the same time, the most significant productivity of the target proteins is achieved by methanol induction of heterologous genes cloned under the control of the AOX1 promoter. Thus, the efficiency of biosynthesis is largely determined by the metabolism of methanol. In this connection, the aim of the work is to develop an optimal strategy for methanol induction of the AOX1 promoter of Komagataella phaffii. The object of the study is the culture of the recombinant phospholipase A2 producing strain Komagataella phaffii. The studies were carried out in a laboratory fermenter Infors Minifors (Switzerland) on a liquid nutrient medium BSM (Basal Salt Medium) We used the generally accepted methods of studying the characteristics of metabolic activity, including the calculation of specific characteristics and productivity of the strain. The result of the study is the determination of the specific rate of consumption of methanol used as a carbon source, which was 19.2±1.8 mg/g*h. Also, the specific growth rate of Komagataella phaffii was determined and amounted to 0.24 h-1.Based on the data obtained during the research, a strategy for the induction of the AOX1 promoter in the cultivation of the methylotrophic yeast Komagataella phaffii was developed by maintaining the methanol concentration in the range of 0.6 to 2% based on the concentration of dissolved oxygen in the medium. The developed strategy of induction of the AOX1 promoter made it possible to obtain at least 1.87 g / l of recombinant protein (phospholipase A2) during cultivation of Komagataella phaffii for 96 h, which is 3.7 times higher than the known results.
Текст научной статьи Разработка стратегии индукции АОХ1 промотора при культивировании метилотрофных дрожжей Komagataella phaffii
Дрожжевые системы экспрессии, в том числе Komagataellaphaffii способны к эффективной секреции рекомбинантных белков непосредственно в культуральную жидкость, что значительно облегчает последующие стадии очистки целевых белков от сопутствующих примесей и вторичных метаболитов [1, 7, 15].
Наибольший уровень секреции рекомбинантных белков при культивировании Komaga-taellaphaffii , до 30% от тотальной фракции белка достигается при метанольной индукции гетерологичных генов, клонированных под контролем АОХ1 промотора, однако точный механизм регуляции промотора АОХ1 до сих пор не изучен [2, 5, 8].
Важно отметить, что АОХ1 промотор репрессируется в присутствии глюкозы или глицерина, что позволяет разделить процесс культивирования на стадию набора биомассы (при наличии в среде репрессирующего источника углерода) и стадию синтеза целевого белка (при наличии в среде метанола) [3, 6, 11, 19].
Цель работы – разработка оптимальной стратегии метанольной индукции АОХ1 промотора Komagataellaphaffii .
Результаты и обсуждение
В ходе выполнения настоящей работы исследована скорость роста биомассы и потребления глицерина, при культивировании Komagataella phaffii в лабораторном ферментере Infors Minifors ( Швейцария ) на жидкой питательной среде BSM (Basal Salt Medium) при следующих параметрах: температура 30 ± 0,1 ºС; рН 6 ± 0,2; скорость вращения мешалки 900 ± 10 об/мин; скорость подачи воздуха 1,7 ± 0,05 л/л/мин; давление 0,05 ± 0,01 МПа.
В процессе культивирования обеспечивалось поддержание концентрации глицерина в диапазоне от 0,5 до 2%, за счет периодического внесения подпиточного раствора, содержащего глицерин и микроэлементы PTM. Избыток источника углерода обеспечивает длительную Log-фазу, характеризующуюся линейной скоростью роста биомассы [4, 9, 10]. В результате чего была определена удельная скорость роста Komagataella phaffii и составила 0,24 ч-1.
Расчет удельной скорости роста Komagataella phaffii проводился по формуле:
X – X
^ = 0Х
X 0 ( t 1 t 0 )
где µ – удельная скорость роста, ч-1; Х 0 – начальная концентрация биомассы, г/л; Х 1 – конечная концентрация биомассы, г/л; t 0 – начальный момент времени, ч; t 1 – конечный момент времени, ч.
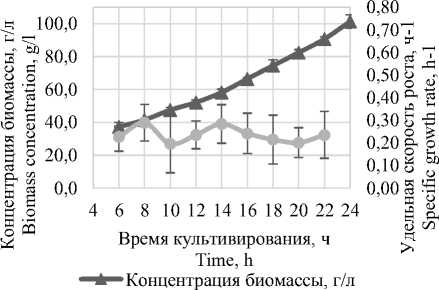
Удельная скорость роста, ч-1
Рисунок 1. Удельная скорость Komagataella phaffii
Figure 1. Specific growth rate Komagataella phaffii
Также проведен расчет удельной скорости потребления глицерина, которая составила 36,99 мг/г×ч. Однако важно учитывать, что скорость потребления глицерина может варьироваться в достаточно широком диапазоне. Рассчитанный показатель устанавливает максимальную скорость потребления глицерина. При этом потенциал роста биомассы ограничивается не исчерпанием питательных веществ, а недостатком растворенного кислорода, при достижении концентрации биомассы более 300 г./л (в пересчете на влажную биомассу). Недостаток кислорода объясняется спецификой массообменных характеристик в заданных условиях ведения процесса [12, 20].
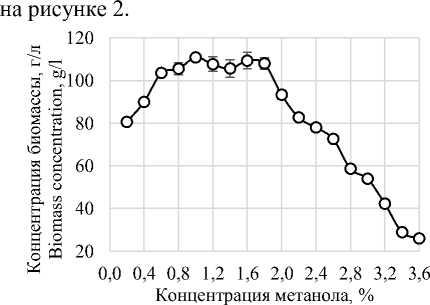
Исследование влияния массовой доли метанола на жизнедеятельность Komagataella phaffii проведено путем культивирования штамма на среде BSM в колбах, с градиентной концентрацией метанола, в диапазоне от 0,2% до 3,6% с шагом 0,2%. Установлено, что при накоплении метанола в культуральной среде более 2% наблюдается значительное снижение скорости роста культуры, что наглядно отражено
Methanol concentration, %
Рисунок 2. Влияние концентрации метанола на прирост биомассы Komagataella phaffii
Figurе2. Effect of methanol concentration on Komagataella phaffii biomass growth
Полученные данные также свидетельствуют о том, что при концентрации метанола менее 0,6% наблюдается снижение скорости прироста биомассы, что вероятно, связано с лимитирующим количеством метанола в среде, в связи с чем особенно важно поддерживать оптимальную концентрацию метанола. Для поддержания оптимальной концентрации метанола был проведен расчет потребления метанола Komagataella phaffii при культивировании в лабораторном ферментере Infors Minifors ( Швейцария ), полученные результаты отражены на рисунке 3.
Q о]
О О СЦ
-
сл О
Время культивирования, ч | Time. h
Концентрация метанола % концентрация биомассы г/л
Концентрация pO2, %
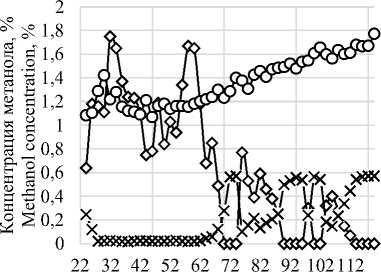
Рисунок3. Динамика потребления метанола
Figurе 3. Dynamics of methanol consumption
Полученные данные свидетельствуют о том, что скорость потребления метанола в процессе культивирования варьируется в зависимости от физиологического состояния культуры, наличия или отсутствия определенных компонентов среды и накопления целевого продукта в диапазоне от 0 до 11,256 мкл/ч/г [13, 18].
Расчет скорости потребления метанола Komagataellaphaffii проводился по формуле:
S – S *10
иМ=—---0---х 1000.
C*t где μM – скорость потребления метанола, мкл/ч/г; S0 – начальная концентрация метанола, %; Sx – конечная концентрация метанола, %;
-
10 – коэффициент пересчета% в мл/л; C – средняя концентрация биомассы, г/л; t – время исследования, ч; 1000 – коэффициент пересчета в мкл/г.
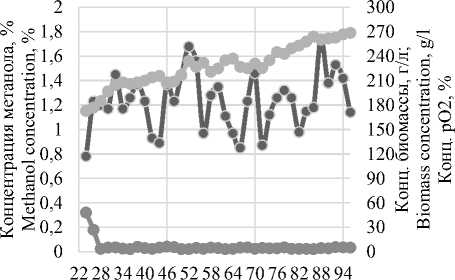
При этом из рисунка 3 видна прямая зависимость между массовой долей метанола в среде и концентрацией растворенного кислорода, рО2, в следствии чего в ПО Irisstandart была разработана рецептура внесения метанола на основании показаний датчика рО2, в соответствии с которой при достижении концентрации растворенного кислорода в среде 20% и менее активировался перистальтический насос со скоростью не менее 3 мл/ч на литр среды, до момента достижения концентрации рО2 80%, но не более 20 мл/ч. Полученные данные представлены на рисунке 4.
Время культивирования, ч | Time, h Концентрация метанола % концентрация биомассы г/л Концентрация pO2, %
Рисунок 4. Апробация стратегии внесения метанола
Figurе 4. Testing the methanol application strategy
Также в ходе выполнения настоящей работы проведено исследование зависимости эффективности синтеза целевого белка от концентрации биомассы на момент начала индукции. Исследование проводилось в диапазоне начальных концентраций биомассы от 100 до 300 г./л. В ходе выполнения работы установлено, что оптимальной концентрацией биомассы на момент начала внесения индуктора является 150 ± 10 г./л [14, 16, 17]. Полученные данные представлены в таблице № 1.
Таблица 1.
Концентрации биомассы на момент начала индукции
Table 1.
Biomass concentrations at the start of induction
|
Концентрация биомассы, г/л Biomass concentration, g/l |
Концентрация целевого белка, г/л Target protein concentration, g / l |
Концентрация биомассы, г/л Biomass concentration, g/l |
Концентрация целевого белка, г/л Target protein concentration, g / l |
|
101,7 |
0,96 |
201 |
1,03 |
|
120,4 |
1,23 |
221,4 |
0,86 |
|
142 |
1,74 |
243 |
0,53 |
|
160,6 |
1,85 |
262,3 |
0,32 |
|
179,7 |
1,37 |
278,9 |
0,18 |
|
300,3 |
0,13 |
||
Концентрация биомассы на момент начала индукции более 160,6 г/л приводит к снижению времени индукции за счет более быстрого достижения состояния дефицита кислорода и отмирания биомассы, что сокращает длительность стадии индукции и как следствие приводит к снижению выхода целевого белка. При концентрации биомассы на момент начала индукции менее 142 г./л также наблюдается снижение выхода целевого белка, что наиболее вероятно связано с неэффективным использованием компонентов питательной среды ввиду недостаточности концентрации биомассы, а также увеличением общего времени видения процесса, что также приводит к повышению отмирания биомассы и дополнительным механическим повреждениям.
На основании полученных данных и ранее полученных значениях удельной скорости потребления метанола (19,2 ± 1,8 мг/г×ч), получена формула расчета требуемого количества метанола:
M = X0qm (1) 1000Vk где Х0 – начальная концентрация биомассы, г/л; qm – удельная скорость потребления метанола, мг/ г×ч; V – рабочий объем питательной среды в ферментере, л; kt – периодичность внесения индукционного раствора, ч-1; 1000 – перевод в г/г×ч.
Важно отметить, что индукция АОХ1 промотора и как следствие синтез целевого белка начинается только после исчерпания в среде глицерина. При этом перестройка метаболизма с глицерина на метанол занимает ориентировочно 4 часа [7, 12].
Заключение
В результате выполнения настоящей работы разработана стратегия индукции АОХ1 промотора при культивировании метилотроф-ных дрожжей Komagataella phaffii , путем поддержания концентрации метанола в диапазоне от 0,6 до 2% на основании концентрации растворенного кислорода в среде.
Разработанная стратегия индукции АОХ1 промотора позволила обеспечить получение не менее 1,87 г./л рекомбинантного белка (фосфолипаза А2) при культивировании Komagataella phaffii на протяжении 96 ч, что в 3,7 раз превосходит результаты, полученные при ведении процесса в соответствии с руководством «Pichia Fermentation Process Guidelines» от Invitrogen.
Список литературы Разработка стратегии индукции АОХ1 промотора при культивировании метилотрофных дрожжей Komagataella phaffii
- Zavec D., Troyer C., Maresch D., Altmann F. et al. Beyond alcohol oxidase: the methylotrophic yeast Komagataella phaffii utilizes methanol also with its native alcohol dehydrogenase Adh2 // FEMS yeast research. 2021. V. 21. №2. 2. P. foab009. doi: 10.1093/femsyr/foab009
- Vogl Т., Fischer .Т.Е., Hyden P., Wasmayer R. et al. Orthologous promoters from related methylotrophic yeasts surpass expression of endogenous promoters of Pichia pastoris//AMB Express. 2020. V. 10. №. l.P. 1-9. doi: 10.1186/sl3568-020-00972-l
- Weninger A., Hatzl, A.M., Schmid C., Vogl T. et al. Combinatorial optimization of CRISPR/Cas9 expression enables precision genome engineering in the methylotrophic yeast Pichia pastoris // Journal of biotechnology. 2016. V. 235. P. 139-149. doi: 10.1016/j.jbiotec.2016.03.027
- Ohsawa S., Nishida S., Oku M., Sakai Y. et al. Ethanol represses the expression of methanol-inducible genes via acetyl-CoA synthesis in the yeast Komagataella phaffii//Scientific reports. 2018. V. 8. №. 1. P 1-11. doi: 10.1038/s41598-018-36732-2
- García-Ortega X., Cámara E., Ferrer P., Albiol J. et al. Rational development of bioprocess engineering strategies for recombinant protein production in Pichia pastoris (Komagataella phaffii) using the methanol-free GAP promoter. Where do we stand? //Newbiotechnology. 2019. V. 53. P. 24-34. doi: 10.1016/j.nbt.2019.06.002
- Ivanova A.V., Sidorin A.V., Sambuk E.V., Rumyantsev AM. et al. The analysis of the polyamine oxidase genes in the methylotrophic yeast Komagataella phaffii // Ecological genetics. 2019. V. 17. №. 4. P. 47-55. doi: 10.17816/ecogen17447-55
- Dalvie N.C., Leal J., Whittaker C.A., Yang Y. et al. Host-informed expression of CRISPR guide RNA for genomic engineering in Komagataella phaffii //ACS synthetic biology. 2019. V. 9. №. 1. P. 26-35. doi.: 10.1021/acssynbio.9b00372
- Barrero J.J., Casler J.C., Valero F., Ferrer P. et al. An improved secretion signal enhances the secretion of model proteins from Pichia pastoris//Microbial cell factories. 2018. V. 17. №. 1. P. 1-13. doi: 10.1186/sl2934-018-1009-5
- Cámara E., Landes N., Albiol J., Gasser B. et al. Increased dosage of AOX1 promoter-regulated expression cassettes leads to transcription attenuation of the methanol metabolism in Pichia pastoris // Scientific reports. 2017. V. 7. №. 1. P. 1-16. doi: 10.1038/srep44302
- Jordá J., JouhtenP., Cámara E., Maaheimo H. et al. Metabolic flux profiling of recombinant protein secreting Pichia pastoris growing on glucose: methanol mixtures//Microbial cell factories. 2012. V. 11. №. 1. P. 1-14. doi: 10.1186/1475-2859-11-57
- Rußmayer H., Buchetics M., Gruber С., Valli M. et al. Systems-level organization of yeast methylotrophic lifestyle //BMC biology. 2015. V. 13. №. 1. P. 1-25. doi: 10.1186/sl2915-0Í5-0186-5
- Murakami M., Taketomi Y., Sato H., Yamamoto K. Secreted phospholipase A2 revisited // The Journal of Biochemistry. 2011. V. 150. №. 3. P. 233-255. doi: 10.1093/jb/mvr088
- Murakami M., Sato H., Miki Y., Yamamoto K. et al. A new era of secreted phospholipase A2 // Journal of lipid research. 2015. V. 56. №. 7. P. 1248-1261. doi: 10.1194/jlr.R058123
- Seresht A.K., NorgaardP., Palmqvist E.A., Andersen A. S. et al. Modulating heterologous protein production in yeast: the applicability of truncated auxotrophic markers // Applied microbiology and biotechnology. 2013. V. 97. №. 9. P. 3939-3948. doi: 10.1007/s00253-012-4263-1
- Rosenson R.S., Hurt-Camejo E. Phospholipase A2 enzymes and the risk of atherosclerosis // European heart journal. 2012. V. 33. №. 23. P. 2899-2909. doi: 10.1093/eurheartj/ehsl48
- Idiris A., Tohda H., Kumagai H., Takegawa K. Engineering of protein secretion in yeast: strategies and impact on protein production//Applied microbiology and biotechnology. 2010. V. 86. №. 2. P. 403-417. doi: Í0.1007/s00253-010-2447-0
- Wang J., Wang X., Shi L., Qi F. et al. Methanol-independent protein expression by AOX1 promoter with trans-acting elements engineering and glucose-glycerol-shift induction in Pichia pastoris // Scientific reports. 2017. V. 7. №. 1. P. 1-12. doi: 10.1038/srep41850
- Самойлова К).В., Тулупов A.A., Сорокина К.Н. Современные подходы к разработке новых ферментов липаз и продуцентов на их основе // Вестник Новосибирского государственного университета. Серия: Биология, клиническая медицина. 2014. Т. 12. № 4.
- Корнеева О.С. Анненков В.А. Получение ферментного препарата липазы методом рекомбинантных ДНК // Технологии пищевой и перерабатывающей промышленности АПК - продукты здорового питания. 2014. № 3.
- Пат. № 2501014, RU, G01N 33/50, 33/48. Способ определения липолитической активности в субклеточных фракциях бактерий / Козлов С.Н., Марков Е.Ю., Николаев В.Б., Урбанович Л.Я. и др. №2012132418/15; Заявл. 15.08.2012; Опубл. 10.12.2013, Бюл. № 34.
