Реакция генотипов сои с различной структурой продуктивности на изменения условий и дифференцирующая способность среды

Автор: Розенцвейг В.Е., Голоенко Д.В., Давыденко О.Г.
Статья в выпуске: 1 (140), 2009 года.
Бесплатный доступ
Изучен вклад числа узлов главного побега и крупности семян в урожайность сои в группах спелости 00 - ранняя 0 в аспекте адаптивной селекции. Анализ связей урожайности с числом узлов и массой 1000 семян методом послойной регрессии в градиенте сред выявил в обоих случаях возрастание коэффициентов регрессии, а также дифференцирующей способности при повышении индекса среды. Это свидетельствует о предпочтительности большого числа узлов и крупносемянности для пластичного сорта. Установлены прогнозируемые прибавки продуктивности от оптимизации этих параметров в различных средах. Регрессия урожайности на число узлов, вероятно, носит нелинейный характер; более высокая отзывчивость изменения коэффициента регрессии на улучшение среды наблюдается в диапазоне сред с индексом 2-2,5 т/га. Наилучшей дифференцирующей способностью как по урожайности, так и по числу узлов характеризуется разреженный посев (20 раст./м2). Полученные результаты могут быть использованы для оптимизации селекционного процесса по сое.
Соя, группы спелости, число узлов, вклад в продуктивность, средовые индексы, коэффициенты регрессии
Короткий адрес: https://sciup.org/142150889
IDR: 142150889 | УДК: 631.52:633.853.52
Response of soybean genotypes differing in yield structure to ecological changes and dif-ferentiative capacity of environment
Breeding for specific adaptability requires estimation of quantitative traits inputs into yield response to environment alterations. About 200 genotypes of early soybean MGs 00 and early 0 were studied for 7 years in Belarus. The stratified regression analysis of „yield - node number‟ and „yield - TSW‟ interrelations in the gradient of environments revealed extension of regression slopes and differentiative capacity as index of environment increased in both cases. Thus, increasing node number and TSW promotes specific adaptability to high-yielding locations within MG 00. Predicted yield gains from the optimization of these traits were estimated for different environments. Regression of yield on node number is likely to be non-linear: regression coefficient was more responsive to environment index over the range of environments 2 to 2.5 t ha-1. Best differentiative capacity for both yield and node number was observed in sparse crops (20 plants m-2). Obtained results may be used for optimizing breeding process for early soybeans.
Текст научной статьи Реакция генотипов сои с различной структурой продуктивности на изменения условий и дифференцирующая способность среды
Для организации селекции на специфическую адаптивную способность необходим градиент экологических сред, обеспечивающий выявление характера реакций генотипов на изменение условий среды [1, 4]. При этом информативность анализа поведения генотипов определяется использованием корректных статистических показателей [2]. Другую сторону проблемы составляет потребность в диагностике признаков, вносящих в данной среде наибольший вклад в продуктивность. Для их эффективной идентификации необходимо наличие (или искусственное создание) сред с высокой дифференцирующей способностью [5]. О практическом опыте создания такой системы агрофонов в селекции сои сообщает Мордвинцев [7]. Так, в селекционном процессе Ершовской ОСОЗ функционирует ряд агрофонов, переменными среды в которых являются полив, срок посева, плотность стеблестоя и др.
Наиболее значительный вклад в продуктивность растения сои в группе спелости 00 вносит число узлов главного побега. Напротив, масса 1000 семян в достаточно широком диапазоне значений признака, согласно оценкам большинства исследователей, не оказывает влияния на продуктивность сои [6, 18, 11, 8, 9]. Корреляционный анализ предоставляет информацию о наличии, направленности и силе связи признаков, однако результативность селекции на урожайность по какому-либо признаку будет определяться прибавкой, достигнутой за счет его оптимизации. Предпринимались попытки оценить прибавки урожайности, обусловленные динамикой того или иного элемента продуктивности, по коэффициенту детерминации R2, который отражает долю вариации зависимого признака, обусловленную независимым [10]. Однако этот метод не является вполне корректным, и предпочтительнее оценивать прибавки продуктивности методом «послойной регрессии» [20]. В контексте этой публикации послойная регрессия представляет систему уравнений регрессии некоторого фактора А (в данном случае, урожайности) на фактор В (элементы продуктивности) на разных уровнях значений фактора С (параметры среды: экологический индекс, сумма осадков и др.). Целью исследования является анализ зависимости урожайности от числа узлов и массы 1000 семян в различных средах и идентификация селекционных сред с наилучшей дифференцирующей способностью.
Материал и методика. В настоящей работе проанализированы данные изучения сортов сои групп спелости 000, 00 и ранней 0 коллекционного питомника компании «Соя-Север» за 2002-2008 гг. Питомник размещался в Лунинецком р-не Брестской обл. Беларуси (52○ с.ш., почва супесчаная, подстилаемая песком, гумус 1,4 %). Генотипы изучались при плотности стеблестоя к уборке 45-50 раст./м2 (рекомендуемая плотность в производстве), а в 2008 г. параллельно и при плотности 20 раст./м2 (далее разреженный посев). Объем выборки составил более 200 вариант. Статистическая обработка данных проводилась с помощью программ MS Excel и Statistica 6.0. Дисперсионный анализ дифференцирующей способности среды (ДСС) осуществлен по методике Кильчевского и Хотылевой [5]. Ввиду значительного различия средовых индексов использовался показатель относительной дифференцирующей способности:
S ek = a2 ДССk x 100% / (u+d k ) = «з1 ДССk X 100% / I j .
Вклад числа узлов главного побега в урожайность. Регрессионная зависимость средней многолетней (2002-2008) урожайности (УР) от количества узлов (КУ) выражалась линейным уравнением:
УР(%) = 6,6 X КУ + 16,0; R2=0,3. (1)
Урожайность здесь и далее выражена в процентах (%) от стандарта. В таблице 1 представлены уравнения регрессии для этой же связи по годам (строки ранжированы по значениям индекса среды Ij ).
Таблица 1 – Регрессия урожайности на число узлов главного побега на фоне изменения индекса среды
|
Год |
I j , т/га |
УР(%) = b X КУ + а |
R2 |
|
2002 |
1,04 |
Y = 4,64x + 36,9 |
0,14 |
|
2004 |
2,08 |
Y = 4,98x + 35,2 |
0,23 |
|
2005 |
2,45 |
Y = 5,69x + 25,5 |
0,19 |
|
2008 |
2,54 |
Y = 7,13х + 16,5 |
0,34 |
|
2007 |
2,55 |
Y = 5,13x + 36,9 |
0,16 |
|
2006 |
3,11 |
Y = 5,94x + 30,4 |
0,28 |
|
2008 (20 р./м2) |
- |
Y = 9,15х – 42,7 |
0,59 |
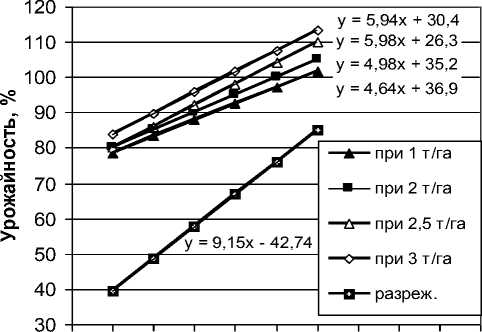
Как видно из таблицы 1 и уравнения 1, в условиях производственного посева каждый дополнительный узел обеспечивал от 4,6 до 7,1 % средней прибавки урожайности, в зависимости от индекса среды, а в разреженном посеве – 9,2 %. Корреляция урожайности с числом узлов в разреженном посеве была очень высокой (0,77), тогда как в плотном колебалась от 0,37 до 0,58 (все коэффициенты значимы на уровне a<0,001). Связь коэффициента регрессии урожайности на число узлов с индексом среды была положительной (коэффициент ранговой корреляции Спирмена r s = 0,71 значим на уровне a<0,11).
Для дальнейшего анализа данные были сгруппированы по уровням индекса среды (округлены до 1,0; 2,0; 2,5 и 3,0 т/га) и рассчитана прогнозируемая урожайность генотипов в зависимости от числа узлов (табл. 2).
Таблица 2 – Прогнозируемая урожайность (% от среднего многолетнего стандарта) сорта сои в зависимости от числа узлов главного побега и индекса среды
|
Год |
Усредненные уровни I j , т/га |
Коэффициенты регрессионной модели |
Число узлов главного побега |
||||||
|
b |
a |
9 |
10 |
11 |
12 |
13 |
14 |
||
|
2002 |
1,04 |
4,64 |
36,9 |
78,7 |
83,3 |
87,9 |
92,6 |
97,2 |
101,9 |
|
2004 |
2,08 |
4,98 |
35,2 |
80,0 |
85,0 |
90,0 |
95,0 |
99,9 |
104,9 |
|
2005, 2007, 2008 |
2,51 |
5,98 |
26,3 |
80,1 |
86,1 |
92,1 |
98,1 |
104,0 |
110,0 |
|
2006 |
3,11 |
5,94 |
30,4 |
83,9 |
89,8 |
95,7 |
101,7 |
107,6 |
113,6 |
|
2008 (20 р./м2) |
- |
9,15 |
–42,74 |
39,6 |
48,8 |
57,9 |
67,1 |
76,2 |
85,4 |
|
Прибавка, % (фон 3 т/га относительно 1 т/га) |
6,6 |
7,8 |
8,9 |
9,8 |
10,7 |
11,5 |
|||
Таким образом, при улучшении среды от крайне экстенсивной (Ij ≈ 1 т/га) до интенсивной (Ij ≈ 3 т/га) короткостебельные сорта с 9-10 узлами повышают свою урожайность относительно среднего многолетнего значения стандарта лишь на 6-8 %, тогда как высокорослые сорта с 13-14 узлами – на 11-12 %. При этом в каждой отдельно взятой среде сорта с большим числом узлов также значительно превосходят низкорослые сортообразцы (смены рангов генотипов не наблюдается). Поэтому для групп спелости 00 и ранней 0 в ранних питомниках (гибридный, однорядковые делянки в СП-1) целесообразно проводить отбор генотипов с большим количеством узлов как при селекции на общую адаптивную способность, так, в частности, и при создании интенсивных сортов с пластичной (т.е. отзывчивой на агрофон) урожайностью. Разумеется, при этом необходима тщательная проработка материала на устойчивость к полеганию. Графически полученные результаты представлены на рисунке 1.
Полученные результаты не согласуются с моделью короткостебельного детерминантного сорта, предложенной Р.Л. Купером для высокопродуктивных сред с ожидаемой урожайностью не менее 3,3 т/га [14, 15]. В 00 группе спелости основным лимитирующим элементом структуры продуктивности является число узлов главного побега, а его более высокие значения свойственны, в среднем, индетерминантному морфоти-пу. В климатических зонах с большими тепловыми ресурсами урожайность сои в продуктивных средах лимитируется, главным образом, полеганием, как показано Купером для групп спелости II–IV, поэтому в агроэкосистемах с высоким энергетическим вкладом (полив, удобрения) преимущество вполне могут иметь детерминантные сорта [19, 16]. Тем не менее, короткостебельный детерминантный морфотип, обладая высокой дисперсией специфической адаптивной способности, оказался весьма узкоспециализированным и чувствительным к засухе и плотности стеблестоя. По этим причинам детерминантным сортам не удалось занять значительных площадей в северной части США [13, 17].
8 9 10 11 12 13 14 15 16 17 18
Число узлов главного побега
Рисунок 1 – Послойная регрессия урожайности сортов сои на количество узлоглавного побега на разных уровнях индекса среды и в разреженном посеве
К проблеме создания дифференцирующего селекционного агрофона. Высокое значение коэффициента регрессии урожайности на число узлов в условиях разреженного посева свидетельствует о том, что данная среда может служить дифференцирующим фоном при селекции на продуктивность. При низкой плотности стеблестоя возрастает вероятность отбора генотипов, формирующих большое количество узлов главного побега, что опосредованно способствует выделению наиболее высокопродуктивных форм. Для проверки этой гипотезы были рассчитаны относительные дифференцирующие способности сред Sek (табл. 3). Очевидно, что в условиях разреженного посева происходит резкое увеличение ДСС как по числу узлов, так и по урожайности.
Таблица 3 – Относительная дифференцирующая способность сред по урожайности и количеству узлов главного побега в зависимости от индекса среды
|
Год |
Ij , т/га |
S ek по УР |
S ek по КУ |
|
2002 |
1,04 |
370 |
15,8 |
|
2004 |
2,08 |
456 |
13,9 |
|
2005 |
2,45 |
305 |
10,4 |
|
2008 |
2,54 |
606 |
16,6 |
|
2007 |
2,55 |
491 |
10,0 |
|
2006 |
3,11 |
704 |
13,6 |
|
2008 (20 р./м2) |
- |
1014 |
24,6 |
Реакция коэффициента регрессионной модели b на изменение индекса среды, возможно, носит нелинейный характер (см. табл. 2), хотя недостаточное число лет наблюдений не позволяет сделать достоверное заключение [1]. Так, при повышении I j с 1 до 2 т/га средняя прогнозируемая прибавка урожайности на каждый узел возрастает с 4,6 до 5,0 %, в интервале 2-2,5 т/га – с 5,0 до 6,0 %, а дальнейшее улучшение среды до уровня 3 т/га и выше не приводит к росту степени отзывчивости урожайности на число узлов (рис. 2). Если это справедливо, то прирост урожайности за счет селекции на количество узлов наиболее значим в диапазоне сред от 2 до 2,5 т/га. Повышение же урожайности с 1 до 2 т/га и с 2,5 до 3 т/га происходит преимущественно за счет агротехники, погоды и других факторов. ДСС по урожайности также возрастала по мере увеличения индекса среды (rs=0,73; a<0,1) и была положительно связана с коэффициентом регрессии урожайности на число узлов (r s =0,79; a<0,05).
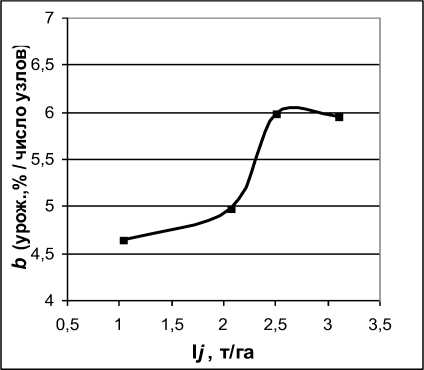
Рисунок 2 – Зависимость коэффициента регрессии урожайности сои в группе спелости 00 на число узлов от индекса среды
Вклад массы 1000 семян в пластичность урожайности. Полученные нами ранее данные о динамике связи крупности семян сои с урожайностью в градиенте сред [10] позволили сделать предварительный вывод о том, что крупносемянность может быть полезной только для интенсивного сорта. Анализ этой связи методом послойной регрессии (табл. 4) с последующей группировкой данных по уровням индекса среды и индекса распределения осадков (табл. 5, 6), в целом, подтвердил ранее опубликованные выводы, позволив уточнить прогнозируемые значения прибавок урожайности в различных средах.
Таблица 4 – Регрессия урожайности (УР) на массу 1000 семян (МТС при разных индексах среды)
|
Год |
I j , т/га |
УР(%) = b X МТС + а |
|
2002 |
1,04 |
Y = –0,220x + 126 |
|
2005 |
2,45 |
Y = 0,004x + 101 |
|
2008 |
2,54 |
Y = 0,130х + 75 |
|
2007 |
2,55 |
Y = –0,002x + 94 |
|
2006 |
3,11 |
Y = 0,090x + 77 |
Таблица 5 – Прогнозируемая урожайность (% от среднего многолетнего стандарта) сорта сои в зависимости от массы 1000 семян и индекса среды
|
Год |
Усредненные уровни I j , т/га |
Коэффициент регрессионной модели |
Масса 1000 семян, г |
||||||
|
b |
a |
140 |
160 |
180 |
200 |
220 |
240 |
||
|
2002 |
1,04 |
–0,220 |
126 |
95,2 |
90,8 |
86,4 |
82,0 |
77,6 |
73,2 |
|
2005, 2007, 2008 |
2,51 |
0,044 |
90 |
96,2 |
97,0 |
97,9 |
98,8 |
99,7 |
100,6 |
|
2006 |
3,11 |
0,090 |
77 |
89,6 |
91,4 |
93,2 |
95,0 |
96,8 |
98,6 |
Таблица 6 – Прогнозируемая урожайность (% от среднего многолетнего стандарта) сорта сои в зависимости от массы 1000 семян и индекса распределения осадков* (ИРО)
|
Год |
Уровни ИРО |
Коэффициенты регрессионной модели |
Масса 1000 семян, г |
||||||
|
b |
a |
140 |
160 |
180 |
200 |
220 |
240 |
||
|
2002 |
2 |
–0,220 |
126 |
95,2 |
90,8 |
86,4 |
82,0 |
77,6 |
73,2 |
|
2005,2007,2008 |
0 и –1 |
0,055 |
87 |
94,5 |
95,6 |
96,7 |
97,8 |
98,9 |
100,0 |
* принимается равным числу месяцев, в которые наблюдались симптомы дефицита влаги; при засухе в репродуктивной фазе (июль–август) ИРО>0, в вегетативной (май–июнь) ИРО<0 [9, 10]
Зарегистрированное, хотя и слабое, увеличение коэффициентов регрессии урожайности на массу 1000 семян по мере улучшения среды позволяет сделать вывод о целесообразности селекции на этот параметр только при создании интенсивных сортов для высоких агрофонов. Например, в случае увеличения массы 1000 семян со 160 до 220 г, средние прогнозируемые прибавки урожайности в благоприятных средах ( I j = 2,5-3 т/га; ИРО≤0) составят от 3 до 5 %. Однако в неблагоприятных средах ( I j ≈ 1 т/га; ИРО>0) эта селекционная стратегия приведет к снижению урожайности в рассматриваемом примере на 13 % (наблюдается смена рангов). Таким образом, селекция на увеличение массы 1000 семян ведет к повышению дисперсии специфической адаптивной способности (СУ САС Кильчевского и Хотылевой, 1997). Целесообразность данного направления селекции должна оцениваться с учетом климатических особенностей региона и уровня агротехники сельскохозяйственных производителей.
Отметим, что на современном этапе селекции генетические прибавки порядка 10 % и более за цикл уже труднодостижимы. Хотя значения рассматриваемых прибавок (3-5 %) в большинстве деляноч-ных опытов не превысят НСР05 и могут не быть достоверно зарегистрированы. Тем не менее, в период развитой селекции постепенное аккумулирование мелких прибавок является основным путем селекционного прогресса [13].
Таким образом, селекция на увеличение числа узлов и массы 1000 семян сои в группах спелости 00 – ранняя 0 приводит к повышению специфической адаптивной способности к продуктивным средам. Наилучшей дифференцирующей способностью характеризуются разреженные посевы и среды с высоким экологическим индексом.
