Реакция эритроцитов крыс на ступенчатую плавательную нагрузку и тест «до отказа» в термонейтральной среде

Автор: Монгалв Н.П., Вахнина Н.А., Бойко Е.Р.
Статья в выпуске: 4 т.256, 2023 года.
Бесплатный доступ
Целью работы явилось определение адаптационной реакции клеток красной крови у крыс после 40 дней возрастающей плавательной нагрузки до 4% от массы тела в термонейтральной среде в тесте «до отказа». Показано, что у животных в результате выполнения ступенчатой физической нагрузки двухкратное увеличение предельного времени плавания сопровождалось снижением концентрации гемоглобина в эритроците, отсутствием ретикулоцитоза и незначительным сдвигом распределения эритроцитов кривой Прайс-Джонса в диапазоне 6,0-6,3 мкм. Предполагаем, что наличие корреляции между количеством эритроцитов и их диаметром может свидетельствовать о проявлении механизма, регулирующего это соотношение. Изменения в эритроцитарном составе, связанные со снижением функциональной напряженности транспортной системы крови по сравнению с идентичной, но однократной нагрузкой в тесте «до отказа», могут являться маркерами физиологически обоснованного адаптивного ответа на ступенчатую физическую нагрузку.
Крысы, эритроциты, ретикулоциты, ступенчатая физическая нагрузка, термонейтральная среда, тест «до отказа»
Короткий адрес: https://sciup.org/142239871
IDR: 142239871 | УДК: 599.323.45: | DOI: 10.31588/2413_4201_1883_4_256_178
Response of erythrocytes to gradually increased swimming exercise and swimming test to exhaustion in thermoneutral water
Aim of study: Exploration of erythrocytes adaptive reactions in rats performing swimming tests “to exhaustion” in thermoneutral water, with weight of 4% of body mass, after 40-day swimming training at that temperature with gradually increasing loads.
Текст научной статьи Реакция эритроцитов крыс на ступенчатую плавательную нагрузку и тест «до отказа» в термонейтральной среде
Эффекты физических нагрузок универсальны и оказывают воздействие на все процессы в организме, влияя на их направленность [22]. При этом морфологическая модификация эритрона [25] сопровождается увеличением количества эритроцитов, высокорезистентных полихроматофильных, обратимо измененных форм клеток [12] и содержанием гемоглобина в крови [6, 7] в связи с необходимостью экстренного восполнения дефицита кислорода [28, 32]. В течение длительных физических упражнений повышенная пролиферативная активность ретикулоцитов [3] с их метаболической активностью и высокой трансформационной способностью [8, 27] ускоряет процесс обновления красной крови и способствует повышению физической работоспособности организма [2]. Постепенно возрастающая физическая нагрузка обеспечивает торроидальность эритроцитов, то есть их наиболее эффективное движение в сосудистом русле крови [16], при этом существенно разнообразие диаметра эритроцитов и их концентрация в крови в норме не менее 3,5-4,0×106 в 1мм3 [23]. Соответствие количества эритроцита с их диаметром у человека и животных имеет диагностическое значение [13], поскольку функционально изменение величины эритроцитов связано с оксигенацией [9] и их сорбционной способностью [15]. Соотношение этих параметров в условиях оптимального уровня эритроцитарного пула дает возможность адекватной оценки реакции системы крови на физическую нагрузку.
Материал и методы исследований. Исследование проводили на половозрелых самцах крыс Wistar с массой тела 250-300 г. Животных содержали по четыре особи в клетке на стандартном рационе вивария, со свободным доступом к воде, при температуре 21±1 °С и 12-часовом освещении. В контрольную группу были включены животные, содержавшиеся на стандартном рационе в условиях вивария ВивК (n=10) и животные экспериментальных групп Плав4 % (n=8) и Плав4 %Т (n=8). Животные группы Плав4 % после предварительной адаптации к воде [24] плавали с грузом 4 % от массы тела, однократно, «до отказа». Плавание животных группы Плав4 %Т также начинали с их адаптации к условиям эксперимента [29], в ходе которой время нахождения крыс в десатурированной воде с температурой +34-36 °С в непрозрачной емкости глубиной 60 см в течение первых 5 дней постепенно увеличивалось с 2-х до 4х минут без нагрузки. Нагрузочное тестирование животных выполняли по 4 мин ежедневно с 6-го по 40 день [29] с модификациями, заключавшимися в постепенном увеличении массы груза с 0 до 4 % массы тела и последующем выполнении плавательного теста «до отказа». Определение полного утомления во время плавательного теста «до отказа» у животных представлено в работе [19]. В крови, стабилизированной гепарином (5000
ед/мл, АКОС, Россия), определяли уровень гемоглобина по Сали ГС-3, гематокрита с использованием центрифуги MPW-310 («Mechanika Precyzyina», Poland), количество эритроцитов в камере Горяева и ретикулоцитов на мазках после инкубирования крови в течение 12-15 мин с 1% раствором бриллиантового крезилового синего. Измеряли диаметр 100 эритроцитов, окрашенных по Романовскому-Гимза (Vital-Development, Россия) с помощью микроскопа PZO (Poland) с масляной иммерсией, увеличение об. 100х ок. с градуированной шкалой 12х [17], с дальнейшим построением кривой Прайс-Джонса [31]. Значимость различий реакций клеток крови у исследованных групп животных определяли с использованием критерия Крускала-Уоллеса, учитывали средние величины показателей (M) и ошибку средней (m).
Различия считались значимыми при Р≤0,05. Рассчитывали коэффициент ранговой корреляции по Спирмену [5].
Результат исследований. Влияние ступенчатой плавательной нагрузки у животных в группе Плав4 %Т проявилось в отсутствии значимых изменений в эритроцитарном составе крови по сравнению с группой ВивК, за исключением снижения концентрации гемоглобина в эритроците (Р≤0,001). У животных группы Плав4 %Т по сравнению с животными группы Плав4% отмечено двукратное увеличение времени плавания (Р≤0,05), уменьшение диаметра эритроцитов (Р≤0,01), их объема (Р≤0,05), содержания гемоглобина в эритроците (Р≤0,05), относительного (Р≤0,05) и абсолютного количества ретикулоцитов (Р≤0,01) (Таблица 1).
Таблица 1 – Время плавания и клеточный состав красной крови крыс в тесте «до отказа» (M±m)
|
Показатель |
ВивК |
Плав4% |
Плав4%Т |
|
Время плавания, мин |
0 |
91,16 ± 32,15 |
184,75 ± 23,67• |
|
Гематокрит, % |
44,84 ± 0,33 |
49,49 ± 0,81 |
46,58 ± 1,76 |
|
Гемоглобин, г/л |
144,90 ± 1,22 |
148,13 ± 2,24 |
141,12 ± 4,69 |
|
Эритроциты, 1012/л |
8,16 ± 0,16 |
7,29 ± 0,19** |
7,79 ± 0,32 |
|
Диаметр эритроцитов, мкм |
6,18 ± 0,02 |
6,40 ± 0,03*** |
6,21 ± 0,04•• |
|
Площадь поверхности эритроцитов, мкм2 |
74,03 ± 0,66 |
79,45 ± 0,69*** |
78,29 ± 1,97 |
|
Площадь поверхности эритроцитов, м2/л |
601,42 ± 9,85 |
579,10 ± 14,86 |
602,10 ± 23,60 |
|
Объем эритроцитов, мкм3 |
55,08 ± 0,97 |
68,57 ± 2,60*** |
59,41 ± 2,17• |
|
Содержание гемоглобина в 1 эритроците, пг |
17,79 ± 0,03 |
20,42 ± 0,62** |
18,03 ± 0,74• |
|
Концентрация гемоглобина в 1 эритроците, % |
32,31 ± 0,28 |
29,94 ± 0,33** |
30,34 ± 0,29*** |
|
Ретикулоциты, 1012/л |
0,10 ± 0,01 |
0,16 ± 0,02* |
0,09 ± 0,01•• |
|
Ретикулоциты, ‰ |
12,61 ± 0,65 |
19,52 ± 2,93* |
11,59 ± 1,41• |
|
Эритроциты в диапазоне 6,0-6,3 мкм, % |
40,40 ± 2,68 |
26,00 ± 2,51** |
34,31 ± 4,50 |
|
Ретикулоциты 1-2 группы в диапазоне 6,4-6,7 мкм, % |
45,85 ± 1,20 |
44,88 ± 2,26 |
45,00 ± 2,36 |
|
Ретикулоциты 3-4 группы в диапазоне 6,4-6,7 мкм, % |
34,50 ± 6,51 |
39,75 ± 2,66 |
39,63 ± 1,38 |
Примечание: различия значимы * - P≤0,05, ** - P≤0,01, *** - P≤0,001 по сравнению с группой ВивК; • - Р≤0,05, •• - Р≤0,01, •••-Р≤0,001 – по сравнению с группой Плав4%
Распределение эритроцитов по диаметру с максимальным их количеством в диапазоне 6,0-6,3 мкм показало тенденцию к правостороннему сдвигу кривой Прайс-Джонса на 24,2% относительно животных группы Плав4% (Рисунок 1).
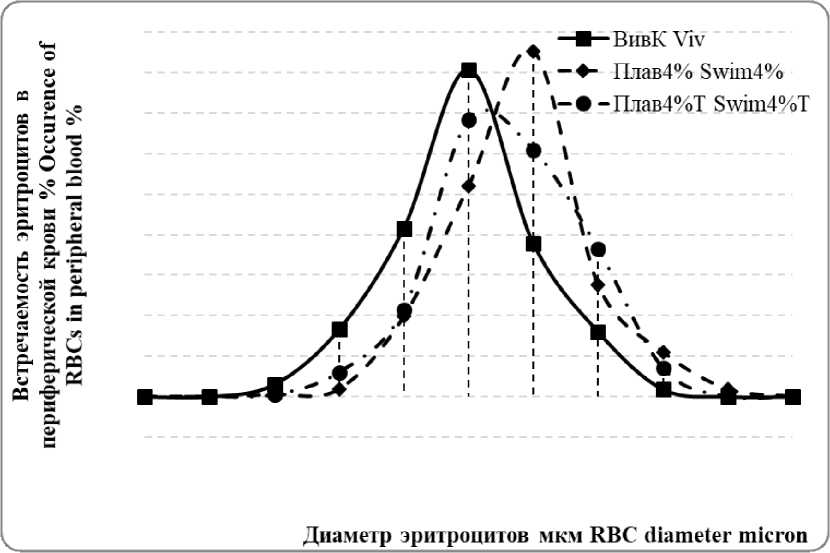
Рисунок 1 – Кривая Прайс-Джонса встречаемости эритроцитов в периферической крови крыс контрольной и экспериментальных групп
Независимо от характера физической нагрузки у молодых эритроцитов с максимальным относительным их количеством в диапазоне 6,4-6,7 мкм, в отличие от зрелых эритроцитов, сдвиг в кривой Прайс-Джонса не выявлен (Рисунок 2).
Отмечено значимое влияние постепенно возрастающей плавательной нагрузки у крыс на взаимосвязь количества эритроцитов разной степени зрелости с их площадью поверхности (Таблица 2).
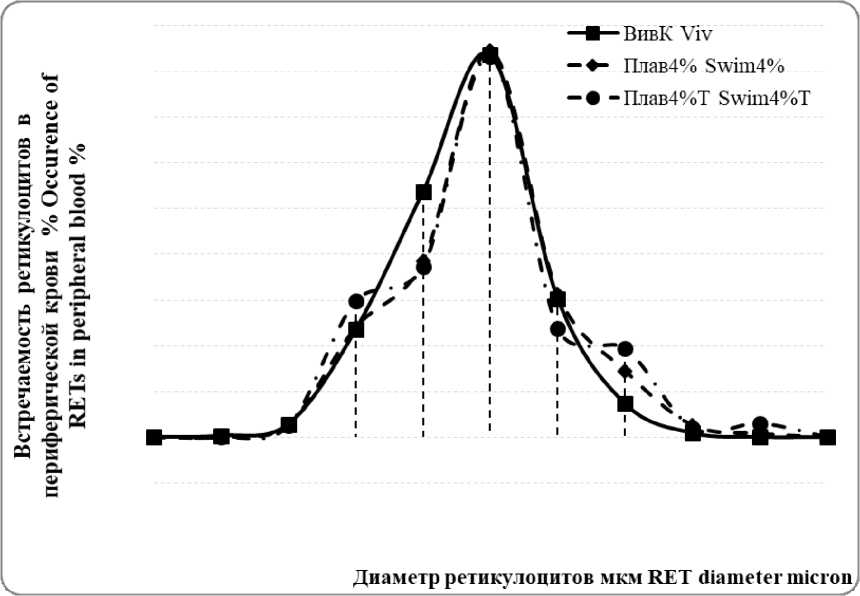
Рисунок 2 – Кривая Прайс-Джонса встречаемости ретикулоцитов в периферической крови крыс контрольной и экспериментальных групп
Таблица 2 – Значение коэффициента корреляции между эритроцитарными показателями у животных исследуемых групп
|
Показатель |
ВивК Viv |
Плав4% Swim4% |
Плав4%Т Swim4%T |
|
Parameters |
Площадь поверхности эритроцитов, мкм2 RBC area surface, µm2 |
||
|
Эритроциты, 1012/л RBC, 1012/l |
-0,224; р=0,514 |
-0,244; р=0,540 |
-0,714; р=0,048 |
|
Эритроциты, мкм3 RBC, µm3 |
0,164; р=0,632 |
0,232; р=0,540 |
-0,262; р=0,501 |
|
Ретикулоциты, 1012/л RET, 1012/l |
Площадь поверхности ретикулоцитов, мкм2 RET area surface, µm2 |
||
|
-0,309; р=0,368 |
0,727; р=0,048 |
-0,899; р=0,004 |
|
В условиях
Ретикулоциты 3-4 группы зрелости, по ступенчатой
плавательной нагрузки двукратное сравнению с ретикулоцитами 1-2 групп увеличение времени плавания животных в группе Плав4%Т по сравнению с группой Плав4%, по-видимому, является проявлением общей закономерности повышения работоспособности организма [11]. Известно, что адекватные физические нагрузки способствуют нивелированию стресс-реакции [19] и поддержанию оптимального уровня оксидативной системы организма животных [1], повышению насыщения крови кислородом, за счет снижением концентрации гемоглобина в эритроците [30]. В свою очередь, уменьшение гипоксического эффекта оказывает влияние на характер трансформации эритроцитов [21]. У животных Плав4%Т тенденция к снижению относительного количества эритроцитов в диапазоне 6,0-6,3 мкм (правосторонний сдвиг кривой Прайс-Джонса) менее выражена чем у животных в группе Плав4% (Таблица 1, Рисунок 1А). Полагаем, что уменьшенный сдвиг кривой Прайс-Джонса при ступенчатой физической нагрузке может быть связан с изменением кислотно-щелочного равновесия циркулирующей крови [31]. Независимо от характера физической нагрузки у молодых эритроцитов с максимальным относительным их количеством в диапазоне 6,4-6,7 мкм (в отличие от зрелых эритроцитов), отсутствие сдвига кривой Прайс-Джонса, может быть связано с их бо́льшей устойчивостью к изменениям внутренней среды и ускоренным созреванием [10].
зрелости, проявляют тенденцию к снижению их относительного количества в данном диапазоне и при этом теряют до 1014% площади мембран в процессе везикуляции [26]. Наличие корреляционной связи между количеством эритроцитов и величиной их площади поверхности (-0,714, р=0,048) у крыс группы Плав4%Т может являться следствием включения механизма регуляции поверхности эритроцитарных клеток в крови, которая, согласно исследованиям А.Л. Чижевского [23], находится в эквивалентной зависимости от поверхности всех капилляров. Известно, что у представителей грызунов отношение количества эритроцитов – 6,9×1012/л к их диаметру – 6,0-7,9 мкм близко к 1,0 [14]. Поэтому у крыс взаимосвязь величин этих параметров может быть менее выражена по сравнению с копытными животными и птицами, имеющими более высокие и низкие коэффициенты – 2,9 и 0,03 соответственно. Разный характер проявления корреляционной связи между количеством ретикулоцитов и величиной их площади поверхности у экспериментальных животных, возможно, обусловлен реактивностью функционально неравнозначных эритроцитов на изменение факторов среды и вариабельностью их диаметра в циркулирующей крови [18, 20]. Отсутствие корреляции между площадью поверхности эритроцитов, обеспечивающей сорбционную функцию [4] и изменением их объема, связанного с диссоциацией гемоглобина [30] в покое или после физической нагрузки, может быть результатом изменения площади поверхности эритроцита в процессе его набухания, прежде всего, среди нормоцитов, активно реагирующих на изменение среды [21]. Предполагаем, что характер набухания эритроцита также может обусловливать появления положительной или отрицательной корреляционной связи между анализируемыми показателями.
Заключение. Полученные результаты отражают морфофункциональное состояние красной крови у животных в зависимости от интенсивности плавательной нагрузки. Установлено, что у крыс постепенное увеличение плавательной нагрузки в термонейтральной среде способствовало незначительному правостороннему сдвигу кривой Прайс-Джонса, снижению концентрации гемоглобина в эритроците и отсутствию ретикулоцитоза. Изменения в эритроцитарном составе могут свидетельствовать об адаптивном ответе, связанном со снижением функциональной напряженности транспортной системы крови при двукратном увеличения времени плавания, по сравнению с идентичной, но однократной нагрузкой в тесте «до отказа». Полагаем, что физическая нагрузка способствует проявлению механизма, регулирующего соответствие количества эритроцитов (в том числе и ретикулоцитов) с величиной их площади поверхности в процессе эритропоэза.
Резюме
Целью работы явилось определение адаптационной реакции клеток красной крови у крыс после 40 дней возрастающей плавательной нагрузки до 4% от массы тела в термонейтральной среде в тесте «до отказа». Показано, что у животных в результате выполнения ступенчатой физической нагрузки двухкратное увеличение предельного времени плавания сопровождалось снижением концентрации гемоглобина в эритроците, отсутствием ретикулоцитоза и незначительным сдвигом распределения эритроцитов кривой Прайс-Джонса в диапазоне 6,0-6,3 мкм. Предполагаем, что наличие корреляции между количеством эритроцитов и их диаметром может свидетельствовать о проявлении механизма, регулирующего это соотношение. Изменения в эритроцитарном составе, связанные со снижением функциональной напряженности транспортной системы крови по сравнению с идентичной, но однократной нагрузкой в тесте «до отказа», могут являться маркерами физиологически обоснованного адаптивного ответа на ступенчатую физическую нагрузку.
Список литературы Реакция эритроцитов крыс на ступенчатую плавательную нагрузку и тест «до отказа» в термонейтральной среде
- Балыкин, М. В. Изменение газового состава крови и процессы свободнорадикального окисления липидов в миокарде при адаптации к физическим нагрузкам / М. В. Балыкин, С. А. Сагидова, А. В. Жарков // Рос. физиол. журн. им. И. М. Сеченова. – 2015. – Т. 101, № 9. – С. 1007-1012.
- Викулов, А. Д. Диаметр эритроцитов как надежный маркер текущего функционального состояния организма и физической работоспособности спортсменов / А. Д. Викулов, В. А. Маргазин, В. Л. Бойков // Лечебная физкультура и спортивная медицина. – 2015. – № 1. – С. 10-14.
- Вычужанова, Е. А. Влияние хронического стресса на острую стресс-реакцию у крыс / Е. А. Вычужанова // Наука и образование: проблемы, идеи, инновации. – 2015. – № 1. – С. 9-11.
- Гареев, Р. А. Некоторые итоги исследований и перспективы изучения адсорбционно-транспортной функции эритроцитов / Р. А. Гареев // Вестник КазНУ. Серия биологическая. – 2010. – Т. 2. – № 44. – С. 103-109.
- Гланц, С. Медико- биологическая статистика / С. Гланц. – Практика, 1999. – 459 с.
- Дроздов, Д. Н. Особенности вариаций показателей периферической крови до и после действия физической нагрузки / Д. Н. Дроздов, А. В. Кравцов // Специфические и неспецифические механизмы адаптации во время стресса и физической нагрузки: материалы II Республиканской научно-практической Интернет-конференции с международным участием. – Гомель, 2014. – С. 99-101.
- Жлобович, И. Н. Функциональное значение особенностей периферической крови у спортсменов с различной направленностью тренировочного процесса / И. Н. Жлобович // Научные труды НИИ физической культуры и спорта Республики Беларусь. – 2014. – № 14. – С. 246-250.
- Зинчук, В. В. Деформируемость эритроцитов: физиологические аспекты / В. В. Зинчук // Успехи физиологических наук. – 2001. – Т. 32. – № 3. – С. 66-78.
- Иванов, К. П. Основы энергетики организма. Т.2. Биологическое окисление и его обеспечение кислородом / К. П. Иванов. – СПб.: Наука, 1993. – 270 с.
- Иржак, Л. И. Дыхательная функция крови в условиях гипероксии / Л. И. Иржак, В. В. Гладилов, Н. А. Мойсеенко. – Медицина, 1985. – 176 с.
- Влияние прерывистой нормобарической гипоксии на системную гемодинамику, биохимический состав крови и физическую работоспособность лиц пожилого возраста / Е. А. Ключникова, Л. В. Аббазова, М. А. Лоханникова, С. С. Ананьев, Д. А. Павлов, М. В. Балыкин // Ульяновский медико-биологический журнал. – 2017. – № 4. – С. 155.
- Клетки крови и современные технологии их анализа / Г. И. Козинец, В. М. Погорелов, Д. А. Шмаров [и др.]. – Москва: Триада-фарм, 2002. – 534 с.
- К вопросу о соотношении количества эритроцитов и их размеров в периферической крови / Г. И. Козинец, Д. А. Шмаров, Т. Н. Левина [и др.] // Клиническая лабораторная диагностика. – 2002. – № 4. – С. 43-45.
- Коржуев П.А. Гемоглобин / П. А. Коржуев. – М.: Наука, 1964. – 286 с.
- Адсорбция углеводов и липидов эритроцитами при недозированной физической нагрузке / С. Г. Макарушко, А. А. Мурзамадиева, З. Ш. Смагулова [и др.] // Рос. физиол. журн. им. И.М. Сеченова. – 2004. – Т. 90, № 8. – С. 140-141.
- Две стратегии реологии эритроцитов крыс Vistar при физической нагрузке / М. Н. Маслова, А. М. Казеннов, Л. Н. Катюхин [и др.] // Рос. физиол. журн. им. И.М.Сеченова. – 2004. – Т. 90. – № 8. – С. 155-156.
- Медведев, В. В. Клиническая лабораторная диагностика. Справочник / В. В. Медведев, Ю. З. Волчек. – СПб.: Гиппократ, 2006. – 360 с.
- Физиологическое распределение эритроцитов на уровне дуги аорты по данным цитометрического и спектрофлуометрического исследования / М. А. Медведев, Г. С. Коваль, Н. В. Рязанцева [и др.] // Вестн. Том. гос. ун-та. – 2007. – Т. 300. – № 2. – С. 170-171.
- Реактивность эритроцитов крыс в условиях физической нагрузки разной интенсивности / Н. П. Монгалёв, Л. Ю. Рубцова, В. Д. Шадрина [и др.] // В мире научных открытий. – 2018. – Т. 10. – № 2. – С. 74-92.
- Монгалёв, Н. П. Вариабельность диаметра эритроцитов взрослого человека в условиях 80-минутного мониторинга / Н. П. Монгалёв, Л. И Иржак // Вестник Сыктывкарского университета. Серия 2 (биология, геология, химия, экология). – 2015. – № 5. – С. 63-67.
- Монгалёв, Н. П. Реактивность нормоцитов красной крови человека в условиях физической нагрузки и острой нормобарической гипоксии / Н. П. Монгалев, Л. Ю. Рубцова, Н. Н. Потолицына. – В кн.: Бойко Е.Р., ред. Физиолого-биохимические механизмыобеспечения спортивной деятельности зимних циклических видов спорта. – Сыктывкар: ООО «Коми республиканская типография», 2019. – 256 с.
- Хныченко, Л. К. Стресс и его роль в развитии патологических процессов / Л. К. Хныченко, Н. С. Сапронов // Обзоры по клинич. фармакол. и лек. терапии. – 2003. – Т. 2. – № 3. – С. 2-15.
- Чижевский, А. Л. Структурный анализ движущейся крови / А. Л. Чижевский. – М.; 1989. – 266 с.
- Intensity of swimming exercise influences tracheal reactivity in rats / A. F. Brito, A. S. Silva, I. L. L. Souza [et al.] // J Smooth Muscle Res. – 2015. – № 51. – Р. 70-81.
- Changes of RBC aggregation in oxygenation-deoxygenation: pH dependency and cell morphology / I. Cicha, Y. Suzuki, N. Tateishi [et al.] // Am. J. Physiol. Heart Circ. Physiol. – 2003. – T. 284. – № 6. – P. 2335-2342.
- A detailed study of time dependent changes in human red blood cells: from reticulocyte maturation to erythrocyte senescence / S. C. Gifford, J. Derganc, S. S. Shevkoplyas [et al.] // British Journal of Haematology. – 2006. – T. 135. – № 3. – Р. 395-404.
- Higher levels of erythrocyte membrane fluidity in sprinters and longdistance runners / T. Kamada, S. Tokuda, S. Aozaki [et al.] // J. Appl. Physiol. – 1993. – № 74. – Р. 354-358.
- Peripheral Circulation / M. Laughlin, M. Davis, N. Secher [et al.] // Compr Physiol. – 2012. – Т. 2. – № 1. – Р. 321-447.
- The effects of swimming training in cold water on antioxidant enzyme activity and lipid peroxidation in erythrocytes of male and female aged rats / A. Lubkowska, I. Bryczkowska, I. Gutowska [et al.] // Int. J. Environ. Res. Public Health. – 2019. – Т. 16, № 4. – Р. 1-15.
- Mairbäurl, H. Red blood cells in sports: effects of exercise and training on oxygen supply by red blood cells / H. Mairbäurl // Frontiers in Physiology. – 2013. – № 4. – Р. 332.
- Price – Jons, C. Red Blood Cell Diameters / C. Price – Jons. – L.: Oxford University, 1933. – 82 р.
- Veranderungen erythrozytar kriterien des meerschweinchens bei hypoxie / G. Schmidt, J. Gross, E. Necas [et al.] // Acta boil. Med. Germ. – 1977. – № 36. – Р. 205-211.
