Reconstructed paleodiets and subsistence strategies of the Central Ciscaucasian population (1000 BC to 1000 AD), based on collagen isotope analysis of bone samples from the Kichmalka II burial ground
, based on collagen isotope analysis of bone samples from the Kichmalka II burial ground")
Author: Babenko A.N., Dobrovolskaya M.V., Korobov D.S., Vasilyeva E.E.
Journal: Archaeology, Ethnology & Anthropology of Eurasia @journal-aeae-en
Section: The metal ages and medieval period
Article in issue: 4 т.49, 2021.
Free access
Short address: https://sciup.org/145146412
IDR: 145146412 | DOI: 10.17746/1563-0110.2021.49.4.080-090
Text of the article Reconstructed paleodiets and subsistence strategies of the Central Ciscaucasian population (1000 BC to 1000 AD), based on collagen isotope analysis of bone samples from the Kichmalka II burial ground
The isotopic composition of the biological remains of people, animals, and plants captures the circumstances of their lives, and this is why analysis of these parameters has become so widespread in archaeological science. This study intended to clarify the traditions of nutrition and subsistence strategies among the population living in the foothills of the North Caucasus in the 1st millennium BC to the 1st millennium AD. The
data on the isotopic composition of carbon and nitrogen in collagen of humans and animals bone tissue from the burials in the Kichmalka II cemetery, as well as data obtained from the study of plant materials from the dung of modern small ruminants, were used. As is known, the reconstructions of human and animal diets are based on the general regularity of the isotopic composition (nitrogen and carbon) dynamics (fractionation) in trophic chains of ecosystems: it naturally changes while moving from one level to another (Ambrose, 1993). The reliability of the reconstruction depends on the validity of our knowledge about the size of the trophic step (Dobrovolskaya et al., 2020). It is also important to mention that both general isotopic features of ecosystems and sizes of trophic steps within these systems cannot be expressed in uniform values. Therefore, each study of isotopic composition of biological evidence from archaeological sites combines general features of an ancient ecosystem with individual bioarchaeological reconstructions.
Since ancient times, the population that lived in the foothills of the North Caucasus practiced diversified agriculture, combining animal husbandry and land cultivation (Kaloev, 1981: 7–32). However, information about agricultural activities in individual communities is quite general and often lacks specific content. For example, for a long time, there was an idea that the main occupation of the carriers of the Koban culture was livestock-breeding, while agriculture played an auxiliary role (Kozenkova, 1989: 65; Markovin, Munchaev, 2003: 166–168). The economic activities of the Alanian population in the late period of its history in the North Caucasus are known in better detail. The main activity of the Alans during this period was previously considered to be agriculture combined with transhumance (Kuznetsov, 1971: 47–147).
In recent years, evidence of land cultivation and animal husbandry by the population of the Kislovodsk Basin over a long period—from the early 1st millennium BC to late 1st millennium AD—has been studied in better detail. Soil and archaeological studies of agricultural lands of various forms (terraces, arable land plots, areas with boundary walls), as well as research on stone enclosures for livestock, which have survived in the mountains, have made it possible to clarify the practices of intense land cultivation among the Koban people (Borisov, Korobov, 2013: 182–195), as well as complexity of economy among the Alanian tribes, where in addition to agriculture, transhumance played an important role (Korobov, 2017: 199–227; Korobov et al., 2018). These studies were supported by the analysis of osteological evidence from the early medieval settlements of the 1st millennium AD and macro-remains of cultivated cereals from settlements and agricultural lands in their vicinity (Korobov, 2017: 202–204, 214–215; Sergeev, 2021).
Studying subsistence strategies and nutrition of populations living in different historical periods in the region, using the methods of isotope biogeochemistry, is a special research field. An article by T. Higham and his colleagues (2010) elucidated the reasons for a significant reservoir effect discovered in dating bone remains from the Klin-Yar III cemetery, in the Kislovodsk Basin. As a research tool, the authors used the values of trophic steps in ecosystems, applying the data on archaeological sites and modern materials. The same year, the results of analyzing isotopic composition of carbon and nitrogen in the collagen of human bone tissue from the Chalcolithic and Bronze Age burials in the North Caucasus were published (Hollund et al., 2010). That article discussed the role of the local environment that could significantly affect the specific features of collagen in local inhabitants (both humans and animals). The results of large-scale isotope studies were published by the international team that studied the diets and lifestyles of some population groups in various periods of the Bronze Age in the North Caucasus (Knipper et al., 2018, 2020). Among other things, this comprehensive study confirmed a significant influence from local ecosystems on the values of both isotopic ratios and trophic steps in these systems.
Our hypothesis is that the traditions of the carriers of the Koban culture, groups of the Sarmatian period, and Alans were also manifested in the features of sophisticated non-specialized economy. This can be established from the data on the isotopic composition of bone tissue collagen in humans and animals from burials.
Materials
This study is based on the material evidence from the Kichmalka II burial ground explored in 2006–2010 by the expedition from the State Hermitage Museum, under the leadership of E.E. Vasilyeva (Vasilyeva, 2009, 2010, 2012; Vasilyeva, Akhmedov, 2015). Bone samples were taken from burials of the Koban culture (7th to early 5th centuries BC), Sarmatian period (1st to 2nd centuries AD), and the Early Middle Ages (late 5th to early 6th centuries AD). The cemetery is located in the foothills of the Central North Caucasus (Zolsky District of the Kabardino-Balkarian Republic), at a height somewhat exceeding 1000 m (Fig. 1).
The analysis involved thirty two samples: eight belonged to skeletal remains of males (an individual from burial 24 was represented by two samples—spongy and compact bone tissue, which made it possible to assess the stability of isotopic composition of collagen in tissues with different rates of bone remodeling); ten samples belonged to females (an individual from burial 28 was also represented by two samples); three samples belonged
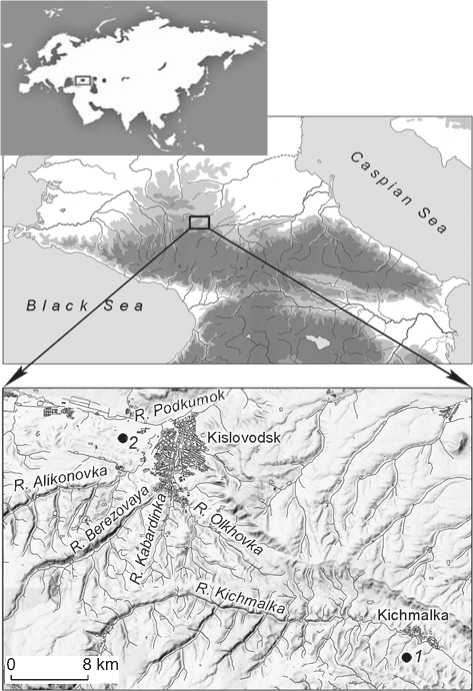
Fig. 1. Location of the Kichmalka II ( 1 ) and Klin-Yar III ( 2 ) cemeteries in the North Caucasus.
to children 5–7 years of age and ca 12 years of age; five samples were taken from sheep/goat bones, four from horse bones, and two from small burrowing animals not associated with ancient times (Tables 1, 2). In addition, data on four modern samples of sheep dung collected at various heights was obtained—on the Bermamyt Plateau (Karachay-Cherkessia Republic) and in the vicinity of Kislovodsk (Table 3). For analyzes, average samples were taken from five to seven pellets.
Methods
Samples were prepared in the Laboratory of Contextual Anthropology at the Institute of Archaeology of the Russian Academy of Sciences. Fragments of bone tissue were washed in distilled water and brought to constant weight. Dry fragments were placed in 1M hydrochloric acid in a ratio of at least 1 g per 50 ml, and were left at a temperature of +3 °C until complete demineralization. Then, the samples were repeatedly washed with distilled water until they reached pH = 7. At the next stage, the samples were placed in 0.1M alkali solution for 24 hours, after which they were washed again. Then, the samples were placed in a solution of hydrochloric acid with pH = 2.5 (10 ml) and transferred to a thermostat where collagen dissolved at a temperature of +65 °C. The liquid was centrifuged for collagen purification. The solution was evaporated in a thermostat at +40 °C. The sample weight for the analysis was ca 400 μg. Collagen preservation was assessed by the C/N ratio (2.9–3.6) (DeNiro, 1985) and by the content of carbon (from 30 to 47 %) and nitrogen (from 11.0 to 17.3 %) in a sample (Ambrose, 1993; Van Klinken, 1999).
Samples of modern manure were treated with hot 5 % HCl solution and washed through a sieve (0.25 mm) for removing mineral grains and large plant residues, in order to average the material as best as possible. Then, the samples were repeatedly washed with distilled water and dried. Thus, the results obtained reflected the content of stable isotopes of carbon and nitrogen directly in the averaged sample of plant residues. The analyzed sample weight was ca 1500 μg.
The isotopic composition of the samples was established using a mass spectrometer (Thermo Finnigan Delta V Plus, Center for Collective Use at the Institute of Ecology and Evolution of the Russian Academy of Sciences, Moscow). For statistical comparison of the samples, the Mann-Whitney U -test was used, since the distribution of the parameters was abnormal.
For establishing the contribution of plant and animal foods to human diet, one needs to assess the difference between the isotopic data of the person and the cultivated plants and animals that he could have eaten. Trophic step is the difference between the isotopic signatures (∆δ13C and ∆δ15N) of producers (plants)/ the first order consumers (herbivores), and the first order (herbivores) and the second order (predators) consumers. The δ13C and δ15N values in bone collagen increase with each trophic level by 0–2 ‰ (Bocherens, Drucker, 2003) and 3–5 ‰ (even up to 6 ‰, in humans) (Ambrose, 1993; O’Connell et al., 2012), respectively. One may estimate the contribution of a specific component to the human diet from how much ∆δ Chuman-animal and ∆δ Nhuman-animal differ from the value of the trophic step. Low values of the indicators most often point to an insignificant share of meat and dairy products, consumed by humans. Values exceeding the trophic step may indicate other food sources (Ambrose, 1993; O’Brien, 2015).
The lack of isotopic data on the plant food of ancient and medieval humans in the region under consideration limits our opportunities for reconstructing their diet. Therefore, we can only proceed from the generally accepted assumptions about the isotopic signals of plants belonging to the C3- and C4-type of photosynthesis. The δ13C value of –18 ‰ marks the beginning of consuming C4-type plants (Pearson et al., 2007).
Table 1. Individual values of δ13С and δ15N in the samples of bone collagen taken from the persons buried at the Kichmalka II burial ground
|
Year of excavations |
Number of burial/ skeleton |
Sex |
Age |
δ 13 C |
δ 15 N |
%C |
%N |
C/N atom. |
|
Koban culture |
||||||||
|
2007 |
3 |
Male |
Over 40 |
–16.84 |
10.28 |
43.3 |
15.2 |
3.3 |
|
2008 |
8 |
ʺ |
30–39 |
–17.66 |
8.78 |
41.9 |
15.4 |
3.2 |
|
2008 |
10 |
Female |
20–25 |
–18.26 |
10.07 |
41.9 |
15.3 |
3.2 |
|
2009 |
20 |
Male |
Over 50 |
–18.16 |
10.23 |
42.6 |
15.7 |
3.2 |
|
2009 |
21 |
Female (?) |
10–12 |
–18.66 |
10.11 |
44.7 |
15.9 |
3.3 |
|
2009 |
24 (а) |
Male |
Over 45 |
–18.61 |
9.62 |
43.3 |
15.0 |
3.4 |
|
2009 |
24 (b) |
ʺ |
Over 45 |
–18.78 |
9.77 |
43.5 |
14.9 |
3.4 |
|
2009 |
25 |
Female |
Around 20 |
–16.68 |
9.42 |
41.8 |
15.1 |
3.2 |
|
2009 |
26 |
ʺ |
Over 50 |
–17.91 |
10.43 |
44.7 |
15.7 |
3.3 |
|
2009 |
23 |
? |
5–7 |
–16.97 |
9.39 |
36.3 |
12.3 |
3.4 |
|
2010 |
30 |
Male (?) |
Around 12 |
–17.32 |
9.85 |
41.4 |
14.2 |
3.4 |
|
Sarmatian period |
||||||||
|
2010 |
29/1 |
Male |
Over 50 |
–17.92 |
9.32 |
42.0 |
15.2 |
3.2 |
|
2010 |
29/2 |
Female |
Over 60 |
–18.07 |
8.72 |
42.2 |
15.3 |
3.2 |
|
2009 |
22 |
ʺ |
Around 20 |
–17.91 |
9.26 |
42.4 |
15.1 |
3.3 |
|
2010 |
31 |
ʺ |
30–49 |
–16.03 |
10.88 |
43.7 |
15.5 |
3.3 |
|
Alanian culture |
||||||||
|
2010 |
28 (а) |
ʺ |
40–49 |
–18.16 |
9.45 |
38.7 |
13.9 |
3.3 |
|
2010 |
28 (b) |
ʺ |
40–49 |
–17.73 |
9.42 |
44.0 |
15.3 |
3.3 |
|
2010 |
33/1 |
Male |
17–20 |
–18.30 |
9.15 |
42.4 |
15.7 |
3.2 |
|
2010 |
33/2 |
Female |
16–18 |
–18.86 |
9.99 |
42.6 |
15.6 |
3.2 |
|
2010 |
34/1 |
Male |
20–25 |
–18.04 |
11.18 |
43.6 |
15.8 |
3.2 |
|
2010 |
34/2 |
Female |
30–39 |
–18.10 |
9.63 |
42.6 |
15.2 |
3.3 |
While making comparison with modern isotope data, one needs to keep in mind the changes in the isotopic composition of the atmosphere, which has become enriched by ancient carbon with low δ13C values in the industrial and postindustrial periods (the Suess effect). The δ13C value of atmospheric CO2 recovered from the bubbles in ice-core decreased from –6 ‰ ca 11 ka BP (Indermühle et al., 1999) to ca –8 ‰ today (White, Vaughn, Michel, 2015). This 2 ‰ shift must be taken into account when interpreting the carbon isotopic composition of archaeological samples.
Results
Carbon. The individual variability of carbon isotope ratios in human bone collagen varied from –18.86 to –16.03 ‰ (see Table 1). The δ13C values for the buried persons of the Koban and Sarmatian periods varied within a fairly wide ranges from –18.78 to –16.68 ‰ and from –18.07 to –16.03 ‰, respectively. The carriers of the Alanian culture showed a slight variability of this indicator (from –18.86 to –17.73 ‰).
The δ13C values in herbivorous domestic animals varied from –20.84 to –19.77 ‰ (see Table 2). Horse
Table 2. Individual values of δ13С and δ15N in the samples of bone collagen taken from animals buried at the Kichmalka II burial ground
|
Year of excavations |
Place of discovery |
Animal |
δ 13 C |
δ 15 N |
%C |
%N |
C/N atom. |
Cultural attribution of the burial |
|
2010 |
Burial 34 |
Burrowing animal |
–23.80 |
5.36 |
47.7 |
12.9 |
4.3 |
Alanian |
|
2010 |
Burial 34, destr. part |
Small ruminants |
–19.95 |
3.73 |
40.1 |
14.4 |
3.3 |
ʺ |
|
2010 |
Burial 33 |
ʺ |
–19.91 |
3.80 |
41.7 |
15.1 |
3.2 |
ʺ |
|
2010 |
Next to burial 31 |
Horse |
–20.34 |
5.52 |
46.2 |
15.1 |
3.6 |
Sarmatian |
|
2010 |
Burial 29 |
Small ruminants |
–19.77 |
4.75 |
42.4 |
15.3 |
3.2 |
ʺ |
|
2009 |
Next to burial 24 |
Horse |
–20.46 |
4.67 |
41.9 |
14.4 |
3.4 |
Koban |
|
2009 |
ʺ |
ʺ |
–20.84 |
4.90 |
42.9 |
15.2 |
3.3 |
ʺ |
|
2009 |
Burial 24 |
Small ruminants |
–20.37 |
5.57 |
41.2 |
15.0 |
3.2 |
ʺ |
|
2007 |
Burial 3 |
Horse |
–20.68 |
5.22 |
41.2 |
14.8 |
3.2 |
ʺ |
|
2009 |
Burial 24 |
Burrowing animal |
–18.73 |
7.09 |
39.5 |
13.3 |
3.5 |
ʺ |
|
2010 |
Burial 32 |
Small ruminants |
–20.01 |
5.79 |
32.1 |
11.2 |
3.4 |
ʺ |
Table 3. Values of δ13С и δ15N in the samples of modern manure of small ruminants
|
Height above sea level, m |
δ 13 C |
δ 15 N |
%C |
%N |
C/N atom. |
|
2048 |
–28.68 |
2.54 |
43.29 |
2.36 |
21.40 |
|
2123 |
–28.35 |
2.45 |
43.10 |
1.89 |
26.61 |
|
2217 |
–28.66 |
2.69 |
42.87 |
2.33 |
21.48 |
|
940 |
–27.91 |
4.44 |
42.81 |
2.37 |
21.05 |
and sheep/goat collagen demonstrated the uniformity of carbon isotopic composition (from –20.84 to –20.34 ‰ and from –20.37 to –19.77 ‰, respectively).
Nitrogen. The δ15N values in human collagen varied within 8.72–11.18 ‰. Individual variability of this indicator was the smallest among the Koban people (8.78– 10.43 ‰). The ranges for the Alans (9.15–11.18 ‰) and people of the Sarmatian period (8.72–10.88 ‰) were wider (see Table 1). The groups were not statistically different.
The isotopic ratios of nitrogen in collagen of herbivorous domestic animals varied from 3.73 to 5.79 ‰. The range of δ15N values in horses was 4.67– 5.52 ‰. For small ruminants of the Koban period, this indicator was 5.57–5.79 ‰. The content of the 15N isotope in the collagen of sheep/goat bones of the Alanian period was minimal and did not exceed 4 ‰ (see Table 2).
Individual variability of isotopic parameters was not associated with sex and age. No significant differences have been found.
The δ13C value of plant residues from modern dung varied from –27.91 to –28.68 ‰, and δ15N from 2.45 to 4.44 ‰ (see Table 3).
Discussion
Wide variation of δ13C and δ15N in the collagen of bone tissue of the Koban people testified to individual diversity of food consumption. The δ13C variability was 2.1 ‰. Two data clouds can be distinguished (Fig. 2): group 1 (burials 3, 8, 23, 25, and 30) significantly differed from group 2 (burials 10, 20, 21, 24, and 26) in high δ13C values (> –17.7 ‰, p = 0.006). They were variable in δ15N, but they did not differ statistically ( p > 0.05). Accordingly, the nutrition of each group was based on different plant components. A conventional boundary around 18 ‰ of δ13C can be drawn between the groups. This makes it possible to consider consumption of C4-plants as a basis of these differences.
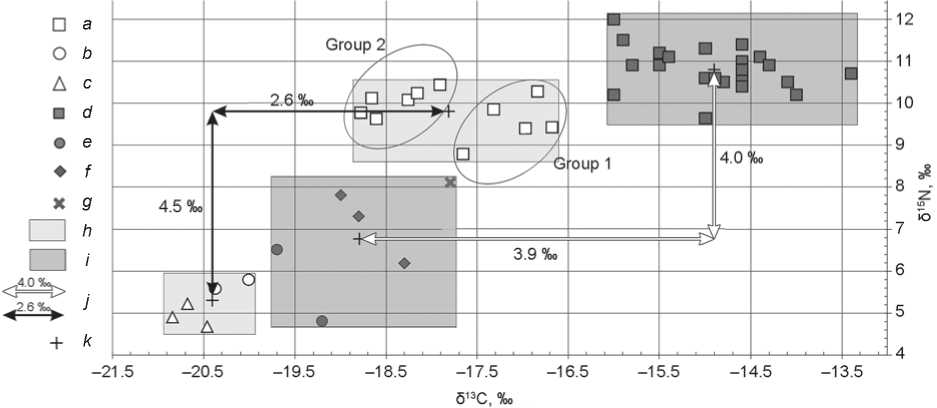
Fig. 2 . Individual δ13C and δ15N values for the samples of bone collagen from humans and animals of the Koban period at Kichmalka II and Klin-Yar III.
a–c – Kichmalka II: a – human, b – small ruminants, c – horse; d–g – Klin-Yar III: d – human, e – small ruminants, f – cattle, g – pig; h , i – area of isotope signals of bone collagen from Kichmalka II and Klin-Yar III, respectively; j – trophic step; k – average value.
The difference between the average δ13C values of individuals from group 1 and herbivores was over 2 ‰, and ∆δ15Nhuman-animal was less than 5 ‰. Thus, the former difference was larger than the trophic step, and the latter difference was within the trophic step (Fig. 2). For some individuals (group 2, ∆δ13C human-animal and ∆δ15N human-animal ), we can reconstruct a diet based on C3-plants and meat products in sufficient quantities. As opposed to the second group, the composition of food among the first group could have included more foods of plant origin with high content of 13C isotope. According to archaeobotanical data, millet ( Panicum miliaceum ) was a cultivated C4-plant in the Koban period in the Kislovodsk Basin, in the immediate vicinity of which the Kichmalka II site is located (Reinhold, Korobov, Belinskij, 2017: 242; Sergeev, 2021: Tab. 8). It can be assumed that there were two food models, and not a single model, in the same culture.
There was a significant similarity between the groups of the Sarmatian and Alanian periods ( p > 0.05): among the main part of the people, values δ13C and δ15N varied in less than 1 ‰. One sample among the human samples of the Sarmatian period was distinguished by higher values of δ13C and δ15N. Unfortunately, only two isotope signals of domestic animals (sheep/goat and horse) of that period are available. As in the case of the Koban people, ∆δ Chuman-animal was over 2 ‰, and ∆δ Nhuman-animal was less than 5 ‰ (Fig. 3). The data obtained indicate that people of the Sarmatian period consumed mainly C3-plants and a sufficient amount of meat and/or milk from animals that fed on this type of plant. The diet of some individuals could have contained a certain share of C4-plants (burial 31).
As opposed to the two periods mentioned above, the difference between the average δ13C values in bone collagen of humans and animals of the Alanian period did not exceed the trophic step (Fig. 4). The δ13C values of almost all the samples (with the exception of one, from burial 28 (b)) indicated a diet based on C3-plants. The data obtained do not contradict the results of archaeobotanical studies in the Alanian settlements of the 5th–8th centuries AD located in the areas of the Kislovodsk Basin adjacent to the Kichmalka II site. Wheat dominated in the regional archaeobotanical complex of the time (65 %), while the share of millet was only 9 % (Sergeev, 2021). One may conclude the dominance of C3-plants in the diet of the Alanian population of the 5th–8th centuries AD.
Another distinctive feature of the early medieval samples was greater difference between the average δ15N values of humans and animals, reaching 6 ‰, and staying within the trophic step (O’Connell et al., 2012). Notably, sheep/goat bones from the Alanian burials showed the lowest nitrogen ratio (less than 4 ‰). Unfortunately, we do not have enough isotopic data for domestic animals. However, it should be taken into account that settlement material evidence from the Kislovodsk Basin reveals the presence of bones not only of small ruminants, but also of cattle, horses, and pigs as food remains in numerous surveyed settlements of the 5th–8th centuries (Korobov, 2017: 202–203). Therefore, it is safe to say that the 6 ‰ difference was caused by the lack of isotopic data. Thus, the Alanian group turned out to be the most compact in terms of the variability of food sources.
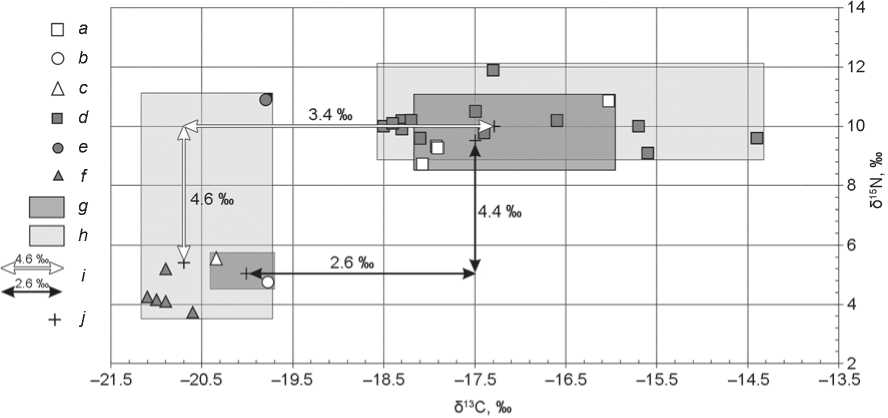
Fig. 3 . Individual δ13C and δ15N values for the samples of bone collagen from humans and animals of the Sarmatian period from Kichmalka II and Klin-Yar III.
a–c – Kichmalka II: a – human, b – small ruminants, c – horse; d–f – Klin-Yar III: d – human, e – small ruminants, f – horse; g , h – area of isotope signals of bone collagen from Kichmalka II and Klin-Yar III, respectively; i – trophic step; j – average value.
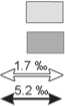
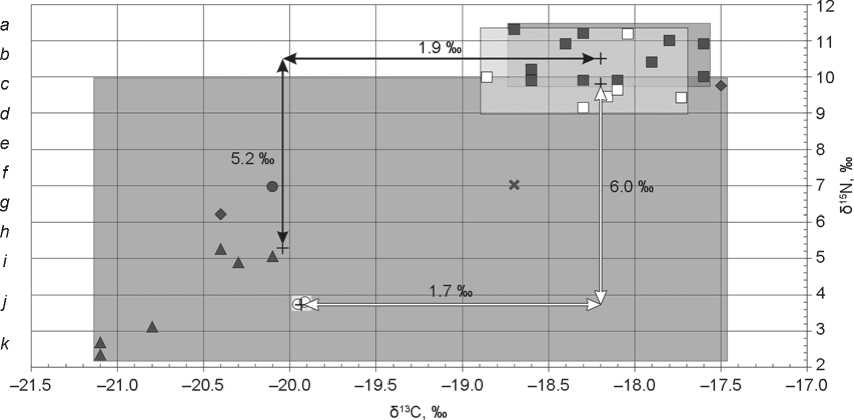
Fig. 4 . Individual δ13C and δ15N values for the samples of bone collagen from humans and animals of the Alanian period from Kichmalka II and Klin-Yar III.
a , b – Kichmalka II: a – human, b – small ruminants; c – g – Klin-Yar III: c – human, d – small ruminants, e – horse, f – cattle, g – pig; h , i – area of isotopic signals of bone collagen from Kichmalka II and Klin-Yar III, respectively; j – trophic step; k – average value.
Research into the isotopic composition of collagen in human and animal bones at the sites of the North Caucasus is at the early stage, which is limited by accumulation of data and attempts to interpret them. This is not an easy task in the conditions of diverse landscapes. For reconstructing the human diet, one needs to take into account a large set of isotopic signals from various sources. In this regard, it is important to compare our results with those published earlier. Comparative evidence was the data on isotopic composition of bone collagen in humans and animals from the Klin-Yar III burial ground in the Kislovodsk Basin (Higham et al., 2010). This site is located at an altitude of 800 m above sea level, at a distance of about 25 km from Kichmalka II (see Fig. 1). Burials of three cultural and chronological periods were also presented at Klin-Yar III. However, unlike Kichmalka II, osteological evidence related to the Koban culture was dated to the earlier time—the 10th–7th centuries BC (Belinskij, Dudarev, 2015: 386–397).
The arrangement of individual data on the diagram of δ13C/δ15N ratio makes it possible to assess the features of each of the cultural and chronological groups from two burial sites (see Fig. 2–4). The individuals from the Koban burials at Kichmalka II and Klin-Yar III significantly differed from each other both in δ15N ( p = 0.000028) and δ13C ( p = 0.000003) values. The ranges of individual δ13C values of these two groups did not even overlap (see Fig. 2). People of the Sarmatian period buried at the Kichmalka II and Klin-Yar III cemeteries showed approximately the same values ( p > 0.05) and wide range of variability of the carbon isotopic composition (from –18.4 to –15.7 ‰). The isotopic values of Alans from the burials at both sites were similar in carbon composition ( p > 0.05). However, the δ15N values for the individuals from Klin-Yar III were statistically higher ( p = 0.035).
The differences in isotopic parameters of humans and animals of the Koban period from the two burial grounds were quite clear, which might had resulted from different landscape and climatic conditions in the vicinity of the sites (Marshall, Brooks, Lajtha, 2007) and/or difference in nutrition (Ambrose, 1993). However, Kichmalka II and Klin-Yar III are located close to each other, in similar landscapes; therefore, the differences in isotopic composition were not associated with habitation conditions. This conclusion is also supported by the fact that such differences were not observed in other groups. There are no reliable climatic reconstructions for the area under discussion in the 10th– 5th centuries BC. However, the synchronicity of the dynamics of glaciers in the North Caucasus and the Alps in the last millennium (Solomina et al., 2016) makes it possible to consider climatic changes in the region using international research. According to the studies of glaciers and lake levels in the Alps (Holzhauser, Magny, Zumbuühl, 2005; Ivy-Ochs et al., 2009; Wirth et al., 2013), the second half of the 1st millennium BC was distinguished by cold and humid conditions. However, the degree of humidity in the north and the south of the Alps was different, owing to the shift of the position of the westerly storm tracks (Westerlies) to the south at that time. It cannot be stated definitively, but can be assumed that in the end of the period under consideration (the time of the burial of the Koban persons at the Kichmalka II burial ground), the amount of precipitation in the region could have increased, as also in the south of the Alps (Wirth et al., 2013). With the increasing moisture content, δ13C and δ15N in C3-plants decreased (Marshall, Brooks, Lajtha, 2007). Therefore, the isotopic signals of bone collagen of humans and animals of the Koban period from Kichmalka II should show lower values of these indicators, which can be observed in Fig. 2.
An indirect argument in favor of this assumption is the termination of the existence of Koban settlements in the Kislovodsk Basin in the mid 1st millennium BC owing to catastrophic changes in agricultural landscapes resulting from climate humidification (Borisov, Korobov, 2013: 191–195).
Individual variability within both groups of the Koban people (from Kichmalka II and Klin-Yar III) in terms of δ13C was great, which suggests different plant food resources. The difference between the average δ13C values of humans and domestic animals from Klin-Yar III was two times greater than the trophic step. The published data on the content of carbon isotopes in the collagen of human bones from other Bronze Age sites in the North Caucasus (Knipper et al., 2020: Tab. 3) show that the average value of δ13C in the Koban people from Kichmalka II (–17.8 ‰) was close to the data from the literature (–18.4 to –19.4 ‰), while those from Klin-Yar III were much higher (–14.3 ‰). The latter indicates a higher proportion of millet in the human diet. Thus, the differences in isotopic signatures of individuals from the Koban burials at both sites are associated with different climatic conditions and food preferences among some groups of people.
The average δ13C value in those buried at the Klin-Yar III cemetery in the Sarmatian period was more than 3 ‰ higher than the average 13C isotope content in herbivores of the same period (see Fig. 3). Such a trophic step as the one from Kichmalka II may indicate the presence of plants of C4-type photosynthesis in the diet of individual persons. The difference between the average δ15N values of humans and animals was within the trophic step. Noteworthy is the isotopic signal of small ruminants (δ15N = 10.9 ‰), anomalous for this region (see Fig. 3). Such high content of a heavy nitrogen isotope may indicate the arrival of the animals from more arid steppe region (Schulting, Richards, 2016; Shishlina, Sevastyanov, Kuznetsova, 2018). Since ∆δ15Nhuman-animal was within the trophic step, it can be concluded that the role of moved livestock (that is moved from other region) in the nutrition of people of the Sarmatian time was small.
The values of individuals from the Alanian burials at both cemeteries testify to the similarity of plant food sources. The average δ13C value in bone collagen of the Alans from Klin-Yar III was over 18 ‰. As at the Kichmalka II site, ∆δ13Chuman-animal (1.9 ‰) did not exceed the trophic step (see Fig. 4), and ∆δ15Nhuman-animal was over 5 ‰ (5.2 ‰). The difference in the human diet of the Alanian period from Kichmalka II and Klin-Yar III was the higher content of a heavy nitrogen isotope at the latter site (by 0.8 ‰). It is difficult to establish the reason for these differences because of the limited amount of data on isotopic signature of domestic animals, especially those from Kichmalka II. This could have resulted from greater availability of food resources of animal origin among the Alans from Klin-Yar III.
The higher δ15N values in humans may be associated with their social status (Knipper et al., 2015). According to scholars, the persons buried in the Sarmatian and Alanian periods came from the same site of elite burials at the Klin-Yar III cemetery (Belinskij, Härke, 2018: 32–34). In this regard, it can be assumed that variety of livestock and availability of meat and dairy products testify to the high social status of the buried. These assumptions require further research.
According to the results of isotope analysis, the average δ13C value was –28.4 ‰ in modern dung (see Table 3), and was –20.26 and –20.02 ‰ in bone collagen of domestic animals (isotopic signatures of the moved livestock with a high content of heavy nitrogen isotope were not taken into account) from the Kichmalka II and Klin-Yar III sites, respectively. The difference between the isotopic signatures of plants and herbivore collagen (∆δ13Cplant-animal) was 5 ‰ (Ambrose, 1993). In our case, taking into account the Suess effect, the trophic step was larger, reaching about 6 ‰.
The difference of about 2 ‰ between the droppings collected at the heights of ca 2000 and 900 m above sea level can be observed. There are not enough samples to draw definitive conclusions, but this does not contradict the literature data on the decreased content of heavy nitrogen isotope in plants with height (Huber et al., 2007). Low δ15N values of small ruminants of the Alanian period from Kichmalka II (3.73–3.8 ‰) and horses of the same period from Klin-Yar III (2.36–3.12 ‰) (see Fig. 4) may be associated with vertical transhumance, that is, seasonal grazing of these animals in alpine meadows, the vegetation of which contained nitrogen enriched with isotope 14N (Makarov et al., 2020: Tab. 3).
The assumption of the existence of transhumance is also supported by archaeological evidence. Comprehensive studies of stone enclosures for livestock at the heights of 2100–2200 m above sea level south of the Kislovodsk Basin, carried out by the Kislovodsk Expedition of the Institute of Archaeology of the Russian Academy of Sciences, have made it possible to date these structures to the Early Middle Ages and Late Bronze–Early Iron Age (Korobov et al., 2018). The absence of collagen samples from animals of the Koban period enriched in a light isotope of nitrogen at both sites might have been caused by insufficient amount of analyzed evidence.
Conclusions
The study of the isotopic composition of collagen in bone tissue of humans and animals from the Kichmalka II burial ground has made it possible to establish the features of food sources and resource zones among the carriers of the Koban culture, people of the Sarmatian period, and Alans. Because of small number of the studied groups of individuals, we can indicate only the most significant differences between food resources used by the representatives of different periods. People of the Koban and Sarmatian periods used C3-and C4-plants for food. Judging by the δ13C values, people of the Early Middle Ages and their animals were included in trophic relations based primarily on C3-plants.
Comparison of our data with the previously published results related to osteological evidence from the Klin-Yar III cemetery has confirmed the important role of millet in the economy and nutrition of the Koban people. Differences between the isotopic parameters of the Kobans buried at these two cemeteries might have been caused by the discrepancy in the chronology of their formation. The Koban burials at Kichmalka II belong to the 7th to early 5th centuries BC—the time of increased moistening.
Data on the isotopic composition of modern dung collected at various heights has made it possible to link low δ15N values of the early medieval small ruminants (Kichmalka II) and horses (Klin-Yar III) with transhumance among the Alans.
