Реконструкция питания и особенностей хозяйства населения Центрального Предкавказья I тыс. до н.э. - I тыс. н.э. по данным изотопного анализа коллагена остеологических материалов из могильника Кичмалка II

Автор: Бабенко А.Н., Добровольская М.В., Васильева Е.Е., Коробов Д.С.
Журнал: Археология, этнография и антропология Евразии @journal-aeae-ru
Рубрика: Эпоха палеометалла
Статья в выпуске: 4 т.49, 2021 года.
Бесплатный доступ
Системы расселения и хозяйствования населения предгорий Центрального Кавказа в железном веке и раннем Средневековье отражают сложные региональные культурные процессы. Палеоэкологические подходы, основанные на анализе изотопного состава углерода и азота в местных экосистемах, являются новой перспективой изучения природно-климатических условий, антропогенного воздействия, традиций землепользования и питания. Рассмотрен изотопный состав коллагена костей 19 человек и 11 животных из могильника Кичмалка II, оставленного носителями кобанской и аланской культур, а также населением сарматского времени. Изотопный состав углерода погребенных аланской эпохи указывает на практически исключительное использование людьми и домашними животными растений С3-типа фотосинтеза. Люди кобанского и сарматского времени включали в свой рацион С4-растения (просо, Panicum miliaceum), что обусловило превышение трофического шага по углероду (∆δ13Счеловек-животное). Установлены статистически достоверные различия в составе углерода, отражающие, вероятно, две пищевые модели в среде кобанского населения. Значения ∆δ15Nчеловек-животное находятся в пределах трофического шага и свидетельствуют о значительном потреблении мясо-молочной продукции всеми рассматриваемыми группами. Сравнение с данными по могильнику Клин-Яр III выявило различия в изотопных подписях людей и домашних животных кобанского времени. К основным причинам этого можно отнести изменение климата в раннем железном веке и разную долю проса в питании кобанцев. Низкое содержание δ15N (менее 4 ‰) в коллагене костей коз, овец и лошадей аланского времени, возможно, отражает практику отгонного альпийского скотоводства
Изотопный состав углерода и азота, северный кавказ, кобанская культура, сарматское время, аланы, трофические связи
Короткий адрес: https://sciup.org/145146502
IDR: 145146502 | УДК: [902/904.67+571.471.3] | DOI: 10.17746/1563-0102.2021.49.4.080-090
Текст научной статьи Реконструкция питания и особенностей хозяйства населения Центрального Предкавказья I тыс. до н.э. - I тыс. н.э. по данным изотопного анализа коллагена остеологических материалов из могильника Кичмалка II
Изотопный состав биологических останков людей, животных и растений запечатлевает обстоятельства их жизни, в связи с чем анализ этих параметров получил столь широкое распространение в археологической науке. В задачи нашего исследования входило выяснение традиций питания и хозяйственной деятельности у населения предгорий Северного Кавказа в I тыс. до н.э. – I тыс. н.э. на основе данных об изотопном составе углерода и азота в коллагене костной ткани людей и животных из погребений могильника Кичмалка II, а также данных, полученных при изучении растительных материалов из помета современного мелкого рогатого скота (МРС). Как известно, реконструкции рационов человека и животных базируются на общей закономерности динамики (фракционирования) изотопного состава азота и углерода в трофических цепях экосистем: при переходе с одного уровня на другой он закономерно меняется [Ambrose, 1993]. От обоснованности наших знаний о величине трофического шага зависит достоверность реконструкции [Добровольская и др., 2020]. Также важно отметить, что как общие изотопные характеристики экосистем, так и размеры трофических шагов внутри этих систем не могут быть выражены едиными величинами. Поэтому каждое исследование изотопного состава биологических материалов из археологических памятников сочетает в себе общую характеристику древней экосистемы с индивидуальными биоархеологически-ми реконструкциями.
Население предгорий Северного Кавказа с древности практиковало комплексное сельское хозяйство, сочетающее занятия скотоводством и земледелием
[Калоев, 1981, с. 7–32]. Однако сведения об особенностях его ведения в определенных сообществах имеют достаточно общий характер и зачастую лишены конкретного наполнения. Так, например, долгое время бытовало представление о том, что основным занятием носителей кобанской культуры было скотоводство, а земледелие являлось вспомогательным [Козенкова, 1989, с. 65; Марковин, Мунчаев, 2003, с. 166–168]. Более подробные характеристики давались хозяйствованию аланского населения в поздний период его истории на Северном Кавказе. Основным видом деятельности алан в этот период считалось земледелие в сочетании с отгонным скотоводством [Кузнецов, 1971, с. 47–147].
В последние годы более детально изучены свидетельства занятия земледелием и скотоводством населения Кисловодской котловины на протяжении длительного времени – с начала I тыс. до н.э. по конец I тыс. н.э. Почвенно-археологические исследования земледельческих угодий разных форм (террасы, пахотные наделы, участки с межевыми стенками), изучение сохранившихся в горах каменных загонов для скота позволили более подробно охарактеризовать практики интенсивного земледелия носителей кобан-ской культуры [Борисов, Коробов, 2013, с. 182–195], а также комплексность хозяйства аланских племен, в котором важную роль помимо земледелия играло и отгонное скотоводство [Коробов, 2017, с. 199–227; Коробов и др., 2018]. Эти исследования подкрепляются результатами анализа остеологического материала с раннесредневековых городищ I тыс. н.э. и макроостатков культурных злаков с поселений и сельскохозяйственных угодий в их окрестностях [Коробов, 2017, с. 202–204, 214–215; Сергеев, 2021].
Изучение хозяйственной деятельности и питания населения различных исторических эпох региона с использованием подходов изотопной биогеохимии составляет особое направление. Статья Т. Хайэма с соавторами [Higham et al., 2010] посвящена выяснению причин значительного резервуарного эффекта, обнаруженного при датировании костных останков из могильника Клин-Яр III в Кисловодской котловине. В качестве инструмента исследования авторы используют величины трофических шагов в экосистемах, привлекая данные по археологическим памятникам и современным материалам. В том же году были опубликованы результаты изучения изотопного состава углерода и азота в коллагене костной ткани индивидов из погребений энеолита и бронзового века на Северном Кавказе [Hollund et al., 2010]. В работе обсуждалась роль локальной среды, способной существенно повлиять на его специфику у местных обитателей (как людей, так и животных). Итоги масштабных изотопных исследований опубликованы международным коллективом, изучавшим питание и образ жизни некоторых групп населения различных периодов бронзового века на Северном Кавказе [Knipper et al., 2018, 2020]. В этом многоплановом исследовании, в частности, подтверждено значительное влияние локальных эко-
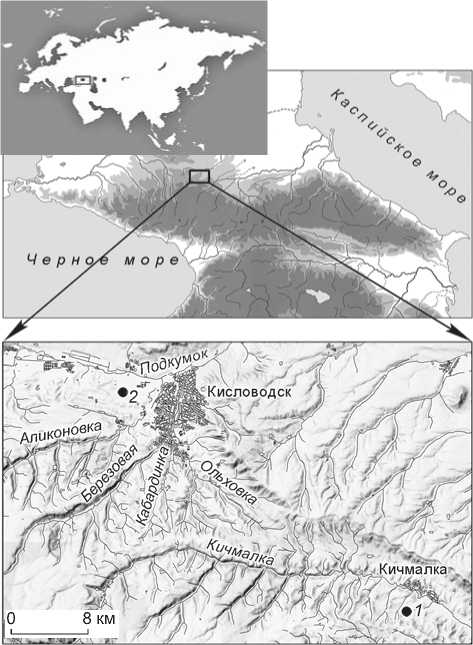
Рис. 1. Расположение могильников Кичмалка II ( 1 ) и Клин-Яр III ( 2 ) на Северном Кавказе.
систем на величины как изотопных соотношений, так и трофических шагов в данных системах.
Наша гипотеза состоит в том, что традиции носителей кобанской культуры, групп сарматского времени и аланов проявлялись и в особенностях ведения сложного неспециализированного хозяйства. Это можно установить по данным об изотопном составе коллагена костной ткани людей и животных из погребений.
Материалы
В основу исследования положены материалы могильника Кичмалка II, исследованного в 2006–2010 гг. экспедицией Государственного Эрмитажа под руководством Е.Е. Васильевой [Васильева, 2009, 2010, 2012; Васильева, Ахмедов, 2015]. Образцы костной ткани были отобраны из погребений кобанской культуры (VII – начало V в. до н.э.), сарматского времени (I– II вв. н.э.) и раннего Средневековья (вторая половина V – начало VI в. н.э.). Могильник расположен в предгорьях Северного Кавказа (Зольский р-н Кабардино-Балкарской Республики), на высотах чуть более 1 000 м (рис. 1).
В анализ вошло 32 образца: 8 от скелетных останков мужчин (индивид из погр. 24 представлен двумя образцами – губчатой и компактной костной ткани, что позволяет оценить стабильность изотопного состава коллагена в тканях с разной скоростью костных перестроек), 10 – женщин (индивид из погр. 28 также представлен двумя образцами), 3 – детей в возрасте 5–7 лет и ок. 12 лет; 5 от костей МРС, 4 – лошадей, 2 – мелких землеройных зверьков, не имеющих отношение к древности (табл. 1, 2). Кроме того, получены данные по четырем современным образцам помета овец, собранного на разных высотах: на плато Берма-мыт (Карачаево-Черкесская Республика) и в окрестности г. Кисловодска (табл. 3). Для анализов отбирались средние образцы из пяти – семи пометин.
Методы
Пробоподготовка проводилась в лаборатории контекстуальной антропологии Института археологии РАН. Фрагменты костной ткани промывались в дистиллированной воде, доводились до постоянного веса. Сухой фрагмент помещали в 1М соляную кислоту в соотношении не менее 1 г на 50 мл и оставляли при температуре +3 °С до полной деминерализации. Далее образец многократно промывался дистиллированной водой до pH = 7. На следующем этапе его на 24 ч помещали в 0,1М раствор щелочи, после чего вновь промывали. Затем образец помещали в раствор соляной кислоты с pH = 2,5 (10 мл) и переносили в термостат,
Таблица 1. Индивидуальные значения δ13С и δ15N образцов коллагена костной ткани погребенных из могильника Кичмалка II
|
Год раскопок |
Номер погребения/ скелета |
Пол |
Возраст |
δ 13 C |
δ 15 N |
%C |
%N |
C/N atom. |
|
Кобанская культура |
||||||||
|
2007 |
3 |
Муж. |
Старше 40 |
–16,84 |
10,28 |
43,3 |
15,2 |
3,3 |
|
2008 |
8 |
» |
30–39 |
–17,66 |
8,78 |
41,9 |
15,4 |
3,2 |
|
2008 |
10 |
Жен. |
20–25 |
–18,26 |
10,07 |
41,9 |
15,3 |
3,2 |
|
2009 |
20 |
Муж. |
Старше 50 |
–18,16 |
10,23 |
42,6 |
15,7 |
3,2 |
|
2009 |
21 |
Жен. (?) |
10–12 |
–18,66 |
10,11 |
44,7 |
15,9 |
3,3 |
|
2009 |
24 (а) |
Муж. |
Старше 45 |
–18,61 |
9,62 |
43,3 |
15,0 |
3,4 |
|
2009 |
24 (б) |
» |
Старше 45 |
–18,78 |
9,77 |
43,5 |
14,9 |
3,4 |
|
2009 |
25 |
Жен. |
Около 20 |
–16,68 |
9,42 |
41,8 |
15,1 |
3,2 |
|
2009 |
26 |
» |
Старше 50 |
–17,91 |
10,43 |
44,7 |
15,7 |
3,3 |
|
2009 |
23 |
? |
5–7 |
–16,97 |
9,39 |
36,3 |
12,3 |
3,4 |
|
2010 |
30 |
Муж. (?) |
Около 12 |
–17,32 |
9,85 |
41,4 |
14,2 |
3,4 |
|
Сарматское время |
||||||||
|
2010 |
29/1 |
Муж. |
Старше 50 |
–17,92 |
9,32 |
42,0 |
15,2 |
3,2 |
|
2010 |
29/2 |
Жен. |
Старше 60 |
–18,07 |
8,72 |
42,2 |
15,3 |
3,2 |
|
2009 |
22 |
» |
Около 20 |
–17,91 |
9,26 |
42,4 |
15,1 |
3,3 |
|
2010 |
31 |
» |
30–49 |
–16,03 |
10,88 |
43,7 |
15,5 |
3,3 |
|
Аланская культура |
||||||||
|
2010 |
28 (а) |
» |
40–49 |
–18,16 |
9,45 |
38,7 |
13,9 |
3,3 |
|
2010 |
28 (б) |
» |
40–49 |
–17,73 |
9,42 |
44,0 |
15,3 |
3,3 |
|
2010 |
33/1 |
Муж. |
17–20 |
–18,30 |
9,15 |
42,4 |
15,7 |
3,2 |
|
2010 |
33/2 |
Жен. |
16–18 |
–18,86 |
9,99 |
42,6 |
15,6 |
3,2 |
|
2010 |
34/1 |
Муж. |
20–25 |
–18,04 |
11,18 |
43,6 |
15,8 |
3,2 |
|
2010 |
34/2 |
Жен. |
30–39 |
–18,10 |
9,63 |
42,6 |
15,2 |
3,3 |
Таблица 2. Индивидуальные значения δ13С и δ15N образцов коллагена костной ткани животных из могильника Кичмалка II
|
Год раскопок |
Место обнаружения |
Животное |
δ 13 C |
δ 15 N |
%C |
%N |
C/N atom. |
Культурная принадлежность погребения |
|
2010 |
Погр. 34 |
Норный зверек |
–23,80 |
5,36 |
47,7 |
12,9 |
4,3 |
Аланское |
|
2010 |
Погр. 34, разруш. часть |
МРС |
–19,95 |
3,73 |
40,1 |
14,4 |
3,3 |
» |
|
2010 |
Погр. 33 |
» |
–19,91 |
3,80 |
41,7 |
15,1 |
3,2 |
» |
|
2010 |
Рядом с погр. 31 |
Лошадь |
–20,34 |
5,52 |
46,2 |
15,1 |
3,6 |
Сарматское |
|
2010 |
Погр. 29 |
МРС |
–19,77 |
4,75 |
42,4 |
15,3 |
3,2 |
» |
|
2009 |
Рядом с погр. 24 |
Лошадь |
–20,46 |
4,67 |
41,9 |
14,4 |
3,4 |
Кобанское |
|
2009 |
То же |
» |
–20,84 |
4,90 |
42,9 |
15,2 |
3,3 |
» |
|
2009 |
Погр. 24 |
МРС |
–20,37 |
5,57 |
41,2 |
15,0 |
3,2 |
» |
|
2007 |
Погр. 3 |
Лошадь |
–20,68 |
5,22 |
41,2 |
14,8 |
3,2 |
» |
|
2009 |
Погр. 24 |
Норный зверек |
–18,73 |
7,09 |
39,5 |
13,3 |
3,5 |
» |
|
2010 |
Погр. 32 |
МРС |
–20,01 |
5,79 |
32,1 |
11,2 |
3,4 |
» |
Таблица 3. Значения δ13С и δ15N образцов современного помета МРС
Пробы современного помета обрабатывались горячим 5%-ым раствором HCl и промывались через сито (0,25 мм) для удаления минеральных зерен и крупных растительных остатков с целью наилучшего усреднения материала. Далее образцы многократно промывались дистиллированной водой и высушивались. Таким образом, полученные результаты будут отражать содержание стабильных изотопов углерода и азота непосредственно в усредненной пробе растительных остатков. Анализируемая навеска составляла ок. 1 500 мкг.
Изотопный состав образцов определялся с помощью масс-спектрометра (Thermo Finnigan Delta V Plus, Центр коллективного пользования Института проблем экологии и эволюции РАН, Москва). Для статистического сравнения выборок использовался критерий Манна–Уитни U , т.к. распределение параметров носит ненормальный характер.
Для определения вклада растительной и животной пищи в рацион человека необходимо оценивать разни- цу между изотопными данными человека, культурных растений и животных, которых он мог употреблять в пищу. Разница между изотопными подписями (∆δ13С и ∆δ15N) продуцентов (растения)/консументов первого порядка (травоядные), а также консументов первого (травоядные) и второго (хищники) порядков является трофическим шагом. Величина δ13С и δ15N в коллагене костей с каждым трофическим уровнем увеличивается на 0–2 ‰ [Bocherens, Drucker, 2003] и 3–5 ‰
(у человека даже до 6 ‰) [Ambrose, 1993; O’Connell et al., 2012] соответственно. По тому, на сколько человек–животное
и ∆δ15Nчеловек–животное отличаются от величины трофического шага, можно оценить вклад того или иного компонента в рацион человека. Ма- лые значения показателей чаще всего свидетельствуют о незначительной доле мясо-молочных продуктов, употребляемых человеком. Значения, превышающие величины трофического шага, могут указывать на другие источники питания [Ambrose, 1993; O’Brien, 2015].
Отсутствие изотопных данных по растительной пище древнего и средневекового человека в рассматриваемом регионе ограничивает возможности реконструкции его питания, поэтому мы можем лишь исходить из общепринятых положений об изотопных сигналах растений С3- и С4-типа фотосинтеза. Величина δ13С –18 ‰ маркирует начало употребления растений С4-типа [Pearson et al., 2007].
При сравнении с современными изотопными данными необходимо иметь в виду изменение в индустриальную и постиндустриальную эпоху изотопного состава атмосферы, которая обогащена древним углеродом с низкими величинами δ13С (эффект Зюсса). Значение δ¹³C атмосферного CO2, восстановленного по пузырькам керна льда, снизилось с –6 ‰ ок. 11 тыс. л.н. [Indermühle et al., 1999], до ок. –8 ‰ сегодня [White, Vaughn, Michel, 2015]. Этот сдвиг на 2 ‰ необходимо учитывать при интерпретации изотопного состава углерода археологических образцов.
Результаты
Углерод. Индивидуальная изменчивость изотопных соотношений углерода в коллагене костной ткани людей варьирует от –18,86 до –16,03 ‰ (см. табл. 1). Значения δ13С у погребенных кобанского и сарматского времени варьируют в довольно широких пределах – от –18,78 до –16,68 ‰ и от –18,07 до –16,03 ‰ соответственно. Носители аланской культуры характеризуются небольшой изменчивостью этого показателя – от –18,86 до –17,73 ‰.
Значения δ13С у травоядных домашних животных варьируют от –20,84 до –19,77 ‰ (см. табл. 2). Коллаген лошадей и МРС характеризуется единообразием изотопного состава углерода (от –20,84 до –20,34 ‰ и от –20,37 до –19,77 ‰ соответственно).
Азот. Значения δ15N в коллагене людей варьируют в пределах 8,72–11,18 ‰. Индивидуальная изменчивость показателя наименьшая у кобанцев (8,78–10,43 ‰). Диапазоны у аланов (9,15–11,18 ‰) и людей сарматского периода (8,72–10,88 ‰) шире (см. табл. 1). Группы статистически не различаются.
Изотопные соотношения азота в коллагене травоядных домашних животных варьируют от 3,73 до 5,79 ‰. Диапазон значений δ15N у лошадей 4,67– 5,52 ‰. У МРС кобанского времени этот показатель составляет 5,57–5,79 ‰. Содержание же изотопа 15N в коллагене костей МРС аланского периода минимально и не превышает 4 ‰ (см. табл. 2).
Индивидуальная изменчивость изотопных параметров не связана с полом и возрастом. Значимых различий не обнаружено.
Величина δ13С растительных остатков из современного помета изменяется от –27,91 до –28,68 ‰, а δ15N – от 2,45 до 4,44 ‰ (см. табл. 3).
Обсуждение
Варьирование в коллагене костной ткани носителей кобанской культуры δ13С и δ15N в широких пределах свидетельствует об индивидуальном разнообразии пищевых рационов. Изменчивость δ13С со ставляет 2,1 ‰. Можно выделить два облака данных (рис. 2): группа 1 (погр. 3, 8, 23, 25, 30) достоверно отличается от группы 2 (погр. 10, 20, 21, 24, 26) высокими значениями δ13С (˃ –17,7 ‰, p = 0,006). По δ15N они изменчивы, но статистически не различаются ( p ˃ 0,05). Следовательно, в основе питания каждой группы лежат разные растительные компоненты. Между выделенными группами можно провести условную границу, которая будет проходить в районе значения δ13С –18 ‰. Это позволяет считать основой этих различий употребление С4-растений.
Разница между средними значениями δ13С индивидов группы 1 и травоядных животных составляет более 2 ‰, а ∆δ15Nчеловек–животное – менее 5 ‰. Следовательно, первая больше трофического шага, а вторая находится в его пределах (рис. 2). Для части индивидов (группа 2, ∆δ С человек–животное и ∆δ N человек–животное ) мы можем реконструировать диету, основу которой составляли С3-растения и в достаточном количестве мясные продукты. В состав же пищи первой группы, в отличие от второй, вероятно, входило больше продуктов растительного происхождения с высоким содержанием изотопа 13С. Согласно археоботаническим данным, на территории Кисловодской котловины, в непосредственной близости от которой расположен памятник Кичмалка II, в кобанское время культивируемым С4-растением было просо ( Panicum miliaceum ) [Reinhold, Korobov, Belinskij, 2017, S. 242; Сергеев, 2021, табл. 8]. Можно предположить, что в рамках одной культуры существовали две пищевые модели, а не одна.
Между группами сарматского и аланского времени наблюдается значительное сходство ( p ˃ 0,05): у основной части индивидов значения δ13С и δ15N варьируют в пределах менее 1 ‰. Среди образцов людей сарматского времени один отличается более высокими значениями δ13С и δ15N. К сожалению, в нашем распоряжении лишь два изотопных сигнала домашних животных (МРС и лошадь) этого времени. Как и в случае с кобанцами, ∆δ13Счеловек–животное составляет более 2 ‰, а ∆δ15N человек–животное – менее 5 ‰ (рис. 3). Полученные данные указывают на употребление в пищу людьми сарматского времени в основном С3-растений
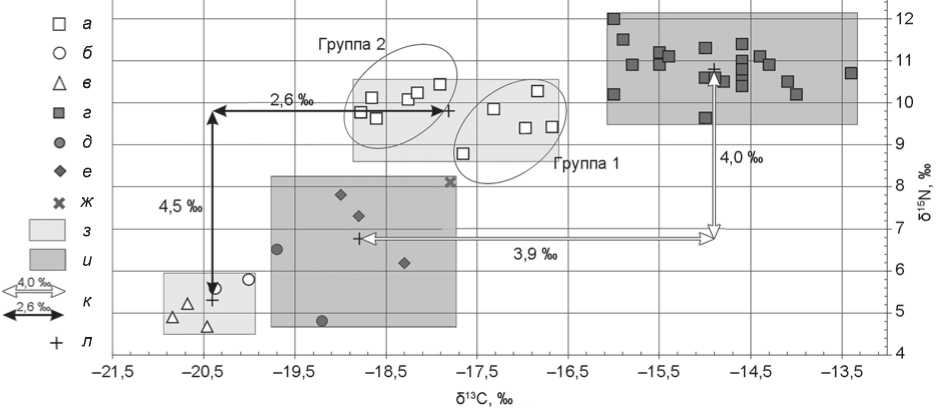
Рис. 2. Индивидуальные значения δ13С и δ15N образцов коллагена костной ткани людей и животных кобанского времени из могильников Кичмалка II и Клин-Яр III.
а – в – Кичмалка II: а – человек, б – МРС, в – лошадь; г–ж – Клин-Яр III: г – человек, д – МРС, е – КРС, ж – свинья; з , и – область изотопных сигналов коллагена костей из могильников соответственно Кичмалка II и Клин-Яр III; к – трофический шаг; л – сред-
нее значение.
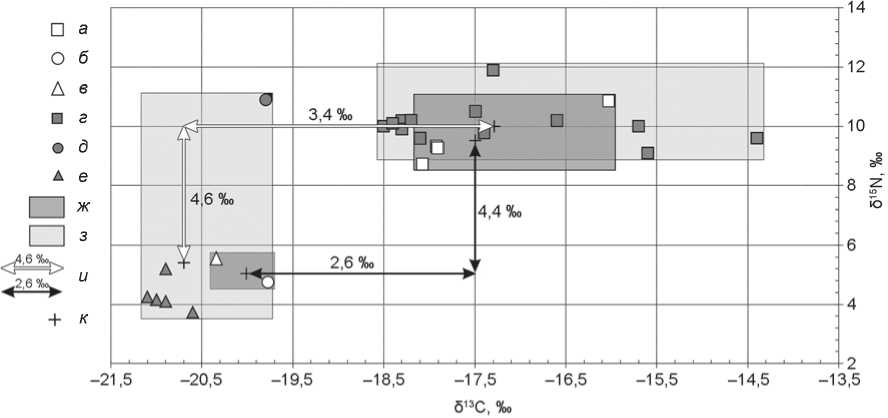
Рис. 3. Индивидуальные значения δ13С и δ15N образцов коллагена костной ткани людей и животных сарматского времени из могильников Кичмалка II и Клин-Яр III.
а – в – Кичмалка II: а – человек, б – МРС, в – лошадь; г–е – Клин-Яр III: г – человек, д – МРС, е – лошадь; ж , з – область изотопных сигналов коллагена костей из могильников соответственно Кичмалка II и Клин-Яр III; и – трофический шаг; к – среднее значение.
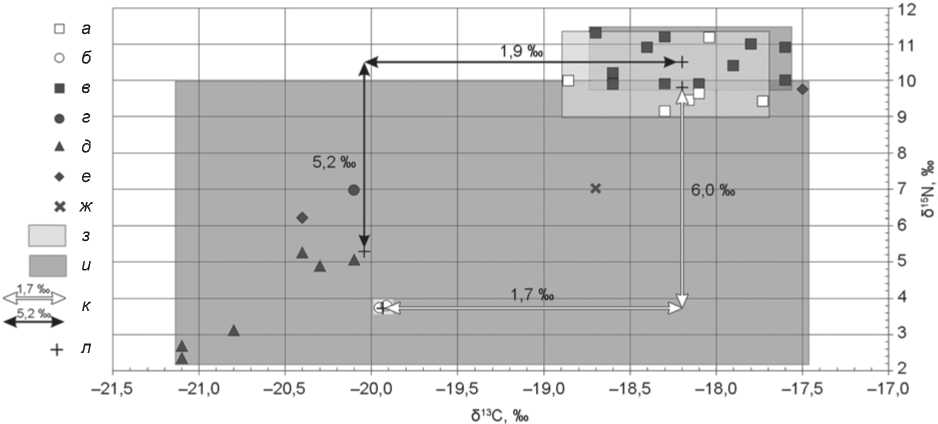
Рис. 4. Индивидуальные значения δ13С и δ15N образцов коллагена костной ткани людей и животных аланского времени из могильников Кичмалка II и Клин-Яр III.
а , б – Кичмалка II: а – человек, б – МРС; в–ж – Клин-Яр III: в – человек, г – МРС, д – лошадь, е – КРС, ж – свинья; з , и – область изотопных сигналов коллагена костей из могильников соответственно Кичмалка II и Клин-Яр III; к – трофический шаг; л – среднее значение.
и в достаточном количестве мяса и/или молока животных, питающихся ими. В рационе отдельных индивидов, возможно, была некоторая доля С4-растений (погр. 31).
В отличие от двух рассмотренных периодов, разница между средними значениями δ13С в коллагене костей людей и животных аланского времени не превышает трофического шага (рис. 4). Величины δ13С практически всех образцов (за исключением одно- го, погр. 28 (б)) указывают на диету, основанную на С3-растениях. Полученные данные не противоречат результатам археоботанических исследований на аланских поселениях V–VIII вв. н.э., расположенных на близлежащей к памятнику Кичмалка II территории Кисловодской котловины. В региональном археоботаническом комплексе этого времени преобладают пшеницы (65 %), а доля проса составляет лишь 9 % [Сергеев, 2021]. Можно констатировать до- минирование С3-растений в рационе аланского населения V–VIII вв. н.э.
Еще одним отличием раннесредневековых образцов является бóльшая разница между средними значениями δ15N людей и животных – 6 ‰. Она находится в пределах трофического шага [O’Connell et al., 2012]. Следует отметить, что кости МРС из погребений алан характеризуются самым низким азотным соотношением (менее 4 ‰). К сожалению, у нас нет достаточного количества изотопных данных по домашним животным. Однако нужно принять во внимание поселенческие материалы с территории Кисловодской котловины, где на многочисленных обследованных поселениях V–VIII вв. присутствовали в качестве кухонных остатков кости не только мелкого, но и крупного рогатого скота, лошадей и свиней [Коробов, 2017, с. 202–203]. Поэтому можно с уверенностью говорить, что разница в 6 ‰ обусловлена недостатком изотопных данных. Итак, аланская группа оказывается наиболее компактной по характеристикам изменчивости источников пищевых ресурсов.
Изучение изотопного со става коллагена костей людей и животных на памятниках Северного Кавказа находится на начальном этапе, который сводится к накоплению данных и попытке их интерпретации, что в условиях разнообразных ландшафтов является непростой задачей. Для реконструкции рациона человека необходимо учитывать большой набор изотопных сигналов различных ресурсов. В связи с этим важно сравнить наши результаты с опубликованными ранее. Сопоставительным материалом послужили данные об изотопном составе коллагена костной ткани людей и животных из могильника Клин-Яр III в Кисловодской котловине [Higham et al., 2010]. Памятник расположен на высоте 800 м над ур. м. на расстоянии ок. 25 км от Кичмалки II (см. рис. 1). На могильнике Клин-Яр III также представлены погребения трех культурно-хронологических периодов. Однако, в отличие от Кичмалки II, остеологический материал, относящийся к кобанской культуре, датируется более ранним временем – X–VII вв. до н.э. [Белинский, Дударев, 2015, с. 386–397].
Расположение индивидуальных данных на диаграмме соотношения значений δ13С/δ15N позволяет оценить особенности каждой из культурно-хронологических групп двух погребальных памятников (см. рис. 2–4). Индивиды из кобанских погребений Кичмалки II и Клин-Яра III достоверно отличаются друг от друга величинами и δ15N (p = 0,000028), и δ13С (p = 0,000003). Области индивидуальных значений δ13С двух этих групп даже не перекрываются (см. рис. 2). Люди сарматского периода, захороненные на могильниках Кичмалка II и Клин-Яр III, характеризуются примерно одинаковыми величинами (p ˃ 0,05) и большим диапазоном изменчивости изо- топного состава углерода (от –18,4 до –15,7 ‰). Изотопные показатели аланов из погребений обоих памятников схожи по составу углерода (p ˃ 0,05), однако значения δ15N у индивидов из Клин-Яра III статистически выше (p = 0,035).
Отличия изотопных показателей людей и животных кобанского времени из двух могильников выражены четко, что может быть обусловлено различными ландшафтными и климатическими условиями в окрестностях памятников [Marshall, Brooks, Lajtha, 2007] и/или разницей в питании [Ambrose, 1993]. Однако Кичмалка II и Клин-Яр III расположены недалеко друг от друга в сходных ландшафтах, поэтому различия изотопного состава не связаны со средой обитания. В пользу последнего говорит и то, что в других группах аналогичных отличий не наблюдается. Надежные климатиче ские реконструкции для изучаемой территории X–V вв. до н.э. отсутствуют. Однако синхронность динамики ледников Северного Кавказа и Альп в последнем тысячелетии [Solomina et al., 2016] позволяет рассматривать климатические изменения региона, опираясь на зарубежные работы. Согласно исследованиям ледников и уровня озер в Альпах [Holzhauser, Magny, Zumbuühl, 2005; Ivy-Ochs et al., 2009; Wirth et al., 2013], вторая половина I тыс. до н.э. характеризуется холодными и влажными условиями. Степень увлажненности, однако, на севере и юге Альп различна в связи со смещением к югу западного переноса в это время. Однозначно нельзя утверждать, но можно предположить, что в конце рассматриваемого периода (время погребения кобан-цев на могильнике Кичмалка II) количество осадков в регионе могло увеличиться, как и на юге Альп [Wirth et al., 2013]. При возрастании увлажненности δ13С и δ15N в С3-растениях уменьшаются [Marshall, Brooks, Lajtha, 2007]. То есть изотопные сигналы коллагена костей людей и животных кобанского времени с памятника Кичмалка II должны отличаться более низкими значениями этих показателей, что можно наблюдать на рис. 2. Косвенным аргументом в пользу данного предположения является прекращение существования кобанских поселений в Кисловодской котловине в середине I тыс. до н.э. вследствие спровоцированных увлажнением климата катастрофических изменений агроландшафтов [Борисов, Коробов, 2013, с. 191–195].
Индивидуальная изменчивость внутри обеих групп кобанского населения (Кичмалка II и Клин-Яр III) по величинам δ13С высока и, возможно, предполагает различные растительные пищевые ресурсы. Разница между средними значениями δ13C людей и домашних животных с памятника Клин-Яр III в 2 раза больше трофического шага. Если рассмотреть опубликованные данные по содержанию изотопов углерода в коллагене костей людей с других памятников Север- ного Кавказа эпохи бронзы [Knipper et al., 2020, tab. 3], то можно отметить, что среднее значение δ13C у носителей кобанской культуры из Кичмалки II (–17,8 ‰) близко к литературным данным (–18,4 ÷ –19,4 ‰), а из Клин-Яра III – значительно выше (–14,3 ‰). Последнее свидетельствует о бóльшей доли проса в питании людей. Таким образом, различия изотопных подписей индивидов из кобанских погребений двух памятников связаны как с разными климатическими условиями, так и с пищевыми предпочтениями отдельных групп людей.
Среднее значение δ13С у погребенных на могильнике Клин-Яр III в сарматский период более чем на 3 ‰ превышает среднее содержание изотопа 13С у травоядных животных того же времени (см. рис. 3). Такой трофический шаг, как и на памятнике Кичмалка II, может свидетельствовать о присутствии в рационе отдельных индивидов растений С4-типа фотосинтеза. Разница между средними значениями δ15N людей и животных находится в пределах трофического шага. Следует отметить аномальный для этого региона изотопный сигнал МРС (δ15N = 10,9 ‰) (см. рис. 3). Такое высокое содержание тяжелого изотопа азота может указывать на пригон животного из более засушливого степного региона [Schulting, Richards, 2016; Shishlina, Sevastyanov, Kuznetsova, 2018]. Так как ∆δ15Nчеловек–животное находится в пределах трофического шага, можно сказать, что роль пригонного скота в питании людей сарматского времени была небольшой.
Показатели индивидов из аланских погребений обоих могильников свидетельствуют о сходстве растительных пищевых ресурсов. Среднее значение δ13С в коллагене костей алан из Клин-Яра III более 18 ‰. Как и на памятнике Кичмалка II, ∆δ13Счеловек–животное (1,9 ‰) не превышает трофического шага (см. рис. 4), а ∆δ15Nчеловек–животное более 5 ‰ (5,2 ‰). Разница в питании людей аланского времени из Кичмалки II и Клин-Яра III выражается в большем содержании тяжелого изотопа азота у последних (на 0,8 ‰). Установить причину различий сложно из-за ограниченного количества данных по изотопной подписи домашних животных, особенно из могильника Кичмалка II. Возможно, это объясняется большей доступностью пищевых ресурсов животного происхождения у алан из Клин-Яра III.
Более высокие значения δ15N у людей могут быть связаны с их социальным положением [Knipper et al., 2015]. По данным исследователей, погребенные в сарматский и аланский периоды происходят с одного участка элитных погребений могильника Клин-Яр III [Belinskij, Härke, 2018, p. 32–34]. В связи с этим можно предположить, что разнообразие домашнего скота, доступность мясо-молочной продукции являются свидетельством высокого социального положения погребенных. Высказанные предположения требуют дальнейших исследований.
Согласно результатам изотопного анализа среднее значение δ13C современного помета составляет –28,4 ‰ (см. табл. 3), а в коллагене костей домашних животных с памятников Кичмалка II и Клин-Яр III – соответственно –20,26 и –20,02 ‰ (изотопные подписи пригонных животных с высоким содержанием тяжелого изотопа азота не учитывались). Разница между изотопными подписями растений и коллагена травоядного (∆δ13Срастение–животное) составляет 5 ‰ [Ambrose, 1993]. В нашем случае, с учетом эффекта Зюсса, трофический шаг больше – ок. 6 ‰.
Можно отметить разницу ок. 2 ‰ между пометом, собранным на высотах ок. 2 000 и 900 м над ур. м. Образцов недо статочно, чтобы делать выводы, однако это не противоречит литературным данным об уменьшении содержания тяжелого изотопа азота в растениях с высотой [Huber et al., 2007]. Возможно, низкие значения δ15N МРС аланского периода из Кичмалки II (3,73–3,8 ‰) и лошадей того же времени из Клин-Яра III (2,36–3,12 ‰) (см. рис. 4) связаны с существованием в раннем Средневековье отгонного скотоводства, т.е. с сезонным выпасом этих животных на альпийских лугах, растительность которых содержит азот, обогащенный изотопом 14N [Макаров и др., 2020, табл. 3].
Предположение о существовании отгонного скотоводства подкрепляется также археологическими данными. Результаты проведенных Кисловодской экспедицией Института археологии РАН комплексных исследований каменных загонов для скота на высотах 2 100–2 200 м над ур. м. южнее Кисловодской котловины позволяют датировать эти сооружения ранним Средневековьем и поздним бронзовым – ранним железным веками [Коробов и др., 2018]. Отсутствие образцов коллагена животных кобанского времени, обогащенных легким изотопом азота, на обоих памятниках может быть связано с недостаточным количеством анализируемого материала.
Заключение
Исследование изотопного состава коллагена костной ткани людей и животных из могильника Кичмалка II позволило охарактеризовать особенности источников питания и ресурсных зон носителей кобанской культуры, людей сарматского времени и алан. Ввиду малочисленности изученных групп индивидов мы можем констатировать лишь наиболее значимые различия между пищевыми ресурсами представителей разных эпох. Люди кобанского и сарматского времени использовали в пищу С3- и С4-растения. Судя по величинам δ13C, представители раннего Средневековья и их животные были включены в трофические связи, базировавшиеся в большей степени на С3-растениях.
Сопоставление наших данных с опубликованными ранее результатами изучения остеологических материалов из могильника Клин-Яр III подтвердило важную роль проса в хозяйстве и питании носителей кобанской культуры. Различия между изотопными показателями кобанцев, погребенных на этих двух могильниках, вероятно, обусловлены расхождением в хронологии их формирования. Кобанские погребения Кичмалки II относятся к VII – началу V в. до н.э. – времени повышения увлажнения.
Привлечение данных об изотопном составе современного помета, собранного на разных высотах, позволило связать низкие величины δ15N раннесредневековых МРС (Кичмалка II) и лошадей (Клин-Яр III) с использованием аланами отгонных форм скотоводства.
Список литературы Реконструкция питания и особенностей хозяйства населения Центрального Предкавказья I тыс. до н.э. - I тыс. н.э. по данным изотопного анализа коллагена остеологических материалов из могильника Кичмалка II
- Белинский А.Б., Дударев С.Л. Могильник Клин-Яр III и его место среди древностей Кавказа и Юго-Восточной Европы начала эпохи раннего железа. – Ставрополь: Дизайнстудия Б, 2015. – 446 с.
- Борисов А.В., Коробов Д.С. Древнее и средневековое земледелие в Кисловодской котловине: итоги почвенно-археологических исследований. – М.: Таус, 2013. – 272 с.
- Васильева Е.Е. Могильник Кичмалка II – новый памятник кобанской культуры на Северном Кавказе // Лавровский сборник: мат-лы XXXIII Среднеазиатско-Кавказских чтений, 2008–2009 гг.: К столетию со дня рождения Л.П. Лаврова: Этнология, история, археология, культурология. – СПб.: МАЭ РАН, 2009. – С. 42–44.
- Васильева Е.Е. Исследования кобанского могильника Кичмалка II в Кабардино-Балкарии // Проблемы хронологии и периодизации археологических памятников и культур Северного Кавказа: XXVI «Крупновские чтения» по археологии Северного Кавказа: тез. докл. Междунар. науч. конф. – Магас: Пилигрим, 2010. – С. 80–82.
- Васильева Е.Е. Погребальный комплекс аланской культуры на Северном Кавказе: Вторая половина V – первая половина VI века н.э. // Кочевники Евразии на пути к империи: Из собрания Государственного Эрмитажа: каталог выставки. – СПб.: Славия, 2012. – С. 176–179.
- Васильева Е.Е., Ахмедов И.Р. Новое погребение аланской знати постгуннского времени из Кабардино-Балкарии // Социальная стратификация населения Кавказа в конце античности и начале средневековья: археологические данные: мат-лы Междунар. науч. конф. (Сухум, 31 мая – 5 июня 2015 г.). – М.: ИА РАН, 2015. – С. 13–16.
- Добровольская М.В., Тиунов А.В., Крылович О.А., Кузьмичева Е.А., Решетова И.К., Савинецкий А.Б., Свиркина Н.Г., Смирнов А.Л. Изотопные маркеры экосистем и питания средневекового сельского населения лесной зоны европейской части России // РА. – 2020. – № 3. – C. 79–95.
- Калоев В.Б. Земледелие народов Северного Кавказа. – М.: Наука, 1981. – 248 с.
- Козенкова В.И. Кобанская культура: Западный вариант. – М.: Наука, 1989. – 196 с.
- Коробов Д.С. Система расселения алан Центрального Предкавказья в I тыс. н.э. (ландшафтная археология Кисловодской котловины). – М.; СПб.: Нестор-История, 2017. – Т. 1. – 384 с.
- Коробов Д.С., Борисов А.В., Бабенко А.Н., Сергеев А.Ю., Чернышева Е.В. Комплексное исследование каменных загонов в окрестностях Кисловодска // РА. – 2018. – № 2. – С. 113–129.
- Кузнецов В.А. Алания в X–XIII вв. – Орджоникидзе: Ир, 1971. – 248 с.
- Макаров М.И., Онипченко В.Г., Тиунов А.В., Малышева Т.И., Кадулин М.С. Почвы и азотное питание растений альпийских экосистем Северо-Западного Кавказа при многолетнем повышении доступности биофильных элементов // Почвоведение. – 2020. – № 8. – С. 1016–1024.
- Марковин В.И., Мунчаев Р.М. Северный Кавказ: Очерки древней и средневековой истории и культуры. – Тула: Гриф и К., 2003. – 240 с.
- Сергеев А.Ю. Зерно и уголь на древних и средневековых пашнях // Аналитические исследования лаборатории естественнонаучных методов. – М.: Таус, 2021. – Вып. 5. – С. 308–336.
- Ambrose S.H. Isotopic analysis of Paleodiets: methodological and interpretive considerations // Investigations of ancient human tissue: chemical analyses in anthropology. – N. Y.: Gordon and Breach Science Publisher Ltd., 1993. – P. 59–130.
- Belinskij A.B., Härke H. Ritual, society and population at Klin-Yar (North Caucasus): Excavations 1994–1996 in the Iron Age to early medieval cemetery. – Bonn: Dr. Rudolf Habelt GmbH, 2018. – 446 p.
- Bocherens H., Drucker D. Trophic level isotopic enrichment of carbon and nitrogen in bone collagen: Case studies from recent and ancient terrestrial ecosystems // Intern. J. of Osteoarchaeology. – 2003. – Vol. 13. – P. 46–53.
- DeNiro M.J. Postmortem preservation and alteration of invivo bone-collagen isotope ratios in relation to paleodietary reconstruction // Nature. – 1985. – Vol. 317. – P. 806–809.
- Higham T., Warren R., Belinskij A., Härke H., Wood R. Radiocarbon dating, stable isotope analysis, and diet-derived offsets in 14C ages from the Klin-Yar Site, Russian North Caucasus // Radiocarbon. – 2010. – Vol. 52, iss. 2. – P. 653–670.
- Hollund H.I., Higham T., Belinskij A . , Korenevskij S. Investigation of palaeodiet in the North Caucasus (South Russia) Bronze Age using stable isotope analysis and AMS dating of human and animal bones // J. of Archaeol. Sci. – 2010. – Vol. 37, iss. 12. – P. 2971–2983.
- Holzhauser H., Magny M., Zumbuühl H.J. Glacier and lake-level variations in west-central Europe over the last 3 500 years // The Holocene. – 2005. – Vol. 15. – P. 789–801.
- Huber E., Wanek W., Gottfried M., Pauli H., Schweiger P., Arndt S.K., Reiter K., Richter A. Shift in soil–plant nitrogen dynamics of an alpine–nival ecotone // Plant and Soil. – 2007. – Vol. 301. – P. 65–76.
- Indermühle A., Stocker T.F., Joos F., Fischer H., Smith H.J., Wahlen M., Deck B., Mastroianni D., Tschumi J., Blunier T., Meyer R., Stauffer B. Holocene carbon-cycle dynamics based on CO2 trapped in ice at Taylor dome, Antarctica // Nature. – 1999. – Vol. 398, iss. 6723. – P. 121–126.
- Ivy-Ochs S., Kerschner H., Maisch M., Christl M., Kubik P.W., Schluchter C. Latest Pleistocene and Holocene glacier variations in the European Alps // Quarter. Sci. Rev. – 2009. – Vol. 28. – P. 2137–2149.
- Knipper C., Held P., Fecher M., Nicklisch N., Meyer C., Schreiber H., Zich B., Metzner-Nebelsick C., Hubensack V., Hansen L., Nieveler E., Alt K.W. Superior in life-superior in death dietary distinction of Central European Prehistoric and Medieval elites // Current Anthropology. – 2015. – Vol. 4. – P. 579–589.
- Knipper C., Reinhold S., Gresky J., Belinskij A., Alt K.W. Economic strategies at Bronze Age and Early Iron Age upland sites in the North Caucasus: Archaeological and stable isotope investigations // Isotopic Investigations of Pastoralism in Prehistory. – L.; N. Y.: Routledge, 2018. – P. 123–140.
- Knipper C., Reinhold S., Gresky J., Berezina N., Gerling C., Pichler S.L., Buzhilova A.P., Kantorovich A.R., Maslov V.E., Petrenko V.G., Lyakhov S.V., Kalmykov A.A., Belinskiy A.B., Hansen S., Alt K.W. Diet and subsistence in Bronze Age pastoral communities from the southern Russian steppes and the North Caucasus // PLOS One. – 2020. – Vol. 15, iss. 10. – URL: https://doi.org/10.1371/journal.pone.0239861 (дата обращения: 15.03.2021).
- Marshall J.D., Brooks J.R., Lajtha K. Sources of variation in the stable isotopic composition of plants // Stable Isotopes in Ecology and Environmental Science. – Malden: Blackwell Publishing Ltd., 2007. – P. 22–60.
- O’Brien D.M. Stable Isotope Ratios as Biomarkers of Diet for Health Research // Annual Review of Nutrition. – 2015. – Vol. 35, iss. 1. – P. 565–594.
- O’Connell T.C., Kneale C.J., Tasevska N., Kuhnle G.G.C. The diet-body offset in human nitrogen isotopic values: A controlled dietary study // Am. J. of Phys. Anthropol. – 2012. – Vol. 149. – P. 426–434.
- Pearson J.A., Buitenhuis H., Hedges R.E.M., Martin L., Russel N., Twiss K.C. New light on early caprine herding strategies from isotope analysis: a case study from Neolithic Anatolia // J. of Archaeol. Sci. – 2007. – Vol. 34. – P. 2170–2179.
- Reinhold S., Korobov D., Belinskij A.B. Landschaftsarchäologie im Nordkaukasus: Studien zu einer neu entdeckten bronzezeitlichen Kulturlandschaft im Hochgebirge des Nordkaukasus. – Bonn: Habelt Verlag, 2017. – 514 S.
- Schulting R.J., Richards M.P. Stable isotope analysis of Neolithic to Late Bronze Age populations in the Samara Valley // A Bronze Age Landscape in the Russian Steppes: the Samara Valley Project. – Los Angeles: Cotsen Institute of Archaeology, 2016. – P. 127–148.
- Shishlina N., Sevastyanov V., Kuznetsova O. Seasonal practices of prehistoric pastoralists from the south of the Russian plain based on the isotope data of modern and archaeological animal bones and plants // J. of Archaeol. Sci.: Reports. – 2018. – Vol. 21. – P. 1247–1258.
- Solomina O., Bushueva I., Dolgova E., Jomelli V., Alexandrin M., Mikhalenko V., Matskovsky V. Glacier variations in the Northern Caucasus compared to climatic reconstructions over the past millennium // Global and Planetary Change. – 2016. – Vol. 140. – P. 28–58.
- Van Klinken G.J. Bone collagen quality indicators for Palaeodietary and radiocarbon measurements // J. of Archaeol. Sci. – 1999. – Vol. 26, iss. 6. – P. 687–695.
- White J., Vaughn B., Michel S. Stable Isotopic Composition of Atmospheric Carbon Dioxide (13C and 18O) from the NOAA ESRL Carbon Cycle Cooperative Global Air Sampling Network, 1990–2014, Version: 2015-10-26. – 2015. – URL: https://gml.noaa.gov/aftp/data/trace_gases/co2o18/fl ask/surface/README_surface_fl ask_co2o18.html (дата обращения: 15.03.2021).
- Wirth S.B., Glur L., Gilli A., Anselmetti F.S. Holocene Flood Frequency across the Central Alps – Solar Forcing and Evidence for Variations in North Atlantic Atmospheric Circulation // Quarter. Sci. Rev. – 2013. – Vol. 80. – P. 112–128.
