Результаты археопаразитологического и палинологического исследования культурных слоев античного поселения Артезиан (Восточный Крым)
")
Автор: Слепченко Сергей Михайлович, Винокуров Николай Игоревич, Бабенко Анна Николаевна, Хрусталев Александр Валерьевич, Иванов Сергей Николаевич
Журнал: Нижневолжский археологический вестник @nav-jvolsu
Рубрика: Статьи
Статья в выпуске: 2 т.21, 2022 года.
Бесплатный доступ
В статье представлены результаты археопаразитологического и палинологического изучения культурных слоев античного поселения Артезиан, расположенного в восточном Крыму. В рамках проведенного исследования удалось определить видовой состав паразитов и палинологический спектр культурных напластований. Распределение яиц паразитов по слоям показывает их обсемененность практически во все периоды существования цитадели. Обилие в исследуемых пробах яиц геогельминтов, которые могут принадлежать как человеку, так и домашним животным, свидетельствует о неудовлетворительном санитарном состоянии поселения и постоянном риске заражения геогельминтозами и кишечными инфекциями, имеющими одинаковый фекально-оральный путь передачи. Особенности изученных спорово-пыльцевых спектров не позволяют провести детальную реконструкцию динамики растительности, окружавшую городище Артезиан на рубеже тысячелетий. Основной причиной является сложность формирования спектров - помимо пыльцы, оседающей из воздуха (пыльцевого дождя), значительную долю составляют пыльцевые зерна, привнесенные из других источников. Интерпретация спорово-пыльцевых спектров, кроме того, осложняется и наличием большого количества микроуглей и обожженных пыльцевых зерен. Несмотря на вышеизложенные трудности изучения образцов из культурных слоев археологического памятника Артезиан, спорово-пыльцевым методом удалось определить, что обитатели городища, вероятно, применяли навоз животных в качестве топлива и что на рубеже тысячелетий вокруг поселения преобладали безлесные открытые ландшафты, покрытые степной растительностью. Немаловажным результатом исследования является демонстрация нераскрытых возможностей исследования культурных слоев археопаразитологическим и палинологическим методами и использование полученных данных в качестве источника биоархеологической информации.
Археопаразитология, гельминты, палинология, культурный слой, спорово-пыльцевые спектры, восточный крым, античность
Короткий адрес: https://sciup.org/149141707
IDR: 149141707 | УДК: 902:576.8 | DOI: 10.15688/nav.jvolsu.2022.2.12
Results of archaeoparasitological and palynological research conducted on cultural layers of the Artesian ancient settlement (Eastern Crimea)
The article presents the results of archaeoparasitological and palynological research of the cultural layers located in the ancient Artesian settlement in the Eastern Crimea. The study revealed the species composition of parasites and palynological spectrum in the cultural layers. Parasite eggs distribution over the layers indicates high content of eggs in almost all periods of the citadel’s existence. High abundance of geohelminth eggs in the studied samples, which might belong to both humans and domestic animals, indicates the unsatisfactory sanitary state of the settlement and the constant risk of infection with geohelminthiasis and intestinal infections possessing the same fecal-oral route of transmission. Features of the studied spore-pollen spectra do not allow a detailed reconstruction of the vegetation dynamics in the vicinity of the Artezian fortress at the turn of the millennium. The major reason is complexityof spectra formation: in addition to pollen deposited from the air (pollen rain), a significant proportion consists of pollen grains brought from other sources. Furthermore, interpretation of the spore-pollen spectra is complicated by presence of a large number of microcharcoals and burnt pollen grains. Despite the above-mentioned challenges in the study of samples from the cultural layers of the Artezian archaeological site using spore-pollen method, we have determined that the inhabitants of the settlement probably used animal dung as fuel, and that at the turn of the millennium, forestless open landscapes covered with steppe vegetation were predominant around the settlement. An important result of the study is the demonstration of unexplored opportunities to investigate cultural layers by archaeoparasitological and palynological methods and of the obtained data use as a source of bioarchaeological information.
Текст научной статьи Результаты археопаразитологического и палинологического исследования культурных слоев античного поселения Артезиан (Восточный Крым)
Способность оболочек яиц кишечных паразитов длительное время противостоять агрессивным воздействиям окружающей среды и возможность идентификации многих из них до рода / вида с помощью обычной световой микроскопии делает их важным источником информации о различных сторонах жизни древнего населения, таких как диета, хозяйственно-культурный тип, состояние здоровья, контакты, миграции древних популяций [Reinhard, 1992].
Яйца кишечных паразитов могут содержаться во многих образцах грунта, полученных при археологических раскопках. При этом обнаружение останков паразитов в кишечном содержимом мумифицированных останков человека, грунте с поверхности крестца и т. д. будет отражать индивидуальную зараженность человека кишечными паразитами [Slepchenko et al., 2015; 2019]. Исследование проб содержимого туалетов и канализационных стоков может показать видовой состав кишечных паразитов, характерных для группы людей [Неделькин и др., 2021; Ledger et al., 2021].
Сложности как методического, так и интерпретационного характера возникают при исследовании культурных слоев городов и многослойных поселений. С одной стороны, наличие яиц эндопаразитов может указывать на присутствие фекалий в грунте и свидетельствовать о зараженности гельминтами популяции людей [Reinhard, 1992; Campbell et al., 2011, p. 23]. С другой стороны, низкие концентрации яиц (так называемая «городская фоновая фауна») могут быть следствием пере-отложенности из ранее засыпанных выгребных ям, переноса с пылью или насекомыми и свидетельствовать лишь о присутствии человека [Reinhard, 1992; Kenward, 2009, p. 25]. Кроме того, при анализе грунта с поселения значительной проблемой является определение видовой принадлежности останков паразитов – далеко не всегда удается отличить яйца паразитов человека от паразитов животных [Kenward, 2009, p. 25]. Таким образом, часто остается не понятным, что же в точности отражает видовой состав паразитов, обнаруженный при исследовании напластований.
На ряд поставленных вопросов можно получить ответы с помощью археоботаничес-ких исследований [Kenward, 2009]. Идентификация в культурном слое индикаторов пищи человека или травоядных животных иногда позволяет различить их экскременты.
Цель исследования – дать характеристику культурным слоям археопаразитологи-ческим и палинологическим методами, определить возможности и ограничения их использования при анализе культурных напластований, попытаться реконструировать динамику растительности, окружавшей городище Артезиан в древности, определить факторы риска развития паразитозов и желудочно-кишечных заболеваний, дать характеристику санитарного состояния поселения.
Городище Артезиан. Античное городище Артезиан расположено в степной зоне Крымского Приазовья, на территории царской хоры Боспорского царства, в центре урочища Артезиан (Салын). От столицы Боспора Пан-тикапея городище отстояло на 30 км, от побережья Казантипского залива Азовского моря – на 5 км (рис. 1). Исследуется с 1986 г. [Винокуров 1998; 2012а]. На раскопах I–IV были выявлены слои и сопряженные с ними строительные остатки античного и средневекового времени. На раскопе I были вскрыты кварталы плотной античной застройки, северная часть цитадели «0» времени Митридата Евпатора Диониса и северо-восточный угол поздней цитадели; на раскопе II – южная часть кварталов цитадели «0», «казарменные» постройки времени Митридата Евпатора, уничтоженные в ходе землетрясения 63 г. до н.э., а также многочисленные хозяйственные и жилищные комплексы первых веков нашей эры. На раскопе III открыты преимущественно оборонительные сооружения и десять помещений ранней цитадели, погибшей в 46/47 г. в начальной фазе боспоро-римской войны 44/34–49 гг.; двор, внутренние помещения и башни 2–5 поздней цитадели второй половины I в. н.э. – III в. н.э. На раскопе IV обнаружены остатки хозяйственно-жилой застройки первых веков н.э. [Винокуров 2005; 2008; 2009; 2010а; 2010б; 2012а; 2012б; 2013; 2014; 2018]. Сохранность открытых античных и средневековых строительных остатков памятника чрезвычайно плохая, отдельные не потрево- женные перекопами участки незначительной площади выявлены только на дне раскопов.
Не исключено, что на месте городища располагался древний город Пароста, упомянутый римским автором Плинием и греческим географом Клавдием Птолемеем. Название Пароста в переводе с древнеиранского – «стоящая впереди», а с индоарийского – «расположенная у устья, у гавани». Действительно, этот пункт был передовым форпостом в сложнейшей системе боспорских укреплений – валов и рвов, усиленных крепостями и фортами, оберегавших жизненно важные центры Боспорского царства от кочевников, вторгавшихся в Крым со стороны Перекопа.
Городище «Артезиан» представляет собой одну из крупных крепостей Крымского Приазовья со сложной планировкой разновременных жилых кварталов и оборонительных сооружений, имеет большое значение как эталонный стратиграфический объект для изучения центральных поселений царской хоры Бос-пора первых веков нашей эры. Городище контролировало «царскую» дорогу вдоль северного склона Артезианского урочища – от Азовского моря к Пантикапею и собственно урочище – значительную по площади естественную локально-территориальную зону Боспо-ра. Важное стратегическое значение этого поселения было несомненным.
Центральное положение Артезиана обеспечивало его доминанту над другими поселениями урочища в торговом, сырьевом, ремесленном и оборонном значениях. Этот населенный пункт был удобен для сбора налогов или контроля за поселениями указанной территории, так как находился на равновеликом удалении от морского побережья и ведущих укрепленных поселений соседних территориально-хозяйственных зон. На городище были размещены представители государственной администрации и царский военный контингент. Возможно, здесь находились люди, занимавшиеся профессионально торговлей, собирающие и реализующие излишки сельскохозяйственной и ремесленной продукции населения урочища.
В прямой видимости от городища находятся высоты, на которых в древности располагались укрепленные поселения, входившие в единую оборонительную систему Боспорско-го царства (Ново-Отрадненское, Либкнехтов- ское, Багерово-Северное и др.). С цитадели городища хорошо просматриваются побережье Казантипского залива с доминирующим над местностью городищем Ново-Отрадное, устье реки Аджиэль, а также полуостров Казантип.
Городище неоднократно горело и разрушалось вследствие варварских штурмов и мощных землетрясений, но всегда восстанавливалось. В ходе исследования памятника обнаружены крепостные и жилые сооружения, уникальные артефакты, надписи, сотни граффити и дипинти, терракотовые фигурки, тысячи монет, многочисленные жилые, хозяйственные и производственные комплексы, а также реперные для боспорской истории слои пожара периода боспоро-римской войны 46/47 г. [Винокуров 2010а; 2010б; 2013; 2018; 2020].
Материалы и методы
Культурный слой городища Артезиан мощностью от 1,20 до 2,80 м (во рву «Цитадели» – до 5,80 м) разделяется на два горизонта: античный первой половины IV в. до н.э. – начала IV в. н.э. и салтовский VIII–IX веков. Более ранние культурные напластования переотложены в античный период.
Непосредственно во время проведения археологических раскопок был зачищен западный борт раскопа III с культурными напластованиями и строительными остатками ранней и поздней цитаделей городища Артезиан (античный горизонт). В борту выделено 13 слоев:
Слой 1. Переотложенная погребенная почва горизонта «В». Связана с устройством террас эпохи эллинизма – рубежа нашей эры.
Слой 2. Слой строительного отеса и желтой глины связан с первичной перепланировкой городища в эпоху эллинизма.
Слои 3–8. Нижний античный горизонт сформировался в период от 370–235/220 гг. до н.э. до начала строительства ранней цитадели.
Слои 9–10. Время строительства, бытования ранней цитадели от середины I в. до н.э. – 46/47 н.э.
Слои 11–12. Нивелировочные слои, связанные c периодом строительства поздней цитадели 49–54 гг. н.э. Образованы из переотложенных напластований периода бытования и слоя разрушения ранней цитадели.
Слой 13. Верхний нивелировочный слой под постройки поздней цитадели середины – второй половины I в. н.э.
Для археологического и спорово-пыльцевого анализа с каждого слоя со всей толщи борта в индивидуальные пакеты было отобрано 13 проб грунта (по количеству слоев) весом от 150 до 350 грамм.
Археопаразитологический анализ. В лабораторных условиях к сухой пробе, предварительно помещенной в химический стакан объемом 800 мл, добавлялся 0,5%-ный раствор тринатрий фосфата (Na3PO4). Химические стаканы с пробами накрывались фильтровальной бумагой, через одну неделю сливалась надосадочная жидкость. Осадок пропускался через сито с диаметром ячей 200 мкм. Сепарацию проб проводили в центрифужных пробирках. Осадок собирали многократным центрифугированием в течение 7 мин. (1500 оборотов в минуту). После этого к пробам добавлялся глицерин [Callen, Cameron, 1960].
При обработке промеров длины яиц нематод рода Trichuris применяли поправочный коэффициент, равный 1,1, на отсутствие полярных пробочек. Для яиц трихурисов с деградированными оболочками с целью получения сравнимых данных также применяли поправочные коэффициенты: 1,1 по длине и 1,04 по ширине для яиц с сильно деградированными оболочками и 1,05 по длине и 1,02 по ширине для умеренно деградированных яиц. Поправочные коэффициенты были высчитаны путем промеров яиц на фотографиях из публичных источников. Измерение длины яиц трематод с несохранившейся крышечкой проводили, как если бы они были с крышечкой. Явно уродливые, деформированные и поврежденные яйца для промеров не применяли.
В качестве описательных статистик для измеряемых признаков яиц использовали показатели абсолютного размаха размера (минимальный – min; максимальный – max), среднее арифметическое (mean – M) со стандартным отклонением (standard deviation – SD). Для вычислений числовых параметров и построения диаграмм применяли программу MS Excel из состава MS Office 2019.
Для определения видовой принадлежности яиц паразитов человека использовали руководства [Ash, Orihel, 2007; Гаевская, 2015], также материалы сайта CDC DPDx – Laboratory Identification of Parasites of Public Health Concern. Яйца гельминтов животных идентифицировали по следующим материалам [Foreyt, 2001; Zajac, Conboy, 2021].
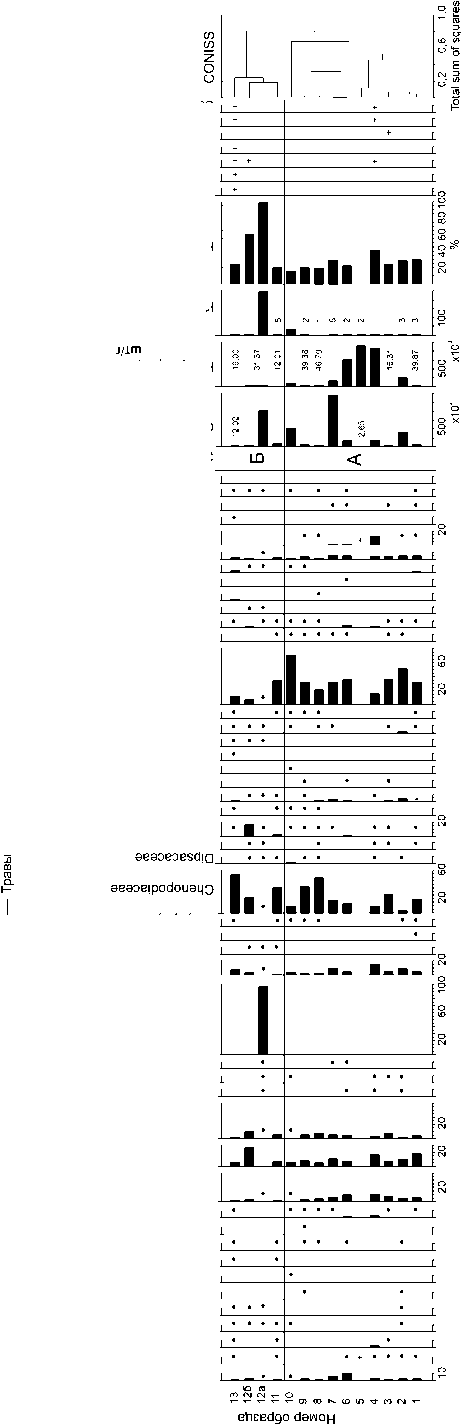
Спорово-пыльцевой анализ. Пробопод-готовка для спорово-пыльцевого анализа 13 образцов с городища Артезиан проводилась по стандартной методике с использованием тяжелой жидкости (KJ + CdJ2). Кроме спор и пыльцы высших растений учитывались и непыльцевые палиноморфы (микроугли, споры грибов, водоросли). В каждом образце подсчитано 300–500 пыльцевых зерен и не менее 500 микроуглей. При расчете доли таксонов в спектре за 100 % принята сумма пыльцы древесных и травянистых растений. Концентрация пыльцевых и непыльцевых палиноморф определялась с помощью добавленных (перед обработкой соляной кислотой – HCl) в каждый образец таблеток, содержащих споры Lycopodium clavatum (butch № 140119321). Построение диаграммы проводилось с применением программы Tilia 2.0.41 [Grimm, 2019]. Па-линозоны выделялись на основании стратиграфически ограниченного кластерного анализа (CONISS) [Grimm, 1987, pp. 13–35]. За функцию расстояния принято евклидово расстояние.
Резул ьтаты
Археопаразитологический анализ. Наиболее широко в образцах были представлены яйца нематод рода Trichuris . Яйца имели характерную удлиненную бочонковидную форму, сужающуюся к полюсам с устьями; скорлупа яиц толстая с двойным контуром. Пробочки на полюсах у всех яиц отсутствовали, большое количество яиц было с деградированной наружной оболочкой (рис. 2, a,b ). При анализе распределения линейных размеров яиц по длине и ширине были выявлены две четко разграниченные неравные по объему группы (рис. 3).
Более мелкие яйца значительно превалировали в образцах, их общее количество превышало 92 %. Частотные и размерные характеристики обеих групп яиц Trichuris приведены в таблице.
Также в образцах регулярно обнаруживались яйца трематод рода Brachylaima. Они имели характерную для брахилаймид эллипсоидную асимметричную форму и толстую, темно-коричневую оболочку с утолщением на противоположном от крышечки конце. Большинство яиц было с отпавшими крышечками, многие яйца были деформированы (рис. 2,с). Размеры яиц довольно сильно варьировали по длине от 27,2 до 40,6 (в среднем 32,9 ± 2,7) мкм и по ширине от 17,3 до 25,6 (в среднем 20,4 ± 1,5) мкм (рис. 4).
Яйца аскарид рода Ascaris в образцах были менее многочисленны и присутствовали не во всех слоях. Они имели коричневый цвет, округлую или овальную форму с толстой бугристой оболочкой и аморфным содержимым (рис 2, d ). Их размеры варьировали от 46,4 до 61,2 мкм в длину и от 35,2 до 46,6 мкм в ширину, в среднем 54,6 ± 4,1 х 41,9 ± 3,5 мкм.
Количество и соотношение яиц гельминтов разных родов в исследованных слоях было очень неравномерным (рис. 5).
Дифференциальная диагностика яиц паразитов. Яйца различных видов нематод рода Trichuris имеют хорошо узнаваемую, но очень сходную между собой морфологию. Благодаря гостальной специфичности их идентификация может быть проведена по виду хозяина, а также по размерам, которые, однако, у разных видов могут значительно перекрываться.
Учитывая археологический контекст материала и размерные характеристики яйца Trichuris sp. I типа могут быть отнесены как к человеческому ( T. trichiura) , так и свиному ( T. suis) видам власоглавов. Оба этих вида практически неразличимы морфологически. Общепринятые референтные значения размеров T. trichiura составляют 49-65 х 2029 мкм, в среднем 54 мкм х 22 мкм [DPDx, 2016], хотя некоторые авторы указывают на более широкий размах размеров, в частности от 47, 45 и даже 43 мкм в длину [Meng et al., 1986, p. 20; Yoshikawa et al., 1989; Areekul et al., 2010; Jaeger et al., 2016]. Примечательно, что в целом (по средним значениям) яйца в нашем материале оказались несколько мельче по длине и одновременно шире по сравнению с референтными значениями. Этот факт, вероятно, связан с тафономическими изменениями яиц, при которых деградация оболочки сопровождается также изменением их формы.
Яйца Trichuris sp. II типа по мерным и морфологическим признакам соответствуют T. muris . Окончательными хозяевами власоглава этого вида являются синантропные грызуны (мыши, крысы).
Яйца брахилаймидных трематод, учитывая географическое положение места исследований и возможный круг хозяев, могут быть отнесены к роду Brachylaima . Широкий размах в размерах, а также вариабельность деталей морфологии заставляют предположить наличие в материале яиц нескольких видов брахилайм. Среди них наиболее вероятны виды, паразитирующие в синантропных грызунах ( B. recurva и др.) и свиньях ( B. suis ).
Обнаруженные яйца аскарид могут принадлежать аскаридам человека ( A. lumbricoides ) или свиней ( A. suum ). Их дифференциация до вида практически невозможна ввиду чрезвычайного морфологического и морфометрического сходства [Ash, Orihel, 2007].
Спорово-пыльцевой анализ. Доля пыльцы древесных пород в изученных образцах не превышает 11 %, хотя и представлена десятью таксонами: сосной ( Pinus ), ольхой ( Alnus ), грабинником / березой ( Carpinus orientalis / Betula ), грабом обыкновенным ( Carpinus betulus ), лещиной ( Corylus ), буком ( Fagus ) ивой ( Salix ), липой ( Tilia ), дубом ( Quercus ) и вязом ( Ulmus ) (рис. 6). Кроме сосны пыльца всех пород встречается единично (менее 1,5 %). Доля пыльцы Pinus варьирует от 1 до 5 % и лишь в слое № 6 достигает 10 %.
Основу (46–85 %) спорово-пыльцевых спектров составляют злаки (Poaceae), маревые (Chenopodiaceae) и астровые (Asteroideae и Cichorioideae). Исключение составляет образец из слоя № 12, спектр которого можно охарактеризовать как монодоминантный, то есть основная часть подсчитанных пыльцевых зерен (97 %) приходится на один таксон (Echium/Onosma, Boraginaceae). Для понимания того, как распределены доли остальных таксонов, на диаграмме представлен и спектр этого образца без Echium/Onosma (№ 12б). В семи из тринадцати образцов доминирует пыльца злаков, в трех (№ 8, 9 и 13) – маревые, в других двух (№ 4, 12) – астровые. В слое № 5 концентрация пыльцы очень низкая (265 шт/г), что не позволило рассчитать доли пыльцы таксонов в этом спектре; на диаграмме присутствие встреченной в этом образце пыльцы отмечено «+». Несмотря на довольно большое таксономическое разнообразие (45 таксонов), пыльца большей части таксонов в образцах представлена единично и составляет 1 % и менее (рис. 6). Практически во всех образцах присутствуют пыльцевые зерна культурных злаков (Cerealia t.) (рис. 7,a).
На диаграмме выделены две палинозо-ны (рис. 6). Кластерный анализ проводился без образца № 12а. Зоны имеют незначительные отличия. Можно лишь отметить, что в спорово-пыльцевых спектрах палинозоны А, в отличие от Б, меньшее разнообразие древесных пород, чуть больше пыльцы сосны ( Pinus ), полыни ( Artemisia ), капустных (Brassicaceae) и злаков (Poaceae). Доля последних в зоне А варьирует от 22 до 70 %, лишь в образце № 4 снижаясь до 16 %, а в зоне Б – от 8 до 35 %.
Общая концентрация пыльцы и спор невысокая, максимальных значений достигает в образцах № 7 и 12 (146 900 и 102 600 шт/г соответственно). В большинстве образцов концентрация низкая и не превышает 20 000 шт/г.
Все образцы насыщенны микроуглем, хотя визуально присутствие золы отмечено только в слоях № 4, 5, 6, 8, 10 и 11. Концентрация микроуглей значительно варьирует в образцах – от 12 000 до более двух миллионов шт/г в зольных слоях (рис. 6). В четырех образцах присутствуют споры копрофиль-ных грибов. Среди непыльцевых палиноморф в образце № 13 также отмечены ценобии водоросли педиаструма ( Pediastrum ). В связи с низкой концентрацией спор грибов и цено-бий на диаграмме отмечено лишь их присутствие (рис. 6).
Кроме спор, пыльцы и непыльцевых палиноморф в образцах подсчитывалось количество комков пыльцы, встреченных при учете пыльцевых зерен и спор. Бóльшая часть комков представляет собой группы недозревших не разлетевшихся пыльцевых зерен одного таксона. В двух образцах (№ 5, 10) встречены большие фрагменты пыльников и даже целые пыльники (рис. 7, b,с ), а также комки пыльцы, состоящие из пыльцевых зерен разных таксонов (№ 11, 12, 13), наибольшее их количество в образце № 12 (рис. 6).
Обсуждение
Археопаразитологический анализ . Осадконакопление, приводящее к формированию культурных слоев на археологических памятниках, происходит неравномерно и зависит от воздействия ряда природных и антропогенных факторов [Karkanas, Goldberg, 2019, pp. 21–148]. При обнаружении в образцах грунта яиц гельминтов не всегда удается точно определить их принадлежность паразитам человека и/или животных. Ситуация осложняется еще и тем что яйца некоторых гельминтов человека и животных практически не имеют морфологических и морфометрических отличий. Таким образом, контекст отбора образцов и наличие сравнительного материала с других памятников очень важен для проведения археопаразитологических реконструкций.
Археопаразитологические исследования на территории Северного Причерноморья пока немногочисленны. В то же время по видовому составу паразитов результаты исследования культурных слоев поселения Артезиан во многом сходны с полученными из других памятников, расположенных на территории Северного Причерноморья.
Так яйца аскарид, власоглавов и бра-хиляймид в достаточном количестве были обнаружены при анализе проб грунта из водотоков хазарского периода города Фанагория VIII–IX вв. н.э. и в пробах содержимого канализационного стока Чоргунской башни расположенной в Юго-западном Крыму и датированной XV в. н.э. [Недель-кин и др., 2021].
Однако соотношение яиц указанных выше видов паразитов в изученных образцах с этих археологических памятников несколько разнится. При исследовании образцов из водостока Фанагории выявлено минимальное число мышиных / крысиных Trichuris muris в сравнении со значительным количеством яиц человеческого / свиного власоглавов и яиц аскарид. При этом в содержимом канализационного стока Чоргунского донжона выявлено обратное соотношение – несколько большее количество яиц человеческого / свиного и мышиных / крысиных власоглавов, при минимальном количестве яиц аскарид.
Интерпретация подобного соотношения яиц паразитов может указывать на то, что в водоток средневековой Фанагории попадали в основном яйца власоглавов и аскарид человека и/или свиней при минимальном участии синантропных грызунов, в то время как отложения канализационного стока сформировались при участии только человека и грызунов (крыс и мышей).
Яйца брахиляймид, обнаруженные в образцах из этих памятников, отличались размерами. Более крупные яйца, обнаруженные в образцах из Фанагории, вероятно, принадлежали паразитам свиней, в то время как бра-хиляймидные яйца из проб канализации Чор-гунской башни были от мышиных / крысиных трематод.
Ввиду близости видового состава паразитов и их соотношения в образцах культурных напластований цитадели поселения Артезиан к пробам грунта из водотока средневековой Фанагории можно предположить и схожесть формирования культурных напластований. Вероятно, их генез проходил в условиях относительно открытых пространств с участием в большей степени человека и, возможно, свиней и минимальном вкладе грызунов.
Спорово-пыльцевой анализ. Динамику тех или иных таксонов в спорово-пыльцевых спектрах культурных слоев археологических памятников часто связывают с изменениями в растительном покрове или даже с динамикой климатических условий. Однако подобного рода интерпретации не всегда могут быть корректными. Состав спорово-пыльцевых спектров культурных слоев сильно зависит от типа источника, откуда поступают пыльца и споры. В случае преобладания пыльцевого дождя, то есть пыльцы, переносимой от цветущих растений по воздуху, образцы из культурных слоев в действительности могут отражать изменения окружающей растительности подобно естественным разрезам (болотным и озерным отложениям, например). В случае же при-вноса пыльцы человеком (строительство, заготовка пищи растительного происхождения, лекарственных трав и т. п.) или животными (экскременты, содержащие пыльцу съеденных цветущих растений) спорово-пыльцевые спектры в большей степени отражают хозяйствен- ную деятельность человека или состав пищи животного нежели фитоценоз окружающий памятник [Бабенко, Сергеев, 2019; Малышев и др., 2021; Сергеев и др., 2022]. Для понимания присутствия того или иного источника пыльцы при изучении археологических памятников крайне необходимо знать, откуда отобраны образцы: открытые контексты, где накапливался культурный слой без нарушений; открытые или закрытые объекты (хозяйственные ямы, полы построек, отхожие места, колодцы и др.), строительные слои или слои, сформировавшиеся в результате разрушения стен конструкций [Deforce 2017; Brown et al., 2017; van Amerongen, 2020; Але-шинская и др., 2021].
Присутствие в спорово-пыльцевых спектрах изученных образцов комков пыльцы и большой доли насекомоопыляемых растений (рис. 6) может свидетельствовать о наличии в слоях, откуда отобраны образцы, экскрементов травоядных животных [Carrion, 2002; Бабенко и др., 2007; Florenzano, 2019]. Комки пыльцы, или даже целые пыльники, в навозе указывают на поедание животными растений в период цветения и/или бутонизации. Нередко спорово-пыльцевые спектры такого навоза являются монодоминантными [Babenko et al., 2015; Бабенко и др., 2021].
Наиболее ярко рассмотренные признаки навоза выражены в образце № 12 – огромное количество комков пыльцы и абсолютное доминирование пыльцы растений – представителей семейства бурачниковые (Boraginaceae) (рис. 7,с,d). К сожалению, возможности спорово-пыльцевого анализа ограничены, и невозможно определить, пыльца конкретно какого вида преобладает. Учитывая современное распространение видов Boraginaceae на Керченском полуострове [Рубцов, 1972, с. 381– 393], морфологию оболочки и размеры их пыльцевых зерен [Beug, 2004, с. 160–161, с. 338–339], можно предположить, что наиболее вероятными представителями семейства являются синяк (Echium) и оносма (Onosma) (рис. 7,e). Представители этих родов неохотно, но могут поедаться травоядными [Орлова, 1990, с. 116–117, с. 125–127; Шабанова и др., 2012, с. 120–121]. В этом же образце, но в спектре без участия пыльцы Echium/Onosma (№ 12б), можно отметить увеличенную, по сравнению с другими образцами, долю пыльцы бобовых Fabaceae (16,6 %). Высокий процент присутствия этой пыльцы в спектре легко объяснить наличием комков пыльцы Fabaceae. Этими же причинами (наличие комков пыльцы) можно объяснить и высокую долю злаков в слое № 10, астровых в слоях № 4 и 12, капустных в слое № 4 и маревых (слой № 13) в изученных образцах (рис. 7,f,g,h,i). Так, в рассмотренных образцах большая доля таксонов, вероятнее всего, не свидетельствует об изменениях в растительном покрове, а на самом деле отражает присутствие экскрементов травоядных животных в образцах.
Наличие в спектрах комков пыльцы, состоящих из пыльцевых зерен не только одного, но и разных таксонов, также может являться свидетельством поступления навоза в культурный слой [Florenzano, 2019]. В трех из 13 изученных образцов (№ 11, 12, 13) присутствуют комки такого рода (рис. 7, j ).
Еще одним доказательством того, что культурные слои городища содержат экскременты травоядных животных, являются споры копрофильных грибов: Chaetomium t., Coniochaeta t., Gelasinospora t., Podospora t., Sordaria t., Sporormiella t. (рис. 8, a , b , c ) [Lee et al., 2022]. Небольшое количество аскоспор в образцах, возможно, связано с климатическими условиями (недостаточная влажность [Lee et al., 2022]) и/или отсутствие периода времени, необходимого для развития грибов (2–4 недели [Lee et al., 2022]). Наибольшим количеством и разнообразием аскоспор характеризуется образец № 13. В нем же обнаружена и зеленая водоросль ( Pediastrum ) (рис. 8, d ), обитающая в пресных водоемах (озеро, пруд или болото) [Komarek, Jankovska, 2001, p. 16–19] и единично присутствует пыльца рогоза ( Typha ). Отличия этого образца по составу непыльцевых палиноморф, вероятно, связано с особенностями формирования культурного слоя. Можно предположить, что слой, описанный как верхний нивелировочный, какое-то время был в доступе для посещения травоядными животными, в экскрементах которых успели развиться копрофильные грибы. Наличие ценобий педиаструма в таком случае указывает на то, что у животных в доступе для питья был пресный водоем (пруд или озеро).
Другое возможное объяснение – присутствие в слое сырца, для производства которого использовали несвежий навоз и воду и/или ил / глину из пресного водоема [Сергеев, Лебедева, 2018; Малышев и др., 2021].
Еще одной отличительной чертой (кроме наличия комков пыльцы) изученных образцов с городища Артезиан является присутствие большого количества микроугля. Следует отметить, что один из источников микроуглей на памятнике – жженный навоз, о чем свидетельствует обгоревшие пыльцевые зерна и комки пыльцы (рис. 7, g , h , i , рис. 8, e , f , g , h , i ). Пыльца под воздействием высоких температур сгорает, а при 200 °С теряет прозрачность [Brooks, Shaw, 1972, pp. 69–87], что затрудняет ее определение и увеличивает долю неопределенных пыльцевых зерен (рис. 8, f ). Особенно ярко это проявилось в образцах № 4 и 5, где концентрация микроуглей достигает максимальных значений. Доля неопределенной обгоревшей пыльцы в первом образце достигает 12,8 % (рис. 6), а во втором – пыльца практически отсутствует. Найденные в образце из слоя 5 обгоревшие пыльцевые зерна и пыльники (рис. 8, e ) указывают на то, что очень низкая концентрация пыльцы в этом образце является следствием именно выгорания при высоких температурах. Систематическую принадлежность пыльцевых зерен некоторых таксонов все же можно установить даже в обожженном состоянии (рис. 8, g , h , i ). Разная концентрация микроуглей в образцах и, видимо, разная степень выгорания органики затрудняют анализ общей концентрации пыльцы и спор.
Присутствие комков пыльцы, копрофиль-ных спор, обожженных пыльцевых зерен и большого количества микроугля в образцах может указать на использование на памятнике навоза в качестве топлива при нехватке древесины.
Спорово-пыльцевые спектры изученных образцов с городища Артезиан (по указанным выше причинам), к сожалению, не дают возможности проследить динамику окружающей растительности. Можно лишь предполагать о преобладании в окрестностях памятника открытых ландшафтов, покрытых разнотравнозлаковыми степными сообществами. Значительная доля пыльцы представителей маревых
(Chenopodiaceae) и присутствие пыльцевых зерен семейства свинчатковые (Plumbaginaceae) в спектрах могут указывать на засушливые климатические условия в регионе на рубеже тысячелетий. Высказанное предположение не противоречит как современным условиям Ленинского района, так и результатам спорово-пыльцевого анализа донных отложений Азовского моря [Cordova et al. 2011; Матишов и др., 2018; Дюжова, 2019].
Заключение
Распределение яиц паразитов по слоям показывает их обсемененность практически во все периоды существования цитадели. Наибольшее видовое разнообразие приходится на античный горизонт за счет постоянного присутствия в образцах яиц аскарид. В период существования ранней цитадели яйца последнего гельминта попадаются только в одном слое, в минимальном количестве. В период существования поздней цитадели, яиц аскарид не выявлено.
Переотложение грунта во время проведения перестроек на территории цитадели, на наш взгляд, не позволяет однозначно судить о сходстве / различии видового состава паразитов в культурных напластованиях поселения Артезиан. Можно лишь сказать, что загрязненность почвы яйцами кишечных паразитов присутствовали практически во все периоды существования цитадели.
Таким образом, обилие в исследуемых пробах яиц геогельминтов, которые могут принадлежать как человеку, так и домашним животным, свидетельствует о неудовлетворительном санитарном состоянии поселения и постоянном риске заражения геогельминтоза-ми и кишечными инфекциями, имеющими одинаковый, фекально-оральный путь передачи.
Особенности изученных спорово-пыльцевых спектров не позволяют провести детальную реконструкцию динамики растительности, окружавшую городище Артезиан на рубеже тысячелетий. Основной причиной является сложность формирования спектров – помимо пыльцы, оседающей из воздуха (пыльцевого дождя), значительную долю составляют пыльцевые зерна из других источников. Сохранившиеся пыльники, комки пыльцы и споры копрофильных грибов указывают на присутствие в культурных слоях экскрементов животных, что согласуется с результатами археопаразитологического анализа. Однако ни по таксономическому составу, ни по соотношению различных таксонов невозможно установить, фекалии каких именно животных попали в культурные слои памятника. Единственное, можно отметить, что если в грунт и поступали экскременты человека, то, вероятнее всего, в небольших количествах. Интерпретация спорово-пыльцевых спектров, кроме того, осложняется и наличием большого количества микроуглей и обожженных пыльцевых зерен. Последнее приводит к нецелесообразности анализа динамики концентрации пыльцы в профиле и увеличению количества неопределенных пыльцевых зерен.
Несмотря на вышеизложенные трудности изучения образцов из культурных слоев археологического памятника Артезиан спорово-пыльцевым методом, можно сформулировать два основных вывода: 1) обитатели городища, возможно, использовали навоз животных в качестве топлива; 2) на рубеже тысячелетий вокруг поселения преобладали безлесные открытые ландшафты, покрытые степной растительностью.
ПРИМЕЧА НИЕ
1 Исследование выполнено при поддержке Российского научного фонда, грант № 21-18-00263 «Археопаразитологические спектры древнего и средневекового населения Северного Причерноморья и юга Русской равнины: новый подход в биоар-хеологических реконструкциях».
1 Research supported by the Russian Science Fundation, project no. 21-18-00263 “Archaeoparasitological spectra of the ancient and medieval population of the Northern Black Sea region and the south of the Russian Plain: a new approach in bioarchaeological reconstructions”.
ПРИЛОЖЕНИЯ
Таблица . Размеры и встречаемость двух типов яиц Trichuris
Table . Size and frequency of two types of Trichuris eggs
|
Параметры |
Trichuris sp. I |
Trichuris sp. II |
|
|
Размеры, среднее ± SD (размах) |
длина |
51,8 ± 3,1 (45,5 – 59,9) |
64,9 ± 3,1 (61,3 – 67,2) |
|
ширина |
25,1 ± 1,8 (19,3 – 29,9) |
33,5 ± 1,8 (29,6 – 35,9) |
|
|
Доля ( n = 285, 100 %) |
92,3 % |
7,7 % |
|
Примечание . Размеры приведены с поправкой на отсутствующие пробочки и деградированную наружную оболочку.
Note. Dimensions are adjusted for missing plugs and degraded outer shell.
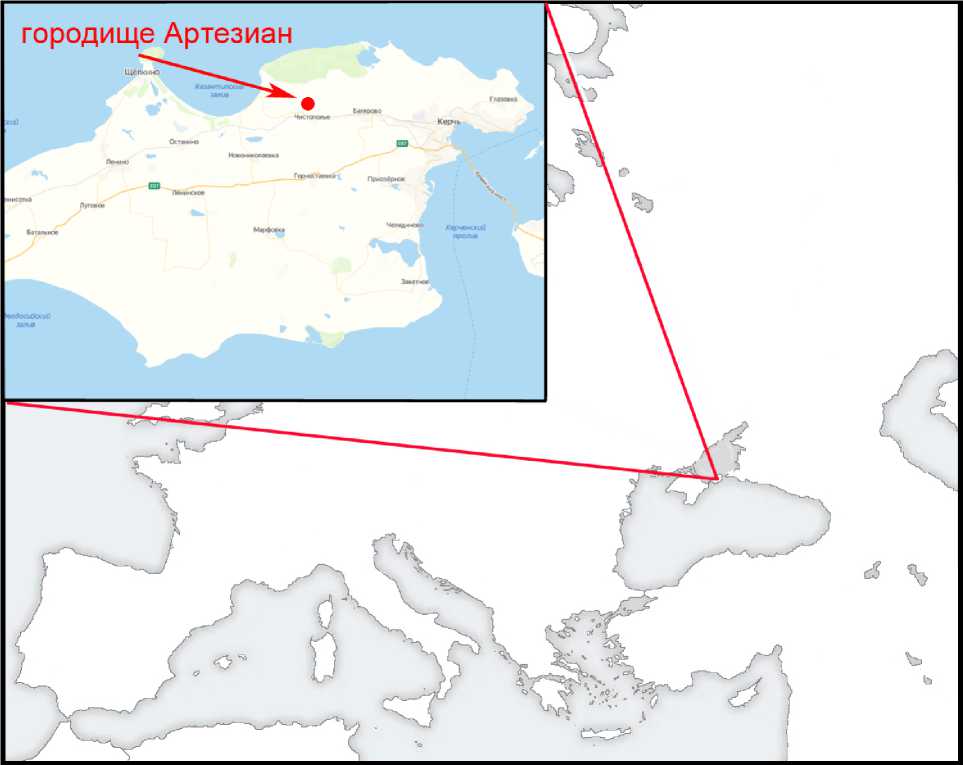
Рис. 1. Расположение городища Артезиан на полуострове Крым
Fig. 1. Location of the ancient Artesian settlement on the Crimean Peninsula
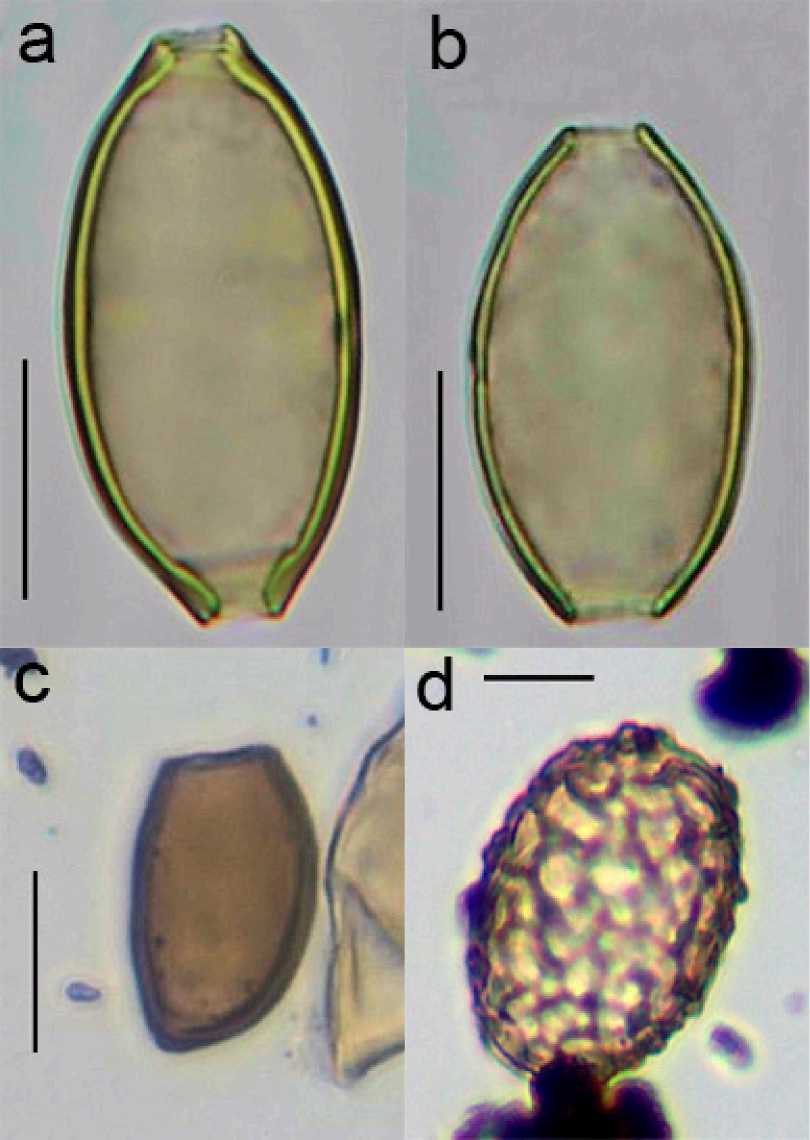
Рис. 2:
a – яйцо Trichuris с сохранившейся оболочкой; b – яйцо Trichuris c деградированной оболочкой; с – яйцо Brachylaima ; d – яйцо Ascaris
Fig. 2:
a – Trichuris egg with a preserved shell; b – Trichuris egg with a deteriorated shell;
c – Brachylaima egg; d – Ascaris egg
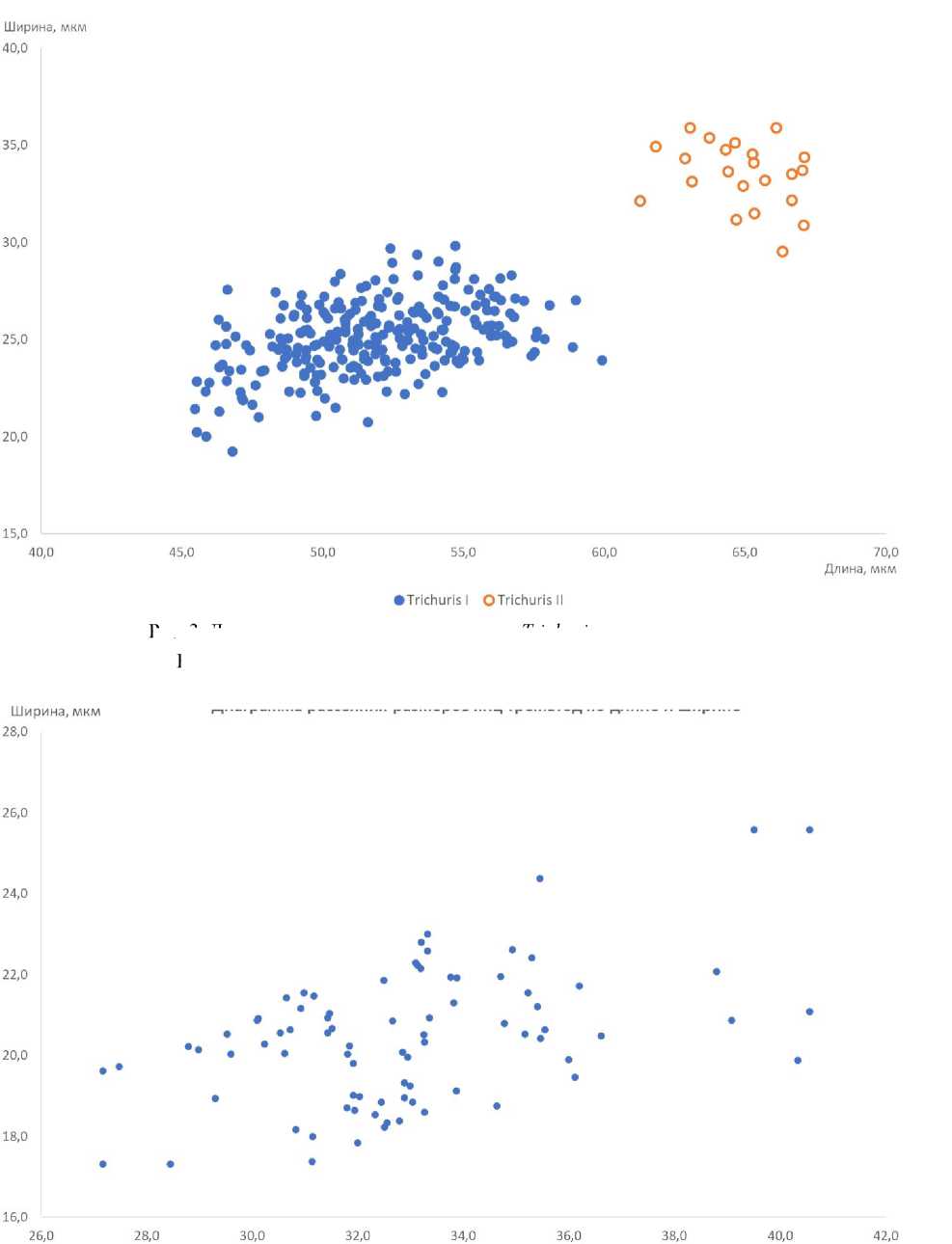
Длина, мкм
Рис. 4. Диаграмма рассеяния размеров яиц трематод по длине и ширине
Fig. 4. Scatter diagram of trematode eggs size by length and width
Рис. 3. Диаграмма рассеяния размеров яиц Trichuris по длине и ширине Fig. 3. Scatter diagram of the Trichuris eggs size by length and width
Диаграмма рассеяния размеров яиц трематод по длине и ширине
Количество яиц гельминтов разных родов по слоям
Количество яиц
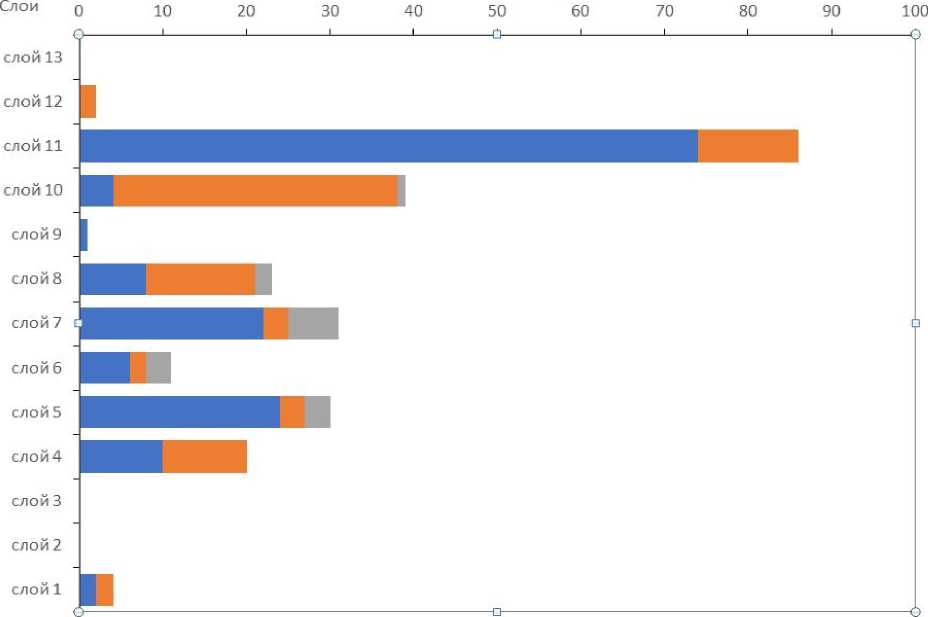
■ Trichuris eBrachylaima eAscaris
Рис. 5. Количество яиц гельминтов разных родов по слоям
Fig. 5. Layer distribution of helminth eggs of different genera
im ‘яЬяшяи mxiaio»
эпизод
1ЯНОЭХЕ1
91Я1Л19 KillЯ и О 01Л1 ОХ 90 В |-|
9E90E0d j/im ‘douo и 1яГ1яшяи KHhBdiHahHOx BBhigo
вшзоио/шпщоз gEgpioi-ioqoig aBepiojaisv
В181ШЭру
9BaoBUi6BJogj q iiinpjodouoj q BajnB)uaoj
9B9OB||AqdoAjB9 j
OBOOBQBUUBOj эвээв|nuвdшвэJ
OBaoBoissBjgj
q BUBpjQSj q B||o^jojodsj q Bjodsopodj q BJodsouiSB|99j 1 BjgBqooiuoQj 1 шn!шo^эвцэJ LunjisEipgdj иэшАос1>1И1/\1 ь5иЬвс11нэЬно>|
1ЯНО£
9)9|IJ) 9JOds3 9B9OEipodA|Odj
" S9|BAjg I
______BqdAu
BEmagdojgo t5BHHU9V9duo9|-|J KEHtfeodog x-gj 9E9OEIJE|nqdOJOSj 1 хэшпу J 9E90BS0dj 9E90E|n0UnUEdj 9E90EU06X|0d j ) B!|E9J9Qj
ЭEЭЭEU!6Eqшn|dJ o6B|UB|dj 9B90EJ9AEdEd J 9B90BJ60U0j 9E90EA|E|/\|j 9B90B!||-|j ЭB9ЭE!ШB-|J 9B9OB!UBJ90j 9B90EqEJJ Bjpgqdgj
9B90BldVj snш|nJ snojanQj e!l!lj xiiesj snBEjj sniAjoQj sn|n|gq snuidjBQj Е|щэд/5!|в}иэио snuidjBQj snuivj snuidj
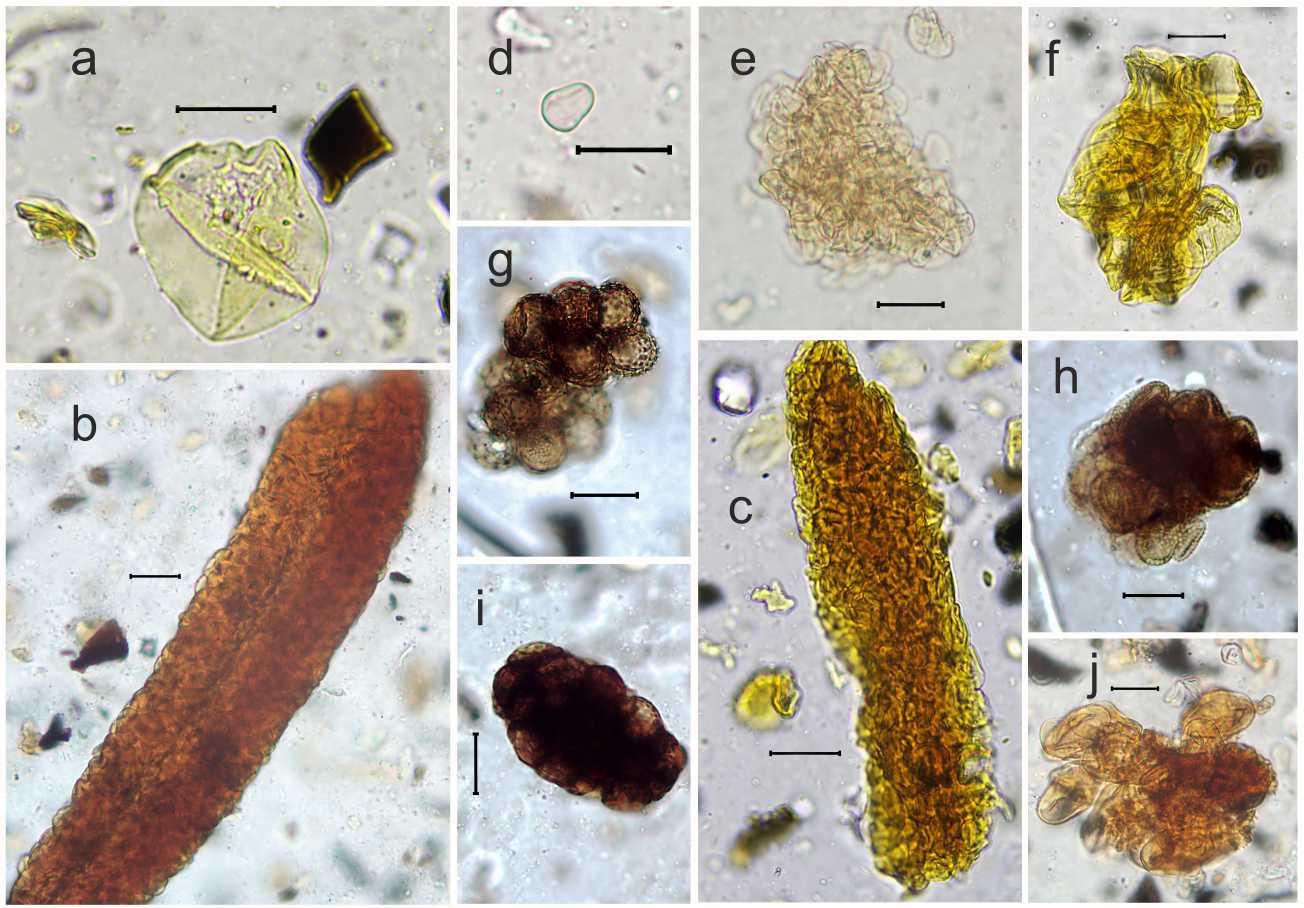
Рис. 7. Пыльцевые зерна некоторых травянистых растений и комки пыльцы (масштабная линейка = 20 мкм): a – Cerealia t. ; b – пыльник; c – фрагмент пыльника; d – Echium/Onosma ; e – комок пыльцы Echium/Onosma ; f – комок пыльцы Poaceae ; g , h , i – обожженные комки Asteroideae , Brassicaceae , Chenopodiaceae ; j – комок пыльцы разных таксонов
Fig. 7. Pollen grains of some herbaceous plants and pollen clumps (scale bar = 20 µm):
a – Cerealia t. ; b – anther; c – anther fragment; d – Echium/Onosma ; e – Echium/Onosma clump; f – Poaceae clump; g , h , i – Asteroideae , Brassicaceae , Chenopodiaceae burnt clumps; j – a clump of different pollen taxa
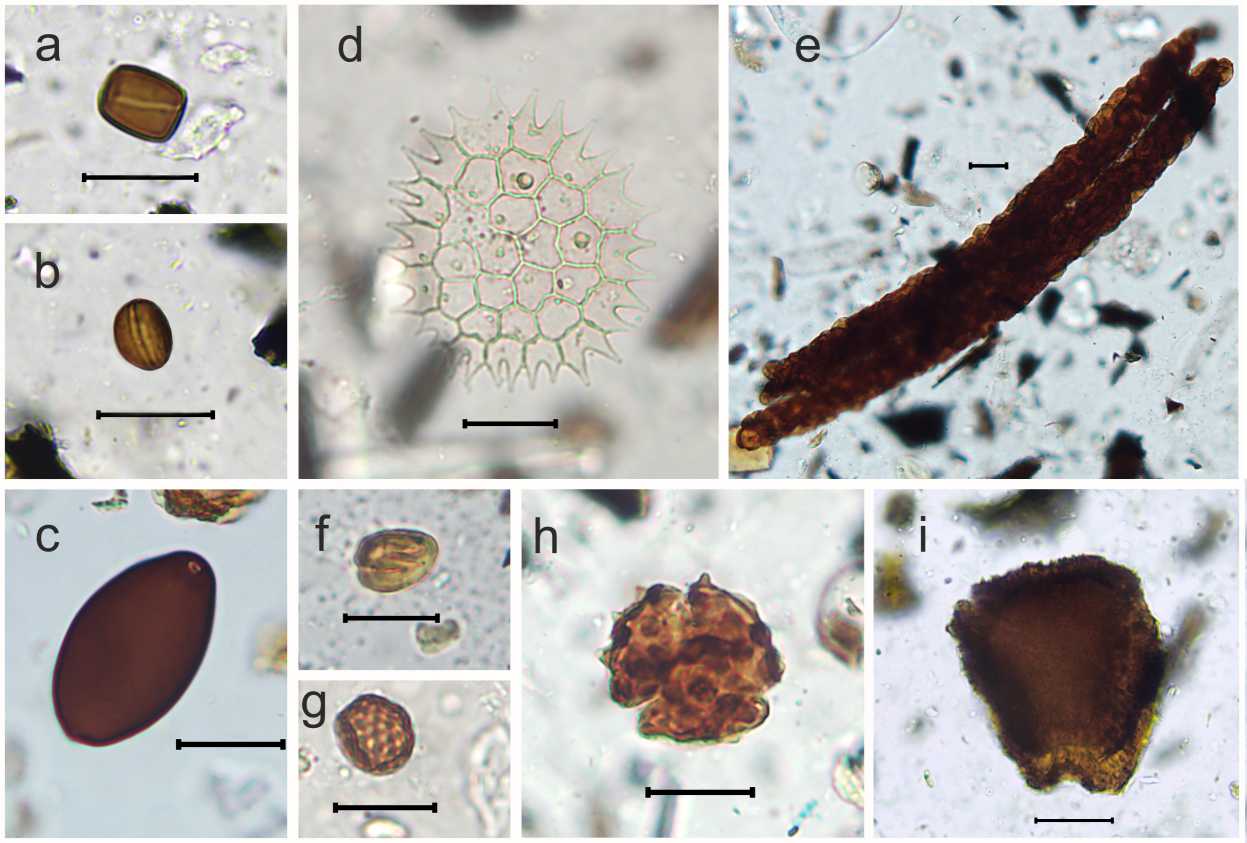
Рис. 8. Непыльцевые палиноморфы и обожженные пыльцевые зерна (масштабная линейка = 20 мкм): a – Sporormiella t .; b – Coniochaeta t. ; c – Podospora t .; d – Pediastrum ;
e – обгоревший пыльник; f – обожженное неопределимое пыльцевое зерно;
g , h , i – обожженные пыльцевые зерна Chenopodiaceae , Asteroideae , Dipsacaceae
Fig. 8. Non-pollen palynomorphs and burnt pollen grains (scale bar = 20 µm): a – Sporormiella t .; b – Coniochaeta t. ; c – Podospora t .; d – Pediastrum ; e – burnt anther; f – burnt unidentifiable pollen grain;
g , h , i – Chenopodiaceae , Asteroideae , Dipsacaceae burnt pollen grains
Список литературы Результаты археопаразитологического и палинологического исследования культурных слоев античного поселения Артезиан (Восточный Крым)
- Алешинская А. С., Бабенко А. Н., Кочанова М. Д., Трошина А. А., 2021. Возможности и особенности применения палинологического анализа на археологических объектах разного типа // Stratum Plus. № 6. С. 361–376. DOI: https://doi.org/10.55086/sp216361376
- Бабенко А. Н., Албегова З. Х., Успенский П. С., 2021. Палинологические исследования артефактов из катакомб № 97 и 98 Даргавского могильника: анализ и интерпретация // Краткие сообщения Института археологии. Вып. 263. С. 91–104. DOI: https://doi.org/10.25681/IARAS.0130-2620.263.91-104
- Бабенко А. Н., Киселева Н. К., Плахт И., Розен С., Савинецкий А. Б., Хасанов Б. Ф., 2007. Реконструкция растительного покрова центральной части пустыни Негев (Израиль) в голоцене по данным пыльцевого анализа зоогенного отложения Ацмаут // Экология. № 6. С. 417–426.
- Бабенко А. Н., Сергеев А. Ю., 2019. Археоботанические исследования городища Маджары // Поволжская археология. № 4 (30). С. 161–170. DOI: https://doi.org/10.24852/pa2019.4.30.161.170
- Винокуров Н. И., 1998. Археологические памятники урочища Артезиан в Крымском Приазовье. М.: Акад. наук СССР. Ин-т археологии. 152 с.
- Винокуров Н. И., 2005. Гибель ранней «Цитадели» городища Артезиан // Периоды дестабилизации и катасроф. Боспорские чтения. Боспор Киммерийский и варварский мир в период античности и средневековья: материалы VI Междунар. конф. Керчь: ИП Литвиненко Е.А. С. 50–60.
- Винокуров Н. И., 2008. Война, пожар или природная катастрофа: археологические критерии оценок последствий катастроф на памятниках античной археологии в Крымском Приазовье // Militaria. Боспорские чтения. Боспор Киммерийский и варварский мир в период античности и средневековья: материалы IX Междунар. конф. Керчь: ИП Литвиненко Е.А. С. 67–77.
- Винокуров Н. И., 2009. Боспоро-римская война 44/45–49 гг. и первая находка гладиуса в Крымском Приазовье // Para Bellum. № 31. С. 9–16.
- Винокуров Н. И., 2010а. Боспоро-римская война 44/45–49 гг. и гибель ранней цитадели городища Артезиан (по материалам раскопок 2004–2008 гг.) // ΣΥΜΒΟΛΑ. Античный мир Северного Причерноморья. Новейшие открытия и находки. Вып. I. М. ; К.: Ин-т археологии РАН. С. 38–44.
- Винокуров Н. И., 2010б. Новые находки времени начала боспоро-римской войны на городище Артезиан в Крымском Приазовье в 2009 году // Древности Боспора. Т. 14. М.: Ин-т археологии РАН. С. 46–65.
- Винокуров Н. И., 2012а. Археологические памятники в Крымском Приазовье (по материалам ААЭ 1988–2011). Тюбинген: LAP LAMBERT Academic Publishing. 632 с.
- Винокуров Н. И., 2012б. Хозяйственно-жилые кварталы городища Артезиан первой половины первого века н.э. // Древности Боспора. Т. 16. С. 51–76.
- Винокуров Н. И., 2013. Городище Артезиан во второй половине I в. до н.э. – первой половине I в. н.э. // Российский научный журнал. № 1 (32). С. 30–40.
- Винокуров Н. И., 2014. Раскопки помещения 10 (2009, 2011, 2013 гг.) ранней цитадели городища Артезиан // Таврические студии. Исторические науки. № 6. С. 98–105.
- Винокуров Н. И., 2018. Два слоя пожара боспоро-римской войны 44/45–49 гг. на городище Артезиан в Восточном Крыму // Древности Боспора. Т. 22. С. 56–72.
- Винокуров Н. И., 2020. Исследование пространства между крепостными стенами и рвом ранней цитадели городища Артезиан: первые результаты раскопок 2019 года // Таврические студии. № 22. С. 31–46.
- Гаевская А. В., 2015. Мир паразитов человека. I. Трематоды и трематодозы пищевого происхождения. Севастополь: ЭКОСИ-Гидрофизика. 410 с.
- Гаевская А. В., 2017. Мир паразитов человека. III. Цестоды и цестодозы пищевого происхождения. Севастополь: ЭКОСИ-Гидрофизика. 358 с.
- Дюжова К. В., 2019. Ландшафтно-климатические изменения в Приазовье в среднем и позднем голоцене (по данным спорово-пыльцевого анализа) // Матишов Г. Г. Палеогеография Приазовья в голоцене. Ростов н/Д: Изд-во ЮНЦ РАН. С. 112–124.
- Малышев А. А., Бабенко А. Н., Спиридонова Е. А., 2021. Городище Гечепсин: опыт комплексных исследований // Краткие сообщения Института археологии. Вып. 26. С. 407–421. DOI: https://doi.org/10.25681/IARAS.0130-2620.264.407-421
- Матишов Г. Г., Дюжова К. В., Новенко Е. Ю., 2018. Изменение ландшафтно-климатических условий Приазовья в среднем и позднем голоцене // Известия Российской академии наук. Серия географическая. № 3. С. 67–78. DOI: https://doi.org/10.7868/S2587556618030081
- Неделькин Е. В., Хрусталев А. В., Бабенко А. Н., Слепченко С. М., 2021. Результаты археопаразитологического и палинологического анализов содержимого канализационного стока Чоргунской башни (Юго-Западный Крым) // Stratum Plus. № 6. С. 377–389. DOI: https://doi.org/10.55086/sp216377389
- Пыльцевой анализ, 1950. М.: Госгеолиздат. 572 с.
- Растительные ресурсы СССР. Цветковые растения, их химический состав, использование: Семейства Caprifoliaceae – Plantaginaceae, 1990 / сост. Т. А. Орлова [и др.] ; отв. ред. П. Д. Соколов ; АН СССР, Ботан. ин-т им. В.Л. Комарова. Л.: Наука. 325 с.
- Рубцов Н. И., 1972. Определитель высших растений Крыма. Л.: Наука. 550 с.
- Сергеев А. Ю., Бабенко А. Н., Куприянов Д. А., Коробов Д. С., 2022. Археологический навоз как источник в системе питания сельскохозяйственных животных на Северном Кавказе во II–IV вв. н.э. (по материалам Киевского городища) // Российская археология. № 4. С. 64–77.
- Сергеев А. Ю., Лебедева Е. Ю., 2018. Сырец как источник растительных макроостатков в культурном слое археологических памятников // Археология и естественные науки в изучении культурного слоя объектов археологического наследия: материалы Междисциплинар. науч. конф. М.: КМК. С. 148–154.
- Шабанова Г. А., Изверская Т. Д., Гендов В. С., 2012. Дикорастущие хозяйственно-ценные растения заповедника «Ягорлык». Кишинев: Eco-TIRAS. 264 с.
- Areekul P., Putaporntip C., Pattanawong U., Sitthicharoenchai P., Jongwutiwes S., 2010. Trichuris vulpis and T. trichiura infections Among Schoolchildren of a Rural Community in Northwestern Thailand: The Possible Role of Dogs in Disease Transmission // Asian Biomedicine. Vol. 4 (1). P. 49–60. DOI: https://doi.org/10.2478/abm-2010-0006
- Ash L. R., Orihel T. C., 2007. Atlas of Human Parasitology. Chicago: ASCP Press. 540 p.
- Babenko A. N., Kuzmicheva E. A., Khasanov B. F., Rosen S., Kiseleva N. K., Savinetsky A. B., 2015. Chapter 15: Dung Deposits as Archives of Environmental Change // Lucke B., Bäumler R., Schmidt M. Soils and Sediments as Archives of Environmental Change. Geoarchaeology and Landscape Change in the Subtropics and Tropics. Erlanger Geographische Arbeiten. Bd. 42. P. 201–217.
- Beug H. J., 2004. Leitfaden der Pollenbestimmung für Mitteleuropa und angrenzende Gebie-te. München: Verlag Dr. Friedrich Pfeil. 542 S.
- Brooks J., Shaw G., 1972. Geochemistry of Sporopollenin // Chemical Geology. Vol. 10. P. 69–87.
- Brown A., Badura M., King G., Gos K., Cerina A., Kalnina L., Pluskowski A., 2017. Plant Macrofossil, Pollen and Invertebrate Analysis of a mid-14th Century Cesspit from Medieval Riga, Latvia (The Eastern Baltic): Taphonomy and Indicators of Human Diet // Journal of Archaeological Science: Reports. Vol. 11. P. 674–682. DOI: https://doi.org/10.1016/j.jasrep.2017.01.016
- Callen E. O., Cameron T. W. M., 1960. A Prehistoric Diet Revealed in Coprolites // New Scientist. Vol. 7. P. 35–40.
- Campbell G., Moffett L., Straker V., 2011. Environmental Archaeology. A Guide to the Theory and Practice of Methods, from Sampling and Recovery to Post-Excavation. 2nd ed. Swindon: English Heritage. 48 p.
- Carrion J. S., 2002. A Taphonomic Study of Modern Pollen Assemblages from Dung and Surface Sediments in Arid Environments of Spain // Review of Palaeobotany and Palynology. Vol. 120. P. 217–232.
- Cordova C. E., Gerasimenko N. P., Lehman P. H., Kliukin A. A., 2011. Late Pleistocene and Holocene Paleoenvironments of Crimea: Pollen, Soils, Geomorphology and Geoarchaeology // Geological Society of America Special Papers. Vol. 473. P. 133–164. DOI: https://doi.org/10.1130/2011.2473(09)
- Deforce K., 2017. The Interpretation of Pollen Assemblages from Medieval and Post-Medieval Cesspits: New Results from Northern Belgium // Quaternary International. Vol. 460. P. 124–134. DOI: https://doi.org/10.1016/j.quaint.2016.02.028
- DPDx – Laboratory Identification of Parasites of Public Health Concern. Stool Specimens – Intestinal Parasites: Comparative Morphology Tables, 2016. URL: https://www.cdc.gov/dpdx/diagnosticprocedures/stool/morphcomp.html (date of access: 03.10.2022).
- Florenzano A., 2019. The History of Pastoral Activities in S. Italy Inferred from Palynology: A Long-Term Perspective to Support Biodiversity Awareness // Sustainability. Vol. 11. P. 404–424. DOI: https://doi.org/10.3390/su11020404
- Foreyt W. J., 2001. Veterinary Parasitology Reference Manual. Ames: Wiley-Blackwell. 235 p.
- Grimm E. C., 1987. CONISS: A FORTRAN 77 Program for Stratigraphically Constrained Cluster Analysis by the Methods of Incremental Sum of Squares // Computers and Geosciences. Vol. 13 (1). P. 13–35.
- Grimm E. C., 2019. TILIA 2.6.1 Version (Computer Software). Springfield: Illinois State Museum. Jaeger L. H., Gijón-Botella H., del Carmen del Arco-Aguilar M., Martín-Oval M., Rodríguez-Maffiotte C., del Arco-Aguilar M., Iñiguez A.M., 2016. Evidence of Helminth Infection in Guanche Mummies: Integrating Paleoparasitological and Paleogenetic Investigations // The Journal of Parasitology. Vol. 102 (2). P. 222–228. DOI: https://doi.org/10.1645/15-866
- Karkanas P., Goldberg P., 2019. Reconstructing Archaeological Sites: Understanding the Geoarchaeological Matrix. Oxford: Wiley-Blackwell. 296 p.
- Kenward H., 2009. Northern Regional Review of Environmental Archaeology: Invertebrates in Archaeology in the North of England // Northern Regional Review of Environmental Archaeology, Research Department Report Series. No 12/2009. Portsmouth: English Heritage. 625 p.
- Komarek J., Jankovska V., 2001. Review of the Green Algal Genus Pediastrum ; Implication for Pollen Analytical Research // Bibliotheca Phycologica. Vol. 108. 127 p.
- Ledger M. L., Micarelli I., Ward D., Prowse T. L., Carroll M., Killgrove K., Mitchell P. D., 2021. Gastrointestinal Infection in Italy During the Roman Imperial and Longobard Periods: A Paleoparasitological Analysis of Sediment from Skeletal Remains and Sewer Drains // International Journal of Paleopathology. Vol. 33. P. 61–71.
- Lee C. M., van Geel B., Gosling W. D., 2022. On the Use of Spores of Coprophilous Fungi Preserved in Sediments to Indicate Past Herbivore Presence // Quaternary. Vol. 5. P. 30–53. DOI: https://doi.org/10.3390/quat5030030
- Meng X., Wang S., Zhou W., Wang B., Han W., Wang L., 1986. The Operculum-Plug Area and Membranous Structure of the Eggs of Trichuris Trichiura // Scanning Electron Microscopy. Vol. 3. P. 1015–1018.
- Reinhard K. J., 1992. Parasitology as an Interpretive Tool in Archaeology // American Antiquity. Vol. 57, iss. 2. P. 231–245.
- Slepchenko S. M., Gusev A. V., Ivanov S. N., Svyatova E. O., 2015. Opisthorchiasis in Infant Remains from the Medieval Zeleniy yar Burial Ground of XII–XIII centuries AD // Memorias do Instituto Oswaldo Cruz. Vol. 110 (8). P. 974-980. DOI: https://doi.org/10.1590/0074-02760150156
- Slepchenko S. M., Pererva E. V., Ivanov S. N., Klepikov V. M., 2019. Archaeoparasitological Analysis of Soil Samples from Sarmatian Burial Ground Kovalevka I, 2nd – 1st Centuries BСE, Russia // Journal of Archaeological Science: Reports. Vol. 26. DOI: https://doi.org/10.1016/j.jasrep.2019.101874
- Stockmarr J., 1972. Determination of Spore Concentration with an Electronic Particle Counter // Geological Survey of Denmark Yearbook. P. 87-89.
- van Amerongen Y. F., 2020. All’s Well? Comparing On- and Off-Site Pollen Samples and Exploring the Potential of Pollen from Man-Made Contexts // Vegetation History and Archaeobotany. Vol. 29. P. 125–131. DOI: https://doi.org/10.1007/s00334-019-00759-w
- Yoshikawa H., Yamada M., Matsumoto Y., Yoshida Y., 1989. Variations in Egg Size of Trichuris Trichiura // Parasitology Research. Vol. 75 (8). P. 649–654. DOI: https://doi.org/10.1007/bf00930964
- Zajac A. M., Conboy A. M., 2021. Veterinary Clinical Parasitology. Ames: Wiley-Blackwell Publ. 365 p.
