Robusticity of hand phalanges: relevance to the origin of the Altai Neanderthals

Author: Mednikova M.B., Shunkov M.V., Markin S.V.
Journal: Archaeology, Ethnology & Anthropology of Eurasia @journal-aeae-en
Section: Anthropology and paleogenetics
Article in issue: 3 т.45, 2017.
Free access
Short address: https://sciup.org/145145319
IDR: 145145319 | DOI: 10.17746/1563-0110.2017.45.3.126-135
Text of the article Robusticity of hand phalanges: relevance to the origin of the Altai Neanderthals
The diaphyseal structure of tubular bones of ancient and modern humans is usually considered in terms of their activity patterns. The biomechanical analysis of human remains, which is based on “engineering” approaches to assessing the level of robustness of skeletal elements, has been the most popular in recent decades (Ruff, 2000а: 71). The shape of bone is determined by many different factors. In early ontogeny, genes play the main role, while non-genetic factors can act generally or locally as well. The systemic change in skeletal robusticity can occur as a result of hormonal regulation, malnutrition, or pathology. Local hypertrophies reflect more limited and specific influences on the bone tissue, and can in some cases be used for the reconstruction of behavioral patterns.
The results of the research on the temporal dynamic of the inner robusticity of the greater tubular bones became the basis for the conception of the leading role of
Archaeological, paleogenetic, and anthropological researches of the recent years have led to numerous discoveries of new groups of Paleolithic humans, such as Denisovans, Altai Neanderthals, “hobbits” from Flores Island, and Homo naledi . The remains of some of these species are rather fragmentary, e.g. represented by isolated finger-phalanges. Comprehensive description and comparative analysis of these remains with the use of modern research techniques remains an important task.
The morphology of the lesser tubular bones (medial phalanges) of the hands of the Middle and Upper Paleolithic specimens from Europe and Asia has been the focus of the present study. Choosing such an object for our research, we assume that those phalanges display a significant correlation with long bones and, consequently, can be used as a “model” of the whole skeleton. This is particularly important when studying isolated and unique finds of fossil humans. Notably, the robustness of the bones of the upper limb, as compared to the lower limb, is less dependent on the total body-mass (Ruff, 2000b: 285). On the other hand, we can expect the bones of the hands of ancient humans, producing and repeatedly using stone tools, to display the most prominent structural changes due to the working hypertrophy of bone.
We set out to study variation of the inner robusticity of the medial phalanges of the hands of Neanderthals and Cro-Magnons, and to discuss possible factors shaping this variation.
Materials and methods
The bones of the 2nd and 4th digits of the hand composed the bulk of the sample for our comparative analysis. The sample included specimens belonging to various taxa of genus Homo : the Neanderthals of Europe and Asia, and the Early and Late Paleolithic H. sapiens (Table 1). Notably, not all of the individuals studied were living in periglacial conditions: the climate of the low-elevation zone of the Altai was milder than in Europe, and there were no glaciers there (Prirodnaya sreda…, 2003).
This study is based mostly on observations of the authors. The European Neanderthals are represented by the specimens from the Musée de l’Homme in Paris and the Museum of Anthropology and Ethnography (Kunstkamera) RAS in St. Petersburg. The remains of an adult male La Ferrassie 1 were found in Dordogne in 1909 (Capitan, Peyrony, 1912). The remains of an adult Neanderthal from Kiik-Koba 1, found in the Crimea in 1925, were identified and comprehensively described by the head of excavations (Bonch-Osmolovsky, 1941).
The Altai Neanderthals are represented by the skeletal remains from Okladnikov and Chagyrskaya caves.
Table 1. Sample
|
No. of specimen |
Specimen |
Taxon |
Hand medial phalanx |
Sex |
Age, years |
|
1 |
La Ferrassie 1 |
European Neanderthal |
4th digit, right |
Male |
40–55 |
|
2 |
Kiik-Koba 1 |
" |
2nd digit, right |
" |
40–49 |
|
3 |
Okladnikov 2 |
Altai Neanderthal |
4th digit, left |
" |
30–39 |
|
4 |
Okladnikov 5 |
" |
2nd digit, right |
Female |
25–39 |
|
5 |
Chagyrskaya Cave 16-3-12 |
" |
" |
" |
25–39 |
|
6 |
Kostenki 14 |
Upper Paleolithic H. sapiens |
4th digit, right |
Male |
25–29 |
|
7 |
Kostenki 8 TII 175 |
" |
3rd digit (?) |
" |
25–39 |
|
8 |
Kostenki 8 TII 173 |
" |
4th digit, right |
" |
25–39 |
|
9 |
Sungir 1 |
" |
2nd digit, right |
" |
35–45 |
|
10 |
Abri Pataud 26227 |
" |
" |
Female |
20–29 |
Archaeological findings provide the strongest evidence of a “ Mousteroid” pattern of the stone tool industries of these cave sites, which were named “Sibiryachikha facies of the Altai Middle Paleolithic” (Derevianko, 2011; Derevianko, Markin, Shunkov, 2013; Derevianko, Shunkov, Markin, 2014). From the anthropological specimens from Okladnikov Cave, mtDNA of Neanderthal type was extracted (Krause et al., 2007). A morphological examination of the specimens has confirmed their Neanderthal affinity (Viola, 2009; Mednikova, 2011a; Mednikova, 2011b, c). The fragments of the mandible (Viola et al., 2012) and the ulna (Mednikova, 2013a) from Chagyrskaya Cave also display Neanderthal anatomical features.
Okladnikov 2. A medial phalanx of the 3rd or 4th digit of the left (?) hand was found in the sq. Г-4 of the layer 4 at Okladnikov Cave in 1984. It is relatively short and broad, and hence fits well into the range of Neanderthals’ morphological variation (Mednikova, 2011b: 55). If it is a phalanx of the 3rd digit, then, according to the lengths and breadths of its proximal and distal epiphyses, it must have been male. But the breadth and height of its shaft put it closer to Neanderthal females. If it is a phalanx of the 4th digit, then it is even more probably male, since it is larger than phalanges of Shanidar males (Ibid.: 71).
Okladnikov 5. A medial phalanx from layer 3–1. Identified as a female right phalanx of the 2nd digit (Ibid.: 67–68). Like Okladnikov 2, it displays a flattened head and a relatively thick shaft (Ibid.: 70–71).
Chagyrskaya 16-3-12 . A medial phalanx, found at Chagyrskaya Cave in 2012, also exhibits some “archaic” features (Mednikova, Potrakhov, Bessonov, 2012). It was determined by the authors to be a female right phalanx of the second digit.
The Upper Paleolithic H. sapiens are represented in this study by one of the earliest anatomically modern individuals in Europe from the site of Kostenki 14 (Markina Gora) (Marom et al., 2012; Nalawade-Chavan, McCullagh, Hedges, 2014). The remains from the nearby site of Kostenki 8 (Telmanovskaya site) are far less famous. They were found by an expedition of the Leningrad branch of the Institute of Archaeology of the Academy of Science of the USSR, led by A.K. Rogachev, presumably in 1959. The larger specimen (TII 175), with destruction in its proximal portion, was identified as a medial phalanx of the 3rd (?) digit of the hand. The medial phalanx TII 173 is completely preserved, and belonged to the 4th digit of the right hand (Mednikova, Moiseev, Khartanovich, 2016). The remains of another early Eastern European H. sapiens (the male from Sungir 1) has been for decades the focus of research. The main features of the morphology of his hand have been discussed recently (Mednikova, 2012; Trinkaus et al., 2014). The medial phalanx from Abri Pataud (Musée de l’Homme, No. 26227) represents the morphology typical of later Western-European H. sapiens of the Proto-Magdalenian time.
The medial phalanges of all the individuals listed above were subjected to microtomography in order to study non-destructively the internal structure of the specimens. The La Ferrassie 1, 2, and Abri Pataud specimens were studied using the equipment of the Musée de l’Homme. All other fossils were scanned using the Xradia Versa XRM-500 X-ray 3D microscope at the laboratory of LLC “Systems for Microscopy and Analysis” (Moscow). The same scanner was previously used for studying the phalanx of the girl from Denisova Cave (Mednikova et al., 2013).
Virtual cross-sections at the middle of diaphysis have been compared in our study. We employed a number of variables conventionally used for description of the cross-sectional geometry of objects (the formula for the ellipse): area of the cortical layer (CA)—a measure of the resistance of a tubular bone to compression and tension; total area of the section (TA)—characterizes the external robusticity of the diaphysis of a phalanx; section area of medullary cavity (MA); and index of corticalization (% CA)—describes the inner robusticity of the diaphysis, i.e. relative thickness of the cortical layer. The polar moment of inertia (J) characterizes the resistance of the bone to compression and torsion. But the use of the latter variable for the reconstruction of biomechanical loadings in humans has been criticized (Lieberman et al., 2004), so we use it cautiously. Also, the polar moment of inertia and the length of the diaphysis (length of tubular bone) are correlated more strongly than are the length and area of the cortical layer (Ruff, 2000b: 284). In our opinion, the index of the relative development of the cortex of medial phalanges is the most informative variable. In order to reduce the influence of the total size of the tubular bones on our results, we will discuss variation of the index of relative development of the cortex in more detail.
Results
We examined 10 phalanges in total, among which there were male and female, large and small, robust and gracile specimens. The longest and thickest phalanges belonged to the Neanderthals La Ferrassie 1, Okladnikov 2, and H. sapiens TII 173, while the shortest and narrowest belonged to the Kostenki 14 and Abri Pataud 26227 individuals (Fig. 1).
The differences in general cross-sectional geometry between the specimens are already apparent during a visual examination of the microCT images (Fig. 2). In the vast majority of cases, the shape of the cross-section is elongated (sub-oval). The phalanges of La Ferrassie 1, Okladnikov 2 and 5 (Neanderthals), and (to a lesser extent)
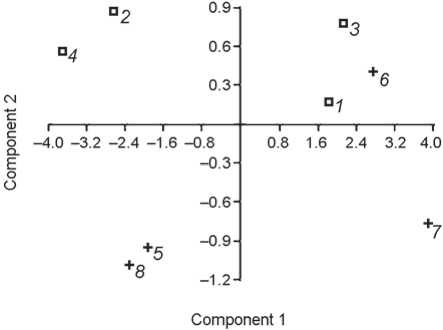
Fig. 1. Principal component analysis of the medial phalanx measurements. Variables: articular length, midshaft height, midshaft width, midshaft circumference.
1 – La Ferrassie 1; 2 – Kiik-Koba 1; 3 – Okladnikov 2; 4 – Chagyrskaya 16-3-12; 5 – Kostenki 14; 6 – Kostenki 8 (TII 173);
7 – Sungir 1; 8 – Abri Pataud 26227.
from Telmanovskaya and Sungir (Upper Paleolithic H. sapiens ) are the most oval in cross-section. It can be explained by the fact that the palmar ridges are not clearly developed at the level of the middle of diaphysis in all individuals: these external structures are best defined in La Ferrassie 1 and Kostenki 14 (both males), but far less prominent in Kiik-Koba 1, Chagyrskaya, Kostenki 8, and Sungir individuals.
On the basis of the total area of the section (TA), the males from La Ferrassie rock shelter (France) and Okladnikov Cave (Altai) display the most robust diaphyses. Among the Upper Paleolithic H. sapiens , TA decreases in the row from Kostenki 8 (both phalanges), Sungir, Kostenki 14, and Abri Pataud.
Abri Pataud 26227, Kostenki 14, and Okladnikov 5 are the individuals with the narrowest diaphyses. The first and third specimens are female from different taxa, the second (according to genetic data) is male.
The index of corticalization displays a notably wide range of inter-individual variation, which is typical of both Neanderthals and H. sapiens (Fig. 3, Table 2).
The % CA is less variable among the European and Siberian Neanderthals as compared to the individuals of the same species from the Altai, from Okladnikov and Chagyrskaya caves. The range of variation of the % CA among the Neanderthals is delimited by these two female individuals: Okladnikov is hyper-robust, while Chagyrskaya is hyper-gracile.
The % CA is quite variable among the European Upper Paleolithic H. sapiens as well, though the difference between the most thick-walled phalanx of Kostenki 14 and the most thin-walled phalanx of Sungir 1 is less, as compared to the same difference among the Neanderthals.
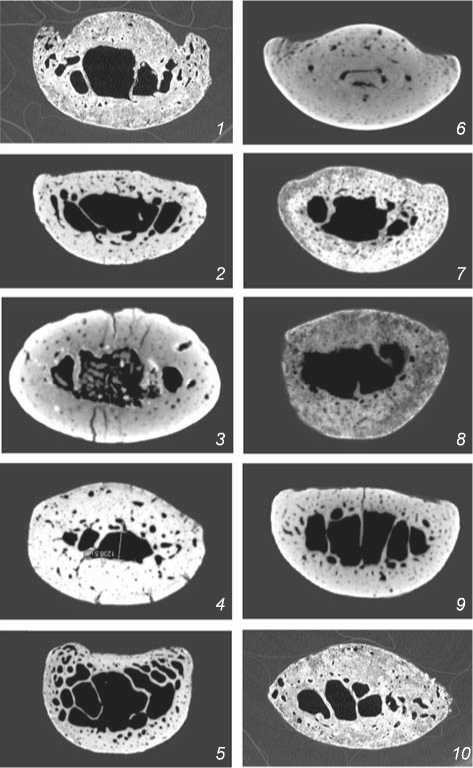
Fig. 2. Cross-sections of the medial hand-phalanges at the level of the middle of the diaphysis in Neanderthals ( 1 – 5 ) and European anatomically modern humans ( 6 – 10 ). The palmar surface is on the top. Numeration of the specimens is the same as in Table 1.
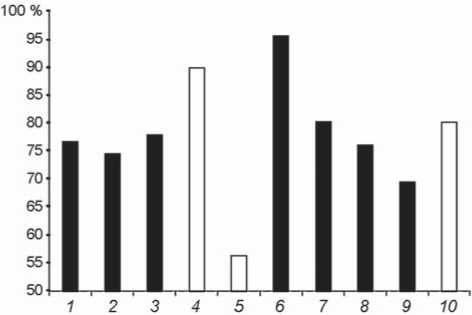
Fig. 3. Distribution of the index of corticalization (% CA) of the medial phalanges according to sex. Numeration of the specimens is the same as in Table 1. Black – males, white – females.
Table 2. Comparative description of inner robusticity of the hand medial phalanges at the level of the middle of the diaphysis in fossil Homo of different taxa
|
No. of specimen |
СА |
ТА |
МА |
% СA |
J |
|
1 |
41.48 |
54.17 |
12.69 |
76.57 |
507.52 |
|
2 |
28.96 |
38.93 |
9.97 |
74.39 |
255.19 |
|
3 |
35.36 |
45.41 |
10.05 |
77.88 |
330.65 |
|
4 |
33.88 |
37.7 |
3.82 |
89.87 |
231.58 |
|
5 |
22.92 |
40.70 |
17.78 |
56.32 |
231.71 |
|
6 |
34.33 |
35.93 |
1.61 |
95.6 |
2.47 |
|
7 |
38.17 |
47.62 |
9.45 |
80.15 |
373.22 |
|
8 |
34.53 |
45.45 |
10.92 |
75.97 |
333.52 |
|
9 |
26.37 |
37.89 |
11.52 |
69.61 |
230.82 |
|
10 |
26.47 |
33.02 |
6.55 |
80.17 |
168.46 |
Contrary to our expectations, the inner robusticity of the medial phalanges among the Upper Paleolithic H. sapiens was in general higher than in most Neanderthals. Also, the most thin-walled diaphysis was observed in the Neanderthal from Chagyrskaya.
Judging from these observations, we can make a preliminary conclusion: such factors as sex, taxon, and biological age do not affect the inner robusticity of the small tubular bones of fossil humans. This makes us consider the results and possible interpretations in more detail.
Discussion
Inner robusticity of small tubular bones as a function of biological age. The thickness of the walls of tubular bones depends on biological age, i.e. status of an individual. During ontogeny, the thickness of the walls is initially increasing, but then, after a stable period of maturity, post-definitive changes occur. These include gradual loss of bone, and thinning of the cortex, and finally lead to osteoporosis.
Importantly, owing to good preservation of the remains of the La Ferrassie 1 and Kiik-Koba 1 Neanderthals and the Sungir 1 and Abri Pataud H. sapiens , their ages are reliably confirmed by other skeletal elements. The La Ferrassie 1 “classical” Neanderthal is the oldest among the fossils studied. He exhibits numerous degenerative changes in various parts of the skeleton, and sutural obliteration. His biological age was estimated at 40– 55 years (Fennell, Trinkaus, 1997: 987). But the microCT of his medial phalanx has not shown the bone resorption typically associated with osteoporosis—neither in dorsal, nor in ventral walls of diaphysis, which are fairly robust.
The zones of intense bone-remodeling, visible on virtual cross-section of the phalanx, join the palmar ridges, and probably have a functional etiology, reflecting the development of the digital flexors (see Fig. 2, 1 ).
The Sungir 1 male is the oldest among the H. sapiens , though he is younger than the La Ferrassie 1 Neanderthal. According to an initial assessment made by G.F. Debets (1967: 160), the degree of obliteration of the cranial sutures of Sungir 1 correspond to an age of 55–65 years. V.V. Bunak (1973: 3) determined the age of this individual as “late maturity”. Analysis of various trait systems has shown that senile degenerative change can be excluded (Buzhilova, Kozlovskaya, Mednikova, 2000: 56). Another recent thorough examination of various age-markers suggests that the Sungir 1 male died between the ages of 35 and 45 years (Trinkaus et al., 2014: 76). This estimation was completely confirmed by a histological study of his bone tissue (Dobrovolskaya, Mednikova, 2015).
Summing up, even the oldest individuals in our sample do not exhibit signs of severe osteoporosis or, otherwise, maximal functional hypertrophy. Thus, the factor of biological age does not influence the degree of robusticity of the phalanges studied.
The role of physical exercise. In 1892, the German anatomist Julius Wolff hypothesized that the shape of a bone was a result of the displacement of bone-elements in the direction of functional tension, which can lead to either increase or decrease in bone-mass (Wolff, 1892). But the possibility of using “Wolff’s law” for assessing the level of physical activity has been debated. Taking into account that the process of gain/loss of bone mass is also affected by age, disease, hormonal status, and heredity (Ruff, Holt, Trinkaus, 2006: 485), the term “functional adaptation of bone” is more in use today.
The presence of the marginal ridges on the palmar surface of the hand phalanges is considered undeniable evidence of a great development of flexor muscles of the digits. Our results show that this trait varied among the Upper and Middle Paleolithic individuals. It can be reasonably supposed that the ridges become more prominent with age. Indeed, La Ferrassie 1 shows a strong hypertrophy of the ridges at the level of the middle of diaphysis. The young male from Kostenki 14 exhibits a very similar pattern. Considering this fact in terms of biomechanics, one should come to the conclusion that the Kostenki 14 individual was more hard-working than the Sungir one, as the bone tissue of the latter has not developed such “working hypertrophy” as a response to long years of physical exercise. This conclusion undoubtedly sounds paradoxical. However, such a formal approach seems to be erroneous, as it does not take into account individual variation in the development of the attachment-sites of digital flexors. For instance, the Kiik-Koba Neanderthal had palmar ridges, and fairly developed ones, but they are found proximal to the midshaft.
There is no doubt that the use and making of stone tools has had an outstanding biomechanical influence on the hand of Paleolithic man. Among numerous studies on this topic published in the last 150 years, the most remarkable are the recent works describing the suite of morphological traits favoring the most effective manufacture and use of scrapers and choppers (Marzke, 2013: 3). M. Marzke’s question is whether the human hand has, during its evolution, become adapted to the manufacture and use of stone tools. Today, there is a consensus among specialists that the adaptation to the use of tools cannot be considered as the only factor of natural selection driving the evolution of the humans’ hands. Different features of the human hand have evolved in different periods, and might reflect the foraging strategy of survival of the early humans. However, it is obvious that the hands of our ancestors have experienced a lengthy and strong influence from toolmaking.
Some modern experiments show the results of stone toolmaking. For example, a study of the activity of the 17 muscles, moving the hand and digits, during the use of Oldowan tools has shown that in early Homo the 1st and 5th digits experienced the maximal loadings (Marzke et al., 1998).
Exploitation of most Paleolithic tools implied two techniques of holding (Marzke, Shackley, 1986). In the first, holding the tool with the palm was accompanied by movements of the digits. In the second, the tool was strongly held with the thumb and the tips of the 2nd and 3rd digits. The robusticity, typical of the Neanderthal’s hand, was a result of the power control of a certain position of the digits, as well as of a permanent manipulation of tools. The hypertrophy of transverse dimensions of the phalanges, particularly distal ones, in Neanderthals indicates that they were probably more capable of a power transverse grip of the tool than were H. sapiens (Churchill, 2001; Niewoehner et al., 2003).
Even if all cultural and technological differences between the Middle and Upper Paleolithic populations are taken into account, it can be concluded that any physical loadings inevitably led to working hypertrophy of small tubular bones of hands. Some Upper Paleolithic individuals (e.g. Sungir 1), while belonging to H. sapiens , had certain “Neanderthaloid” features: widening of the ungual phalanges, robust ridges on the palmar surface of the proximal phalanx. These features point towards an intense biomechanical loading experienced by the right hand (Mednikova, 2012).
Another factor in the hand’s morphogenesis was the landscape typical of the areas inhabited by many populations of Homo in the Middle and Upper Paleolithic: these populations were usually living in mountainous or hilly terrains. Experimental data on rock-climbing might be important for understanding the skeletal features associated with the physical activity of humans. Modern qualified sport-climbers display a number of structural changes in their hands and certain digits (Sylvester, Christensen, Kramer, 2006). According to that study, climbing exerts an intense and lengthy mechanical influence on the hand’s bones. Specifically, a remodeling leading to a thickening of the cortex occurs in the metacarpals and phalanges. Compared to the control group, the climbers show an increased cross-sectional area and secondary moment of the area. They display a larger total width of the hand-bones, which is due to additional subperiosteal deposits but not to a widening of the medullary cavity. The robustness of the hand-bones is associated with modern styles of climbing and so-called “scrambling on large boulders”. These styles require maximal athletic training, which, in turn, leads to the strongest change in the hand.
The Neanderthals studied were living in mountainous conditions (cliffs of Dordogne, Crimea, Altai) and could have experienced the physical loadings described above. But the anatomically modern humans from Kostenki and Sungir were surrounded by the landscape of the East European Plain, which requires climbing skills much more rarely.
As can be seen, the hypothesis as to exclusively biomechanical influence on the inner robusticity of the hand phalanges cannot fully explain our observations.
Robusticity of the hand-bones of the Middle and Upper Paleolithic humans in the light of their origins. Thanks to the emergence of paleogenetics, we now know much more about the time of divergence of human clades, and about their genetic relationships. In our sample, three groups of the Middle and Upper Paleolithic humans are represented: “classical” European Neanderthals, Altai Neanderthals, and anatomically modern H. sapiens (AMH).
Let us consider the results of the present study in terms of the origins of these groups.
The “classical” Neanderthal from La Ferrassie 1 (as well as, probably, the Kiik-Koba 1 Neanderthal) represents the Eurasian human population of the time before contact with the AMH.
The Altai Neanderthals are a genetically separate group: Southern Siberia is part of the contact zone between Denisovans and Neanderthals. The results of a high-coverage sequence of genomes of humans from the Denisova Cave clearly document the genetic contacts between the two species (Krause et al., 2010; Prüfer et al., 2014).
There are remains from three Altai caves that were genetically assigned to Neanderthals. An overview of existing data on the postcranial remains of Altai Neanderthals shows a high level of inter-individual variability in this group of fossil humans and some trends of this variability (Mednikova, 2014, 2015). A proximal toe-phalanx and a distal finger-phalanx from Denisova Cave (Mednikova, 2011c, 2013b), as well as tubular bones of various individuals from Okladnikov Cave display a thickness of the walls that is exceptional even for Neanderthals. But the postcranial remains from Chagyrskaya Cave belonged to more “typical” Neanderthals (see, e.g., (Mednikova, 2013a)) and did not exhibit extraordinary inner robusticity.
The results of the present examination of the medial hand-phalanges fully confirm the previous conclusions regarding differentiation of the Southern Siberian Neanderthals into at least two morphological variants. It has been hypothesized that these variants might be related to different waves of migration of H. neanderthalensis to the Altai. On the other hand, it might be a result of genetic contact by the Neanderthals with humans of different taxa: for example, Denisovans (Mednikova, 2015). Taking into account the close proximity of the said caves, genetic contact between their inhabitants cannot be excluded either. In any case, the wide range of morphological variation is a phenotypic reflection of the complicated population history of the Altai Neanderthals.
The term “Altai Neanderthals” emerged during study of the complete genome of a female from Denisova Cave (Prüfer et al., 2014). We know that this individual belonged to the “hyper-robust” variant of the medullary cavity morphology (Mednikova, 2011c). It would be logical to assume that the thickening of the walls of tubular bones in that population was due not only to biomechanical loading, but also to the influence of hereditary factors: for example, admixture with relict Denisovans. However, the results of this study demonstrate that some European AMH were also characterized by thicker walls of small tubular bones, as compared to the “classical” Neanderthals and to the Chagyrskaya humans. This observation is based mostly on the morphology of the
Upper Paleolithic specimens from the sites in the Middle Don basin—among which the Kostenki 14 male is the most interesting, since his phalanges are particularly robust. The skeleton of this individual is very well preserved; for instance, the bones of both hands were present in full. A study of the skeleton has led to the conclusion that it was more robust as compared to modern Eurasian population (Mednikova, Moiseev, Khartanovich, 2016). A radiological examination has shown the presence of numerous medullary stenoses (i.e. displacement of the marrow by cortical bone) in the distal and medial phalanges of both hands, as well as in the proximal phalanges of the left hand (see Fig. 2, 6 ). A variant of congenital hypothyroidism seems to be the most plausible differential diagnosis. An extreme degree of medullary stenosis of tubular bones of the hand among the Upper Pleistocene human fossils was previously observed only in the ungual phalanx of the Neanderthal from Denisova Cave (Mednikova, 2013b)
A paleogenetic study has shown that the Kostenki 14 individual belonged to the metapopulation ancestral to many modern humans and Upper Paleolithic H. sapiens , including the Abri Pataud specimen. But the Kostenki 14 genome contains longer Neanderthal tracts (Seguin-Orlando et al., 2014).
Thus, weighting of the skeleton due to the thickening of the walls of tubular bones was observed among the Middle and Upper Paleolithic Eurasian individuals in two hybrid populations. One of them is considered as the ancestor of the Upper Paleolithic and, partially, modern Europeans, and displays a Neanderthal admixture. The second is the Neanderthals admixed with Denisovans (samples from Denisova and Okladnikov caves).
However, a recent study of the Altai Neanderthals’ genome has revealed some unexpected details of their “mixed ancestry”: an admixture from H. sapiens (Kuhlwilm et al., 2016). Before this study, it was widely accepted that the admixture between Neanderthals and AMH occurred outside Africa some 65–47 ka BP. M. Kuhlwilm and co-authors compared the genomes of the Altai Neanderthal and the Denisovan with sequences of 21 chromosomes of two Neanderthals from Spain and Croatia. They found out that a population that had early diverged from the rest of African (“sapient”) population made a contribution to the gene-pool of the ancestors of the Altai Neanderthals some 100 ka BP. But in Denisovans and the two European Neanderthals, such a “sapient” ancestry was not detected. According to these findings, it was concluded that the admixture between the Altai Neanderthals and modern humans took place probably in the Near East, some several tens of thousands of years earlier than was previously thought.
Thus, since the inner hyper-robusticity of the Kostenki 14 tubular bones is definitely not a Denisovan heritage, early AMH could have been the link between that individual (and, in broader sense, his metapopulation) and the Altai Neanderthals. Then, the hypertrophy of the walls of tubular bones was genetically determined. The origin of this genetic heritage was migration (in the case of Kostenki 14) or admixture (in the case of the Altai Neanderthals) from the AMH from a lower latitude.
Conclusions
At the beginning of our study, we expected to find confirmation for the hypothesis as to the increased inner robusticity of tubular hand bones in Neanderthals as compared to the Upper Paleolithic European H. sapiens . These expectations were based on the existing view that Neanderthals were relatively more capable of the powergrip of a tool and, finally, on the idea of the leading role of biomechanical influences in the thickening of the bone walls. However, our results demonstrate that the phalanges of the European AMH were usually more thick-walled. Contrary to our expectations, inner robusticity does not depend on either the sex or the age of an individual. Obviously, it does not depend on formal taxonomic status either. For instance, the Altai Neanderthals from Okladnikov Cave also exhibit an inner hyper-robusticity. If the results of the study of other small tubular bones are taken into account, the same applies to the Denisova Cave Neanderthals.
According to paleogenetic studies, the Altai Neanderthals of Denisova (and, we believe, Okladnikov) caves are to some extent the descendants of H. sapiens who lived more than 100 ka BP. Thus, our answer to the question “what Pleistocene Homo population was the ‘origin’ of the exceptionally robust skeleton of the Altai Neanderthals?” is: the early anatomically modern humans from the tropics.
This study has confirmed an earlier hypothesis as to the morphological heterogeneity of the Altai Neanderthals. This variation probably reflects the complicated population history of this group, which has acquired the genetic heritage of two other taxa. In future, other traces of admixture from Denisovans or early AMH might be detected through the study of the Altai Neanderthals’ morphology.
Acknowledgements
This study was supported by the Russian Science Foundation (Project No. 14-50-00036). Authors express their gratitude to A. Froment and the Musée de l’Homme in Paris for the opportunity of getting microCT images of the La Ferrassie 1 and Abri Pataud 26227 phalanges; and to V.I. Khartanovich (Museum of Anthropology and Ethnography (Kunstkamera) RAS in St. Petersburg) for permission to study the Kostenki 14 and 8 remains.
