Роль хронического экспериментального поражения печени матери в нарушении генеративной функции потомства

Автор: Брюхин Г.В., Сизоненко М.Л., Романов Ю.Н., Зубарев И.В., Ласьков Д.С.
Журнал: Человек. Спорт. Медицина @hsm-susu
Рубрика: Проблемы здравоохранения
Статья в выпуске: 8 (267), 2012 года.
Бесплатный доступ
Изучено влияние экспериментального поражения печени различного генеза у самок крыс на морфофункциональное становление генеративной функции потомства. Установлено, что патология печени матери в условиях эксперимента обусловливает нарушение фолликулогенеза в яичниках и сперматогенеза в семенниках потомства
Патология печени, потомство, сперматогенез, фолликулогенез
Короткий адрес: https://sciup.org/147153076
IDR: 147153076
The role of mother chronic experimental liver injury in disorder of the posterity generative function
The influence of female rats experimental liver injury of various genesis on morphofunctional status of the posterity generative function was studied. It was established that female liver pathology in experimental conditions cause the disorder of posterity folliclegenesis in ovaries and spermatogenesis in testes.
Текст научной статьи Роль хронического экспериментального поражения печени матери в нарушении генеративной функции потомства
Одной из важнейших причин перинатальной патологии являются экстрагенитальные заболевания женщин, в структуре которых особое место, в силу своей распространенности, занимают болезни гепатобилиарной системы [9].
Ранее нами было показано, что у самок крыс с хроническим экспериментальным поражением гепатобилиарной системы имеет место задержка наступления беременности, пролонгирование беременности до 28–29 дней. При этом наблюдалось снижение численности пометов, характеризующихся выраженными признаками физиологической незрелости [2], что проявляется нарушением морфофункционального становления систем жизнеобеспечения, в том числе иммунной, кроветворной, пищеварительной, нейроэндокринной [1].
Эти теоретические предпосылки явились причиной изучения особенностей становления генеративной функции у 60-дневного потомства самок крыс с экспериментальным поражением гепатобилиарной системы различного генеза.
Материал и методы исследования. Работа выполнена на белых лабораторных крысах «Вис-тар» (30 взрослых половозрелых самок) и их 60-дневом потомстве (60 крысят из 31 помета). Для достижения поставленной цели у взрослых половозрелых крыс-самок моделировалось хроническое поражение печени токсической и аутоиммунной этиологии.
Хроническое токсическое поражение («токсическая» – «Т»-группа) печени вызывалось путем однократного внутрибрюшинного введения взрослым половозрелым крысам Д (+)-галактозамина гидрохлорида («Sigma – G500», США) на 0,9 %-ном растворе натрия хлорида в дозе 250 мг/кг массы тела животного [3]. Экспериментальную «галактозаминовую» группу («Г»-группу) составили 20 крысят из 10 пометов.
Вторую группу составили животные, у которых создавался аутоиммунный процесс с преимущественным поражением печени («аутоиммунная» – «А»-группа) путем введения им 0,2 мл супернатанта шестидневной культуры E.coli (штамм АТСС 25922) [6]. Эту экспериментальную группу («Е»-группу) составили 17 крысят из 9 пометов.
О развитии патологического процесса в печени экспериментальных животных судили на основании морфологических изменений (очаговые бионекротические изменения гепатоцитов, дис-комплексация гепатоцитов, периваскулярная лим-фогистиоцитарная инфильтрация, гипертрофия и гиперплазия купферовских клеток), биохимических критериев (повышение концентрации билирубина, аланин- и аспартатаминотрансфераз и др.) и высокого титра печеночных аутоантител (1:640 и 1:1280).
Контрольную группу («К»-группа) составили интактные животные (23 крысенка из 12 пометов).
Объектом исследования явилось непосредственно потомство самок крыс с экспериментальным поражением гепатобилиарной системы на 60-й день после рождения, у которого проводилась оценка генеративной функции. Прежде всего осуществлялось определение весового индекса муж- ских и женских половых желез. Оценка генеративной функции семенников проводилась на серийных гистологических срезах, окрашенных железным гематоксилином по Гейденгайну по общепринятой методике [8] с определением площади семенных извитых канальцев, суммарного количества сперматогенных клеток и их субпопуляционного состава, а также количества канальцев со слущенным эпителием и числа гигантских сперма-тогенных клеток.
Одним из показателей, отражающих фертильность сперматозоидов, является характер их подвижности. В связи с этим нами проводилась оценка подвижности эпидидимальных сперматозоидов по общепринятой методике с определением числа неподвижных (0 баллов), «дергающихся» на месте (1 балл), слабоподвижных (2 балла) и прогрессивно-подвижных (3 балла) сперматозоидов. Для постановки этого теста зрелые сперматозоиды получали из придатка семенника [5]. Затем с использованием камеры Горяева в полученной взвеси подсчитывали сперматозоиды в 5 больших квадратах в течение трех часов через каждые 15 минут.
Для оценки фолликулогенеза в яичниках потомства экспериментальных животных на серийных гистологических срезах, окрашенных гематоксилином и эозином, а также железным гепаток-силином, проводился морфометрический анализ, позволяющий произвести дифференцированный подсчет овариальных фолликулов разных пулов. При этом использовалась общепринятая классификация Lintern Moore (1967), по методике Pedesen a Peters [10], основанная на оценке фолликула с определенным количеством фолликулярных клеток и его диаметра. Во избежание ошибок и повторов при подсчете учитывались овариальные фолликулы на максимальных сечениях ооцитов, маркерами которых служили ядрышки. Фолликулы, имеющие в окружении менее 10 клеток, относили к стадии 1–2 (покоящиеся); 1–20 клеток – к стадии 3а; 21–60 клеток – к стадии 3б; 61–100 клеток – к стадии 4; от 101 до 200 клеток – к стадии 5а; 201–400 клеток – к стадии 5б (растущие); 401–
600 клеток – к стадии 6; больше 600 – к стадии 7–8 (полостные). Для активности атрезии подсчитывали общее количество атретических фолликулов.
Полученные результаты обработаны статистически с использованием пакета прикладных программ «Statistica 6,0». Достоверность результатов оценивалась с помощью непараметрического критерии Манна–Уитни.
Результаты исследования и их обсуждение. Площадь поперечного сечения и диаметр семенных извитых канальцев оказались сниженными по сравнению с таковым в контроле. Так, если у интактных 60-дневных крысят диаметр семенных канальцев составил 234,47 ± 5,527, то у подопытных животных токсической и аутоиммунной групп исследуемый показатель составил всего 189,13 ± 5,196 и 198,78 ± 4,69 соответственно. Аналогичная закономерность выявлена и при изучении площади поперечного сечения канальцев. Так, у интактных крысят данный показатель составил 43574 ± 1592 мкм2, а у подопытных крысят всего 28387 ± 1005 мкм2 и 31298 ± 584 мкм2 соответственно.
Важнейшим показателем структурно-функционального состояния семенника является характеристика сперматогенного пласта. Сперматоген-ный пласт изучался нами с учетом суммарного содержания сперматогенных клеток и их субпопуляционного состава. Обнаружено, что у интактных крысят суммарное содержание сперматогенных клеток составляет 265,9 ± 33,53, а у подопытных животных как токсической (235,21 ± 3,29), так и аутоиммунной (242,15 ± 2,63) групп снижено.
Наибольший интерес представляют данные дифференцированного анализа сперматогенных клеток с учетом их зрелости [8].
Нами установлено, что у потомства самок крыс с экспериментальным поражением печени имеет место изменение субпопуляционного состава сперматогенных клеток по сравнению с контролем (табл. 1).
Как видно из табл. 1, какой-либо закономерности в изменении числа сперматогоний различной степени зрелости (А, П, Б) у эксперименталь-
Таблица 1
Субпопуляционный состав сперматогенных клеток семенных извитых канальцев семенников экспериментальных животных (M ± m)
|
Сперматогенные клетки |
Группа |
||
|
Контрольная (К) |
Опытная 1 (О1) |
Опытная 2 (О2) |
|
|
Суммарное содержание сперматогоний |
44,94 ± 0,86 |
13,36 ± 0,66* |
46,12 ± 3,00 |
|
Сперматогонии А |
22,38 ± 0,18 |
20,65 ± 0,31* |
20,72 ± 0,11* |
|
Сперматогонии П |
8,87 ± 0,18 |
9,08 ± 0,4* |
9,75 ± 0,22* |
|
Сперматогонии Б |
13,69 ± 0,3 |
13,63 ± 0,33* |
12,28 ± 0,49* |
|
Сперматоциты I и II |
194,57 ± 1,87 |
179,3 ± 2,5* |
179,79 ± 1,56* |
|
Сперматиды |
16,46 ± 1,19 |
8,61 ± 0,5* |
10,59 ± 0,79* |
|
Сперматозоиды |
9,94 ± 0,86 |
3,93 ± 0,49* |
6,53 ± 0,92* |
|
Суммарное кол-во сперматогенных клеток |
265,9 ± 3,52 |
235,21 ± 3,2* |
242,15 ± 2,6* |
Результаты статистически достоверны по сравнению с контролем (р < 0,05).
Проблемы здравоохранения
ных животных выявить не удается. В то же время содержание сперматоцитов 1 и 2 порядка, сперматид и сперматозоидов в семенных извитых канальцах подопытных крысят снижено по сравнению с интактными животными. Вместе с тем у подопытных крысят выявлено существенное увеличение по сравнению с контрольной группой числа семенных извитых канальцев со слущенным эпителием, являющегося чувствительным индикатором интенсивности апоптотических процессов. Так, если у интактных крысят содержание канальцев со слущенным эпителием составляет 7,11 ± 1,3, то у подопытных животных обеих групп исследуемый показатель оказался существенно увеличенным и составил 25,5 ± 3,1 и 27,44 ± 4,0. Заслуживают внимания также данные, свидетельствующие об увеличении в семенных извитых канальцах подопытных крысят числа гигантских сперматогенных клеток. Так у интактных крысят содержание таких клеток в канальцах составило 0,32 ± 0,02, а у подопытных животных обеих экспериментальных групп этот показатель оказался равным 0,64 ± 0,13 и 0,55 ± 0,08.
Таким образом, анализ результатов данной серии исследования позволяет констатировать, что у потомства самок крыс с экспериментальным поражением гепатобилиарной системы имеет место нарушение процесса сперматогенеза, о чем свидетельствует, прежде всего, снижение суммарного содержания сперматогенных клеток, а также количества сперматоцитов, сперматид и сперматозоидов в семенных извитых канальцах.
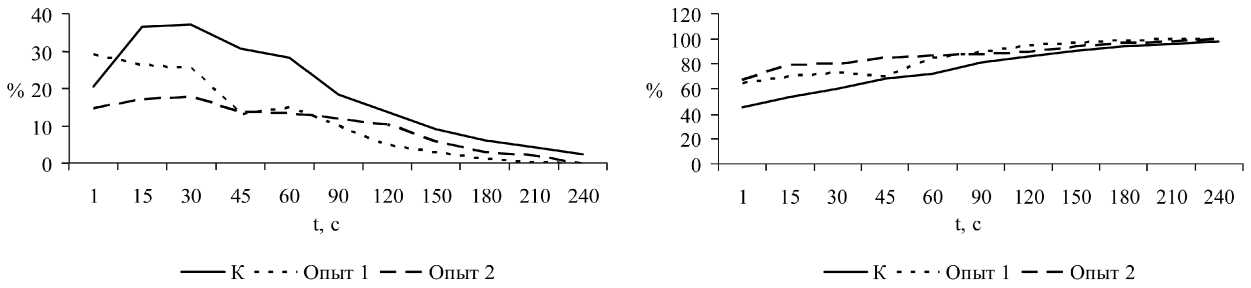
Анализ двигательной активности сперматозоидов позволил выявить следующую закономерность . У подопытных половозрелых крысят общее содержание в 1 мл эпидидимальных сперматозоидов снижено по сравнению с контролем (154,47 ± 2,74)·106 и составляет соответственно (69,24 ± 1,365)·106 и (99,29 ± 1,716)·106. Кроме того, у интактных крысят первоначально содержание прогрессивно-подвижных сперматозоидов составило 18,90 ± 0,691 %, в то время как у крысят опытной группы 1 этот показатель был равен всего 8,25 ± 1,876 %, а у животных второй опытной группы 1,86 ± 1,742 %. Через 15 минут после начала исследования у подопытных крысят отсутствовали прогрессивноподвижные сперматозоиды, в то же время у интактных животных число половых клеток данной популяции составило 0,25 ± 0,125 %. Аналогичная закономерность выявлена и при анализе содержания слабоподвижных сперматозоидов (рис. 1).
Таким образом, на всех сроках исследования у подопытных крысят обеих групп число фертильных сперматозоидов (прогрессивно-подвижных и слабоподвижных) снижено по сравнению с контролем.
При этом у подопытных крысят обеих групп содержание нефертильных («дергающихся» на месте и неподвижных) сперматозоидов на большинстве сроков исследования оказалось повышенным по сравнению с группой сравнения (рис. 2).
В следующей серии нашего исследования проведен анализ особенностей фолликулогенеза в яичниках потомства самок крыс с эксперимен-

Рис . 1. Динамика изменения содержания эпидидимальных слабоподвижных сперматозоидов
а)
б)
Рис. 2. Динамика изменения содержания эпидидимальных «дёргающихся» (а) и неподвижных (б) сперматозоидов
Таблица 2
Популяционный состав фолликулов по степени зрелости в яичниках экспериментальных животных (M ± m)
|
Показатель |
Группа |
||
|
К |
О1 |
О2 |
|
|
Суммар. кол-во фоллик. |
702 ± 53 |
526 ± 34 |
627 ± 58* |
|
Фолликулы 1 типа |
164 ± 28 |
80 ± 4 |
138 ± 22* |
|
Фолликулы 2 - 3 а типа |
90 ± 11 |
47 ± 6* |
58 ± 6* |
|
Фолликулы 3 б типа |
64 ± 5 |
40 ± 4 |
76 ± 7* |
|
Фолликулы 4 типа |
62 ± 6 |
74 ± 6* |
85 ± 5 |
|
Фолликулы 5 а типа |
96 ± 9 |
54 ± 3* |
52 ± 6* |
|
Фолликулы 5 б типа |
112 ± 16 |
107 ± 8 |
98 ± 6 |
|
Фолликулы 6 типа |
42 ± 3 |
40 ± 5* |
54 ± 6 |
|
Фолликулы 7– 8 типа |
96 ± 9 |
84 ± 7* |
66 ± 5 |
|
Малые фолликулы |
254 ± 25 |
127 ± 27 |
196 ± 46* |
|
Средние фолликулы |
222 ± 16 |
168±15 |
213 ± 14* |
|
Большие фолликулы |
226 ± 19 |
231 ± 24 |
218 ± 19 |
Результаты статистически достоверны по сравнению с контролем (р < 0,05).
тальным поражением печени (табл. 2). Одним их важнейших показателей, отражающих функциональное состояние яичников, является суммарное содержание фолликулов различной степени зрелости [4]. Нами установлено, что у подопытных животных суммарное содержание фолликулов снижено по сравнению с контролем.
Так, у интактных крысят этот показатель составил 702 ± 53, в то время как у подопытных животных и токсической (526 ± 34), так и аутоиммунной (627 ± 58) групп суммарное содержание фолликулов различной степени зрелости оказалось сниженным. При этом дифференцированный подсчет овариальных фолликулов позволил выявить определенную закономерность: у подопытных крысят содержание больших фолликулов практически не отличается от контроля, в то время как число малых фолликулов, являющихся резервом будущих половых клеток, существенно снижено (см. табл. 2). Уменьшение числа растущих фолликулов в яичниках подопытных крысят обеих групп в период становления половой зрелости согласуется с результатами, полученными при анализе числа атретических фолликулов в их яичниках. У потомства самок крыс с экспериментальным поражением печени различного генеза число атрети-ческих фолликулов существенно увеличено по сравнению с контролем. Так, у подопытных животных токсической и аутоиммунной групп число атретических фолликулов соответственно составило 39,54 и 31,67 %, в то время как у животных контрольной группы исследуемый показатель оказался равным 30,7 %.
Таким образом, результаты данной серии исследования позволяют констатировать, что у потомства самок крыс с экспериментальным поражением печени, в яичниках имеет место нарушение процесса фолликулогенеза, что, прежде всего, находит свое проявление в уменьшении числа рас- тущих фолликулов и как следствие уменьшение суммарного количества фолликулов, а также в усилении процессов атрезии фолликулов.
Заключение . Нарушение условий внутриутробного развития при моделировании поражения гепатобилиарной системы матери приводит к нарушению процессов формирования коррелятивных взаимоотношений в гипофизарно-тиреоидно-адреналово-половой системе плода, что обусловливает задержку структурного становления репродуктивной системы. Развивающийся комплекс изменений репродуктивной системы потомства может возникнуть в результате массированного проникновения через фетоплацентарный барьер патологических продуктов метаболизма при токсической патологии печени, а также сенсибилизированных цитотоксических Т-лимфоцитов, специфических антител, циркулирующих иммунных комплексов и других гуморальных факторов иммунной системы, что наиболее характерно для аутоиммунного поражения печени. Под влиянием этих факторов происходит нарушение процессов мейоза, усиление дегенеративной гибели половых клеток и уменьшение их суммарного количества. В целом, выявленные нарушения процессов сперматогенеза и фолликулогенеза позволяют говорить, что у самок крыс с экспериментальным поражением гепатобилиарной системы рождается потомство с нарушением становления репродуктивной функции.
Список литературы Роль хронического экспериментального поражения печени матери в нарушении генеративной функции потомства
- Брюхин, Г.В. Влияние хронических заболеваний печени матери на развитие потомства в условиях эксперимента/Г.В. Брюхин//Роль патологии печени матери в нарушении развития, реактивности и резистентности потомства в условиях эксперимента: сб. -Челябинск, 2009. -С. 6-16.
- Венгеровский, А.И. Механизм действия гепатопротекторов при токсическом поражении печени/А.И. Венгеровский, А.С. Саратиков//Фармакология и токсикология. -1988. -№ 1. -С. 89-93.
- Волкова, О.В. Морфогенетические основы развития и функции яичников/О.В. Волкова, Т.Г. Боровая. -М., 1999. -254 с.
- Егорова, Г.М. Токсикология новых промышленных химических веществ/Г.М. Егорова, Н.Г. Иванов, И.В. Саноцкий. -Л., 1966. -С. 33-36
- Моделирование воспалительного процесса в печени/Б.А. Сааков, А.И. Поляк, В.Е. Рычнев и др.//Моделирование, методы изучения и экспериментальная терапия патологических процессов. -М., 1967. -С. 119-123.
- Николаева, Л.Б. Первая беременность и первые роды: моногр./Л.Б. Николаева, Г.А. Ушакова. -М.: ГЭОТАР-Медиа. -2011. -264 с.
- Ухов, Ю.И. Морфометрические методы в оценке функционального состояния семенников/Ю.И. Ухов, А.Ф. Астраханцев//Арх. анат., гистол. и эмбриол. -1983. -№ 3. -С. 66-72.
- Шехтман, М.М. Руководство по экстрагенитальной патологии у беременных/М.М. Шехтман. -М.: Триада-Х, 2003. -816 с.
- Pedersen, T. Proposal for a classification of oocytes and follicules in the mouse ovary/T. Pedersen, H. Peters//J.Reprod. Fertil. -1968. -Vol. 17. -P. 555-557.
