Роль макрофагов в регуляции биосинтеза белка в клетках асцитной карциномы Эрлиха

Автор: Суменкова Д.В., Князев Р.А., Поляков Л.М., Панин Л.Е.
Журнал: Сибирский онкологический журнал @siboncoj
Рубрика: Лабораторные и экспериментальные исследования
Статья в выпуске: 2 (38), 2010 года.
Бесплатный доступ
Изучена роль перитонеальных макрофагов мышей с асцитной карциномой Эрлиха в поглощении и метаболической деградации белкового компонента липопротеинов высокой плотности. Обнаружено стимулирующее влияние кортизола на эти процессы. Показаны различия в спектрах внутриклеточных белков опухоль-ассоциированных макрофагов и перито- неальных макрофагов здоровых мышей, в частности, по содержанию аполипопротеина Е. Показано участие макрофагов в регуляции скорости биосинтеза белка в опухолевых клетках под влиянием липопротеинов высокой плотности и кортизола. Аполипопротеин Е может играть роль отрицательной обратной связи в данном механизме повышения скорости биосинтеза белка в опухолевых клетках.
Макрофаги, экспериментальная опухоль, биосинтез белка
Короткий адрес: https://sciup.org/14055609
IDR: 14055609 | УДК: 612.122.3:577.122:612.014:616-006.6
The role of macrophages in regulation of biosynthesis of proteins in cells of Еhrlich carcinoma
The role of peritoneal macrophages of mice with Ehrlich carcinoma in uptake and metabolic degradation of a protein component of high density lipoproteins has been studied. Stimulating effect of cortisol on these processes has been revealed. Differences in spectra of endocellular proteins between tumor-associated macrophages and peritoneal macrophages of healthy mice, in particular, in the level of apolipoprotein E have been shown. Participation of macrophages in the regulation of the protein biosynthesis rate in tumor cells under the influence of high density lipoproteins and cortisol has been shown. Apolipoprotein Е can play a role of a negative feedback in the mechanism of increasing the rate of protein biosynthesis in tumor cells.
Текст научной статьи Роль макрофагов в регуляции биосинтеза белка в клетках асцитной карциномы Эрлиха
Известно, что макрофаги играют важную роль в формировании противоопухолевой защиты, а их цитотоксическая активность связана с секрецией ряда цитокинов и активированных кислородных метаболитов. Одним из факторов, участвующих в реализации противоопухолевого эффекта макрофагов, может быть аполипопротеин Е, содержание которого составляет 10–25 % от общего секретируемого этими клетками белка [17]. В литературе аполипопротеин Е известен как ингибитор пролиферации клеток различных тканей, включая опухолевые [9, 11, 12, 13, 16]. Вместе с тем в последние годы получены данные об участии макрофагов в прогрессии опухоли. В ряде работ показано, что с ростом опухоли количество макрофагов, их противоопухолевая и антигенпрезентирующая активность снижаются [7, 10, 15]. Функциональные характеристики макрофагов модулируются опухолевым микроокружением, при этом клетки приобрета- ют способность поддерживать опухолевый рост, стимулируя ангиогенную активность опухоли, усиливая ее инвазивный и метастатический потенциал [8, 10, 14]. Таким образом, судьба опухоли во многом определяется функциональной активностью инфильтрующих ее макрофагов.
Ранее в Институте биохимии СО РАМН было открыто явление стимуляции резидентными макрофагами биосинтеза белка в паренхимных клетках органов и тканей в процессе регенерации [2]. В основе механизма этого явления лежит образование биологически активного комплекса «аполипопротеин А-I-тетрагидрокортизол». Показано, что макрофаги кооперативно захватывают липопротеины высокой плотности (ЛПВП) и кортизол. ЛПВП при участии лизосомальных гидролаз подвергаются дезинтеграции с освобождением аполипопротеина А-I, а кортизол подвергается действию внутриклеточных редуктаз с образованием тетрагидрокортизола. Полученные продукты формируют биологически активный комплекс, который, попадая в соматические клетки, взаимодействует с ДНК и усиливает экспрессию генов [4]. Мы полагаем, что этот механизм реализуется и в системе «макрофаг – опухолевая клетка» и может лежать в основе опухолевого роста. Изучению данного вопроса и посвящено настоящее исследование.
Материал и методы
В качестве экспериментальной модели использовали культуру клеток асцитной карциномы Эрлиха, выделенную из мышей линии CBA массой 15–20 г в лог-фазе опухолевого роста (Институт цитологии и генетики СО РАН, г. Новосибирск), в качестве контроля – макрофаги здоровых мышей, выделенные из перитонеального «лаважа». Мышей забивали под легким эфирным наркозом с помощью цервикальной дислокации. Свежевыделенные клетки ресуспендировали в среде RPMI-1640 («Биолот», Россия), рН 7,4, содержащей 20 мM HEPES («ICN Biomedicals, Inc», США), 2 мM L-глутамина («Вектор», Россия), 100 ед/мл пенициллина, 50 мкг/мл гентамицина, 5,6 мM глюкозы. Жизнеспособность клеток, оцениваемая методом исключения трипанового синего («Serva», Германия), составляла не менее 95 %. Инкубацию проводили в СО2-инкубаторе («Cole-Parmer», США) в атмосфере, содержащей 5 % СО2 и 95 % воздуха, при температуре 37ºС, используя 6-луночные планшеты («Orange Scientific», США). Плотность клеток составляла 1,5 тыс/мм2. Для получения культуры клеток асцитной карциномы, свободной от макрофагов, исходную суспензию клеток инкубировали 30 мин, а после адгезии макрофагов содержимое лунки переносили в другой планшет.
Скорость биосинтеза белка в культуре клеток асцитной карциномы определяли по включению 14С-лейцина («Amersham», Англия). Метку добавляли в количестве 2 мкКи/мл среды за 3 ч до окончания инкубации. Реакцию останавливали добавлением 0,2н раствора NaOH. Для измерения радиоактивности содержимое лунки переносили на целлюлозные фильтры («Whatman 3 MM», Англия), которые последовательно промывали от несвязавшейся метки раствором 10 % трихлоруксусной кислоты и смесью этанол-эфир (1:1).
Фильтры для измерения радиоактивности белка предварительно обрабатывали 0,1 М раствором лейцина в 10 % трихлоруксусной кислоте. Радиоактивность образцов измеряли на жидкостном сцинтилляционном счетчике («Mark-III», США) и выражали в имп/мин на 1 мг белка. Количественное определение белка проводили по методике Варбурга и Кристиана [1].
Для анализа спектра внутриклеточных белков макрофагов клетки лизировали раствором, содержащим 10 мМ фосфорнокислого натрия, рН 7,2, 85 мМ NaCl, 5 мМ KCl, 0,5 % дезоксихолат натрия и 1 % Тритон X-100. Выделение липопротеинов плазмы крови проводили методом изоплотностного ультрацентрифугирования в растворах KBr в присутствии 3 мМ ЭДТА-Na2 на центрифуге «Optima L-90K, Beckman-Coulter» (Австрия). Полученные липопротеины диализовали 24 ч против 0,05 М калий-фосфатного буфера, рН 7,4, содержащего 0,15 М NaCl и 0,3 мМ ЭДТА-Na2 при 4 ° С. Аполипопротеины А-I (апоА-I) и Е (апоЕ) получали путем делипиди-рования липопротеиновых фракций высокой (ЛПВП) и очень низкой плотности (ЛПОНП), соответственно, охлажденной смесью этанол-ацетон (1:1) с последующей многократной отмывкой эфиром и гель-фильтрации (колонка: 1,6 х 100 см, Сефароза CL-6B («Amersham Biosciences», Швеция), элюент: 0,01 М трис-HCl буфер, pH 8.6, содержащий 6 М мочевину, 0,01 % азид натрия, 1мМ фенилметансульфонил фторид). Анализ чистоты аполипопротеинов проводили методом диск-электрофореза в полиакриламидном геле с додецилсульфатом натрия («Serva», Германия). В качестве маркеров использовали набор низкомолекулярных белков-стандартов («Pharmacia», Швеция), включающий фосфорилазу – 94 кДа, альбумин – 67 кДа, овальбумин – 43 кДа, карбоангидразу – 30 кДа и лактальбумин – 14,4 кДа. Белковые полосы визуализировали 0,1 % Кумасси G-250 в смеси метанола и 10 % уксусной кислоты (1:1).
Обессоливание аполипопротеинов проводили с помощью гель-фильтрации (колонка: 20 х 0,8 см, Сефадекс G-25 («Pharmacia», Швеция), элюент: 0,05 М калий-фосфатный буфер, рН 7,4, содержащий 0,15 М NaCl). Иммуноэлектроблоттинг проводили «полусухим» методом с использованием нитроцеллюлозных мембран
(«Schleicher & Schuell», ФРГ) c диаметром пор 0,45 мкм. Специфические антитела против апоА-I и апоЕ, выделенных из плазмы крови человека, получали описанным нами ранее методом [3]. Электрофореграммы и данные иммуноэлектроблоттинга обрабатывали с помощью компьютерной программы TotalLab (Biosystematics, New Horizons in gel imaging and analysis). Статистическую значимость полученных результатов оценивали с помощью t-критерия Стьюдента при уровне значимости p < 0,05.
Результаты и обсуждение
При изучении роли макрофагов в регуляции биосинтеза белка в опухолевых клетках представлялось интересным сравнить спектры внутриклеточных белков перитонеальных макрофагов здоровых мышей и мышей с асцитной карциномой Эрлиха. Электрофоретический анализ внутриклеточных белков показал, что их спектры отличаются между собой (рис. 1). В белковом спектре опухоль-ассоциированных макрофагов было обнаружено меньшее количество низкомолекулярных и повышенное содержание средне- и высокомолекулярных фракций. Среди внутриклеточных белков следует отметить банд, соответствующий по молекулярной массе
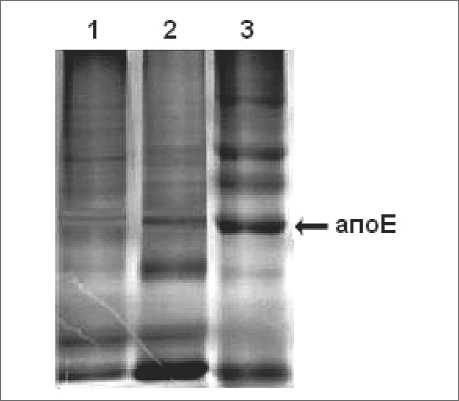
Рис. 1. Электрофореграмма внутриклеточных белков перитонеальных макрофагов мышей:
1 – опухоль-ассоциированные макрофаги; 2 – макрофаги здоровых мышей; 3 – белки ЛПОНП апоЕ (34 кДа). Этот банд был более выражен в лизате макрофагов здоровых мышей, что было подтверждено и методом иммунофер-ментного анализа. Иммуноэлектроблоттинг и денситометрический анализ показали, что в опухоль-ассоциированных макрофагах снижено содержание апоЕ в дегликозилированной проформе почти в 2 раза по сравнению с контролем, что может свидетельствовать о нарушении посттрансляционной модификации данного белка.
Для доказательства роли макрофагов в опухолевом росте и участия липопротеинов в этом процессе мы инкубировали клетки с ЛПВП (1 мг белка на 1 мл среды), выделенных из плазмы крови человека. Присутствие в инкубационной среде ЛПВП приводило к поглощению их макрофагами (рис. 2А). Среди множества внутриклеточных белков макрофагов в клеточном лизате появился белковый банд, соответствующий по молекулярной массе апоА-I (28,3 кДа). Этот банд по иммунохимической характеристике действительно соответствовал апоА-I человека, что было доказано методом иммуноэлектроблоттинга с использованием специфических антител (рис. 2Б, дорожки 2, 4). Кроме основной формы апоА-I, идентифицированы продукты метаболической деградации этого белка с молекулярными массами от 26 до 6 кДа. Количественный анализ поглощенного макрофагами белкового компонента ЛПВП показал, что опухоль-ассоциированные макрофаги отличаются значительно меньшей способностью к поглощению и метаболической деградации апоА-I.
Известно, что макрофаги обладают высокой активностью 5 α - и 5 β -редуктаз, восстанавливающих ∆-4,3-кетогруппу в кольце А стероидных гормонов с образованием тетрагидросоединений [6]. Продуктом метаболизма ЛПВП и кортизола в макрофагах является комплекс апоА-I с тетрагидрокортизолом. В связи с этим мы провели исследования по влиянию кортизола на поглощение и метаболическую деградацию белкового компонента ЛПВП. Оказалось, что кортизол в концентрации 1 × 10-6 М оказывает стимулирующее влияние на поглощение ЛПВП и метаболическую деградацию апоА-I как макрофагами здоровых мышей, так и макрофагами мышей с карциномой (рис. 2Б, дорожки 3, 5). Но
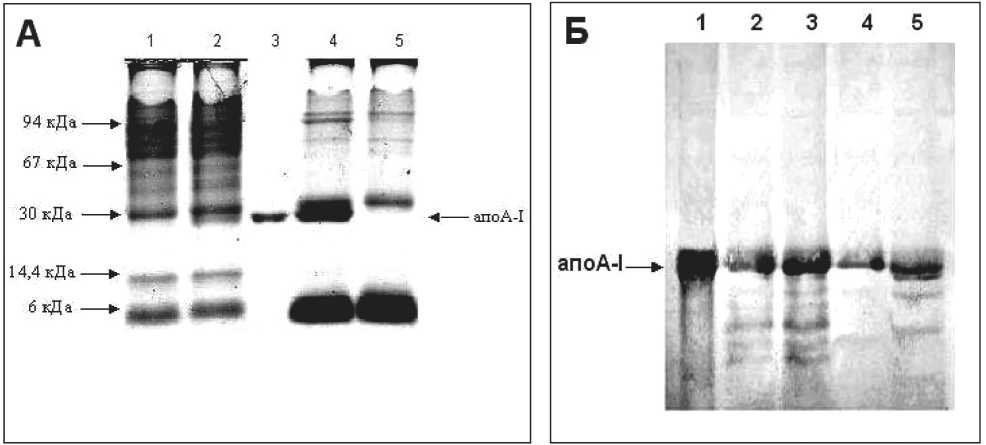
Рис. 2. Оценка поглощения и метаболической деградации белкового компонента ЛПВП перитонеальными макрофагами мышей: А – результаты электрофореза в 12,5 % ПААГ: 1 – опухоль-ассоциированные макрофаги; 2 – опухоль-ассоциированные макрофаги + ЛПВП; 3 – апоА-1 (стандарт); 4 – макрофаги здоровых мышей + ЛПВП; 5 – макрофаги здоровых мышей;
Б – результаты иммуноэлектроблоттинга с использованием специфических апоА-1-антител: 1 – апоА-1 (стандарт); 2 – макрофаги здоровых мышей + ЛПВП; 3 – макрофаги здоровых мышей + ЛПВП + кортизол; 4 – опухоль-ассоциированные макрофаги + ЛПВП; 5 – опухоль-ассоциированные макрофаги + ЛПВП + кортизол
при этом более выраженный эффект кортизола был отмечен для опухоль-ассоциированных макрофагов. Количественный анализ показал, что добавление гормона увеличивало поглощение ЛПВП макрофагами здоровых мышей в 1,6 раза, а опухоль-ассоциированными – в 2,2 раза. Таким образом, опухоль-ассоциированные макрофаги поглощают ЛПВП, метаболизируют их белковый компонент, а кортизол усиливает этот процесс.
Следующим этапом работы явилось изучение роли макрофагов в регуляции скорости биосинтеза белка по включению 14С-лейцина в клетках асцитной карциномы Эрлиха под влиянием ЛПВП и кортизола. Добавление ЛПВП и кортизола к клеткам асцитной карциномы, среди которых содержится 40 % опухоль-ассоциированных макрофагов, приводило к увеличению скорости биосинтеза белка по сравнению с контролем без добавок на 15 %, а по сравнению с культурой клеток карциномы с добавками, но без макрофагов – на 29 % (таблица).
Учитывая антипролиферативный эффект апоЕ, который секретируют макрофаги, и отмеченный нами факт снижения его содержания в опухоль-ассоциированных макрофагах, мы
Скорость биосинтеза белка в клетках асцитной карциномы Эрлиха
Таблица
|
Группа и условия инкубации |
имп/мин × 103/мг белка, M+m, n=6 |
|
1.Клетки карциномы |
816,1 ± 14,3 |
|
2.Клетки карциномы + ЛПВП + кортизол |
937,2 ± 35,4 * |
|
3.Клетки карциномы + апоЕ + ЛПВП + кортизол |
733,2 ± 28,3 * # |
|
4.Клетки карциномы без макрофагов + ЛПВП +кортизол |
728,1 ± 20,5 * # |
Примечание: * – различия статистически значимы по сравнению с группой 1 (р<0,05); # – различия статистически значимы по сравнению с группой 2 (р<0,05).
поставили задачу изучить влияние экзогенного апоЕ на скорость биосинтеза белка в опухолевых клетках асцитной карциномы Эрлиха. Добавление хроматографически очищенного апоЕ в концентрации 10 мкг/мл среды к клеткам асцитной карциномы приводило к снижению скорости биосинтеза белка (таблица). Следовательно, апоЕ подавлял эффект комплекса апоA-I-тетрагидрокортизол, образующегося в макрофагах из ЛПВП и кортизола, что согласуется с ранее полученными нами данными об ингибирующем влиянии апоЕ на биосинтез белка в культуре гепатоцитов здоровых крыс [5].
Таким образом, роль макрофагов в регуляции опухолевого роста можно рассматривать с позиции как ингибирующей, так и стимулирующей активности. На наш взгляд, это зависит от того, какая программа реализуется в большей степени: активация синтеза и секреции апоЕ, ингибирующего пролиферацию опухолевых клеток, или поглощение ЛПВП и стероидных гормонов с образованием биологически активного комплекса, усиливающего биосинтез белка в опухолевых клетках.
