Роль морфологических изменений паравентрикулярного ядра гипоталамуса и тиротропоцитов аденогипофиза в формировании тиреопатий

Автор: Полякова Л.В., Калашникова С.А., Самусев Р.П., Краюшкин А.И., Фогель А.В., Калашникова Е.А.
Журнал: Волгоградский научно-медицинский журнал @bulletin-volgmed
Статья в выпуске: 1 т.21, 2024 года.
Бесплатный доступ
Тиреопатии включают обширную группу заболеваний щитовидной железы, которые могут быть ассоциированы с синдромом хронической эндогенной интоксикации при различных соматических заболеваниях. Системное влияние эндогенных токсических соединений затрагивает также центральное звено регуляции эндокринной системы, где ведущую роль играет паравентрикулярное ядро гипоталамуса и тиротропоциты аденогипофиза. В связи с этим целью настоящего исследования было выявить закономерности изменений паравентрикулярного ядра и тиротропоцитов аденогипофиза. Исследование было выполнено на белых беспородных крысах самцах массой 120-200 г с моделированием хронической эндогенной интоксикации и оценкой периода после прекращения токсического воздействия. Животные выводились на 67, 90, 120 и 150-е сутки эксперимента с последующим гистологическим исследованием гипоталамуса и гипофиза. Было установлено, что морфологические изменения характеризовались проявлением неспецифических адаптационно-компенсаторных процессов паравентрикулярного ядра и тиротропоцитов, которые проявлялись в виде снижения биологической активности клеток и относительным ее восстановлением к окончанию эксперимента, что морфологически проявлялось в виде увеличения нейросекрета клеток паравентрикулярного ядра, увеличения процента полиядрышковых клеток, а также развития дистрофических изменений тиротропоцитов с появлением клеток «тиреоидэктомии». Относительная компенсация данных патологических изменений происходила на 150-е сутки эксперимента с восстановлением синтетической функции клеток, что проявлялось в виде нормализации ядерно-цитоплазматического соотношения и равномерного распределения субстанции Ниссля. Учитывая тот факт, что щитовидная железа находится под контролем гипоталамо-гипофизарной системы, данные патологические изменения могут играть ключевую роль в развитии тиреопатий при синдроме хронической эндогенной интоксикации.
Паравентрикулярное ядро, гипоталамус, тиротропоциты, гипофиз, щитовидная железа, тиреопатия, эндогенная интоксикация
Короткий адрес: https://sciup.org/142242050
IDR: 142242050 | УДК: 616-091.816
The role of morphological changes in the paraventricular nucleus of the hypothalamus and thyrotropocytes of the adenophypophysus in the formation of thyropathies
Abstarct. Thyropathies include an extensive group of thyroid diseases that may be associated with chronic endogenous intoxication syndrome in various somatic diseases. The systemic influence of endogenous toxins includes the important role in the regulation of the endocrine system, where the main place is the paraventricular nucleus of the hypothalamus and thyrotropocytes of the adenohypophysis. In this regard, the present study was identified in connection with changes in the paraventricular nucleus and thyrotropocytes of the adenohypophysis. The study was performed on white male rats weighing 120-200 g with modeling chronic endogenous intoxication and assessing the period after the onset of toxic effects. The duration of the experiment was the 67, 90, 120 and 150 days using histological technique for the hypothalamus and pituitary gland. It was found that morphological changes were characterized by the manifestation of nonspecific adaptive-compensatory processes of the paraventricular nucleus and thyrotropocytes, which manifested themselves in the form of a decrease in the biological activity of cells and its relative recovery by the end of the experiment, which morphologically manifested itself in the form of an increase in the neurosecretion of cells of the paraventricular nucleus, an increase in the percentage of polynucleolar cells, as well as the development of dystrophic changes in thyrotropocytes with the appearance of "thyroidectomy" cells. Relative compensation of these pathological changes occurred on the 150th day of the experiment with the restoration of the synthetic function of the cells, which manifested itself in the form of normalization of the nuclear-cytoplasmic ratio and uniform distribution of the tigoroid substance. Given the fact that the thyroid gland is under the control of the hypothalamic-pituitary system, these pathological changes can play a key role in the development of thyropathies in chronic endogenous intoxication syndrome.
Текст научной статьи Роль морфологических изменений паравентрикулярного ядра гипоталамуса и тиротропоцитов аденогипофиза в формировании тиреопатий
Морфофункциональная перестройка щитовидной железы (ЩЖ) находится под контролем гипоталамо-гипофизарной системы, где первичным уровнем регуляции является гипоталамус и его нейросекреторные ядра [1, 2]. В гипоталамусе имеется большое представительство ядер, которые вырабатывают либери-ны и статины, в том числе тиролиберин и тиро-статин медиальной части паравентрикулярного ядра гипоталамуса. Эффекты тиролиберина не ограничиваются эндокринными клетками гипофиза, о чем свидетельствует широкое представительство рецепторов (TRH-R1, TRH-R2) в различных отделах центральной нервной системы (кора головного мозга и мозжечка, гиппокамп, ствол мозга, спинной мозг), а также активация вторичных внутриклеточных мессенджеров через многокомпонентную систему рецепторов G-белка (GPCR). Известно, что только одна треть от всего вырабатываемого тиролиберина касается гипоталамо-гипофизарной оси, что не только приводит к морфофункциональным изменениям ЩЖ, но и регулирует функцию органов, находящихся под контролем отделов центральной нервной системы, вовлеченных в патогенез развития эндотоксинового шока, боли, а также физиологических циркадных ритмов, двигательной активности, вегетативной регуляции [3]. Таким образом, тиролиберин может играть важную роль в патогенезе соматических заболеваний, сопровождающихся синдромом хронической эндогенной интоксикации (ХЭИ). В экспериментальных исследованиях установлено, что при однократном введении бактериального липополисахарида (ЛПС) при полном отсутствии клинических симптомов была выяв- лена дизрегуляция секреции тиролиберина и тиреотропного гормона [4]. Известно, что при развитии инфекционных заболеваний, что подразумевает наличие эндогенной интоксикации, происходят нарушения в гипоталамо-гипофи-зарно-тиреоидной системе, однако данные о структурно-функциональных изменениях органов эндокринной системы отсутствуют. В связи с этим выявление структурных нарушений паравен-трикулярных ядер гипоталамуса и тиротропоци-тов гипофиза являются актуальной проблемой современной эндокринологии.
ЦЕЛЬ РАБОТЫ
Установить закономерности структурнофункциональных изменений органов гипотала-мо-гипофизарной системы крыс при хронической эндогенной интоксикации.
МЕТОДИКА ИССЛЕДОВАНИЯ
Нами были изучены органы центральной эндокринной системы (гипоталамус, гипофиз) 70 белых беспородных крыс массой 130–200 г при моделировании ХЭИ в течение 60 суток и после прекращения токсического воздействия длительностью 7, 30, 60 и 90 суток. Экспериментальные группы были подразделены следующим образом: 60, 60 + 7, 60 + 30, 60 + 60, 60 + 90 суток, контрольная группа. Для моделирования ХЭИ использовали модель сочетанного введения ЛПС и тетрахлорметана (ТХМ), разработанную в Волгоградском государственном медицинском университете [5].
Животные содержались в стандартных условиях согласно межгосударственному стандарту «Руководство по содержанию и уходу за лабораторными животными» ГОСТ 332162014, получали корма в соответствии с ГОСТ Р 50258-92 «Комбикорма полнорационные для лабораторных животных».
Выведение животных из эксперимента производили путем передозировки нембутало-вого наркоза с последующим забором органов для гистологического исследования, которое включало окраску гемато-ксилином и эозином, по Нисслю, по Маллори.
Анализ изображений производился с помощью аппаратно-программного комплекса для визуализации LASv4 фирмы Leica.
Статистический анализ был произведен с помощью программы Graph Pad Prism 8.0.1.244 с определением распределения нормальности по критерию Шапиро – Уилка с достоверностью p < 0,05. Морфометрические параметры включали в себя расчет объема ядра по формуле Vя =πlb2 / 6, где l – больший диаметр в мкм; b – меньший диаметр ядра в мкм; объема ядрышка Vя-ка = πd3 / 6, где d – диаметр ядрышка, мкм; объемной доли полиядрышковых клеток (ОД,%); объемной доли нейросекрета в клетках (ОД, %); ядерно-цитоплазматического соотношения (ЯЦО) по формуле ЯЦО = Sя/Sц, где Sя – площадь ядра клетки, мкм2; Sц – площадь цитоплазмы, мкм2 объемной доли сосудов (ОД, %).
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯИ ИХ ОБСУЖДЕНИЕ
При исследовании паравентрикулярного ядра гипоталамуса (ПВЯ) после прекращения токсического воздействия на 7-е сутки (группа 60 + 7) было установлено, что морфологические изменения нейросекреторных клеток включали достаточно плотное их расположение, где определялись крупные нейроны с отчетливо расположенным ядром и ядрышком (рис. 1).
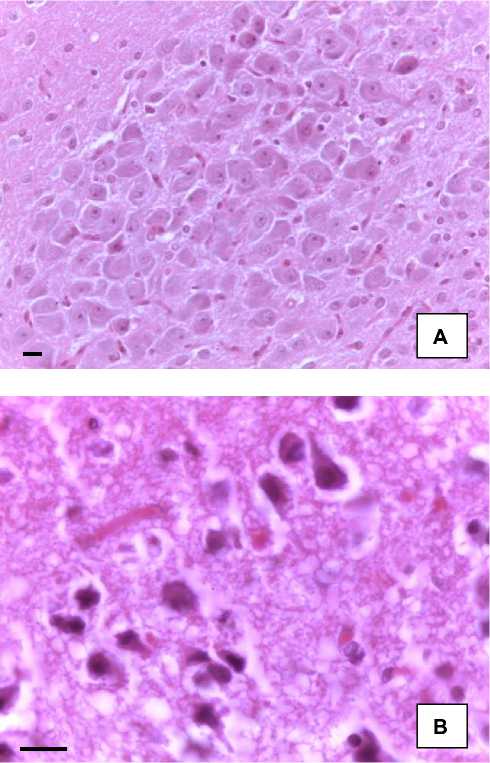
Рис. 1. Морфологические изменения ПВЯ гипоталамуса крыс после прекращения токсического воздействия: А, Б – группа 60 + 7; В, Г – группа 60 + 90. Окр. гематоксилином и эозином (а, г), по Нисслю (б, д).
Об. ×40; ок. 10. Об. ×100; ок. 10. Масштабный отрезок – 10 мкм
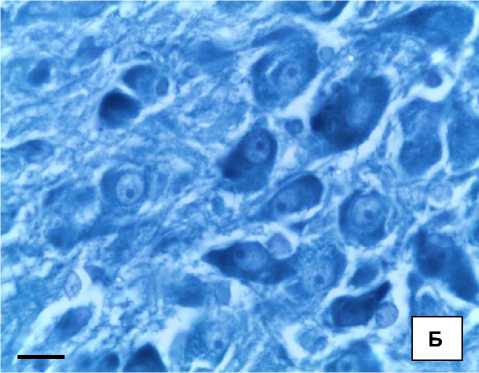
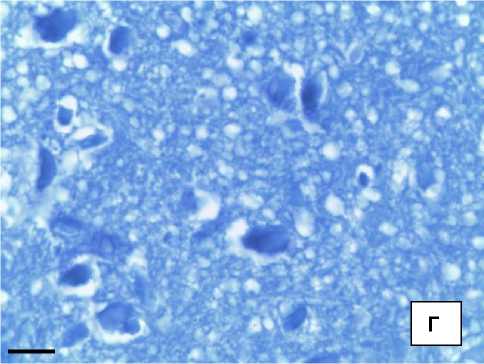
Увеличение объема клеток происходило за счет цитоплазмы, в которой при окраске по Нис-слю было выявлено увеличение ОД субстанции Ниссля в 1,4 раза (p < 0,05), что свидетельствовало об усилении синтетических процессов в клетке, так как по своей структуре оно представляет собой гранулярную эндоплазматическую сеть (грЭПС), что может свидетельствовать о развитии компенсаторно-приспособительной реакции в ответ на длительное повреждение (см. табл.). В связи с этим изменялся Vя, который увеличивался в 1,7 раза (p < 0,05) при от- носительной сохранности ядрышкового аппарата клетки. Однако число полиядрышковых ядер увеличивалось в 2,2 раза (p < 0,05). По мере увеличения сроков эксперимента на 30-е сутки после прекращения токсического воздействия (группа 60 + 30) происходило заметное снижение ОД нейросекрета в клетках, что, по-видимому, связано с функциональной перегрузкой нейронов, которое сочеталось с уменьшением Vя и Vя-ка в 1,5 и 3,4 раза соответственно (p < 0,05), что может свидетельствовать о развитии кариопикноза.
Морфометрические показатели нейроэндокринных клеток ПВЯ гипоталамуса, M ± m
|
Экспериментальные группы |
Морфометрические показатели |
|||
|
V ядер, мкм3 |
V ядрышек, мкм3 |
ОД полиядрышковых клеток,% |
Содержание нейроскрета в клетках,% |
|
|
Контрольная группа |
142,87 ± 7,11 |
5,71 ± 0,29 |
3,51 ± 0,13 |
22,63 ± 1,49 |
|
60 + 7 |
238,09 ± 11,68* |
5,73 ± 0,21 |
7,69 ± 0,38* |
30,78 ± 1,11* |
|
60 + 30 |
94,86 ± 4,66* |
1,66 ± 0,05* |
8,71 ± 0,43* |
17,91 ± 3,25 |
|
60 + 60 |
60,51 ± 2,01* |
2,16 ± 0,11* |
5,88 ± 0,29* |
19,43 ± 0,95 |
|
60 + 90 |
132,01 ± 6,59 |
3,05 ± 0,15* |
3,45 ± 0,17 |
21,13 ± 0,52 |
*p < 0,05 по сравнению с группой контроля.
Наряду с этим увеличивалось количество полиядрышковых клеток в 2,2 раза, что связано с изменением биологической активности клеток в условиях ХЭИ. Изменения морфометрических показателей в группе 60 + 60 характеризовались выраженными изменениями V я и V я-ка с уменьшением в 2,4 и 2,6 раз по сравнению с контрольной группой (p < 0,05).
Данные морфологические изменения соответствовали ядерной гиперхромии, клетки приобретали несколько угловатую форму и наблюдались незначительные явления перицеллюлярного отека.
Со стороны цитоплазмы клетки количество нейросекрета восстанавливалось и распределялось относительно равномерно.
В течение длительного периода после прекращения токсического воздействия в группе 60 + 90, по-видимому, происходило возмещение утраченных ультраструктур с относительным восстановлением ядерного аппарата клетки, о чем свидетельствовало изменение показателей V я и V я-ка , которые не имели достоверных отличий от показателей контрольной группы. Развитие адаптационно-компенсаторных процессво на фоне ХЭИ также проявлялось в снижении ОД полиядрышковых клеток.
Субстанция Ниссля располагалась достаточно компактно в цитоплазме клетки. Таким образом, морфологические изменения ПВЯ гипоталамуса являются обратимыми на фоне ХЭИ и сопровождаются частичным восстановлением нейросекреторных функций клеток.
При анализе следующего звена гипоталамо-гипофизарной системы – аденогипофиза, было установлено, что тиротропоциты располагались неравномерно и составляли от 1 до 5 % клеток в поле зрения, что не имело отличий от контрольной группы.
Известно, что тиротропоциты относятся к базофилам, имеют угловатую форму, где округлое ядро несколько вытянутое, расположенное в центре, а цитоплазма имеет зернистый вид за счет секреторных гранул.
Согласно морфологии клеток, с окраской по Маллори, нами было установлено, что в экспериментальной группе 60 + 7 наблюдались дистрофические изменения с увеличением ОД цитоплазмы (рис. 2) и, в отдельных случаях, формированием мелких вакуолей с появлением на более поздних сроках эксперимента так называемых клеток тиреоидэктомии, которые при различных интоксикациях могут увеличиваться до 16,75 %.
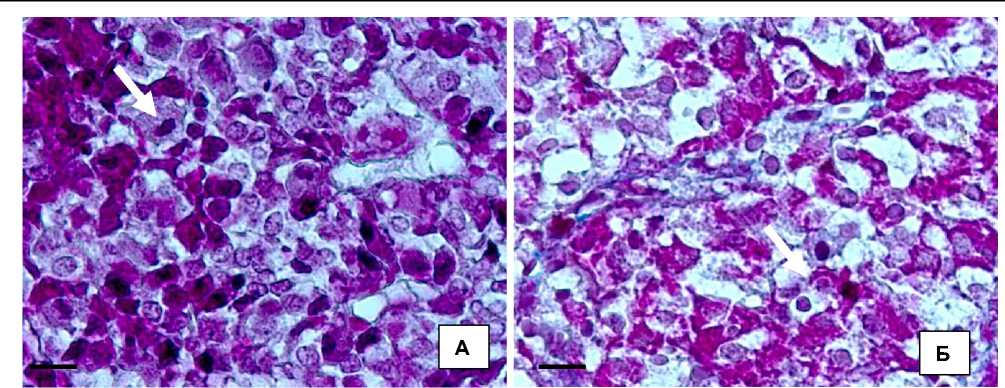
Рис. 2. Морфологические изменения тиротропоцитов аденогипофиза крыс:
А – дистрофические изменения тиротропоцитов в группе 60 + 7; Б – единичные апоптотические тельца и сохранение вакуольной дистрофии тиротропоцитов. Окр. по Маллори. Об. ×100; ок. 10. Масштабный отрезок – 10 мкм
Морфологические изменения тиротропо-цитов в группе животных 60 + 30, характеризовались нарастанием вакуольной дистрофии, где цитоплазма приобретала светлый вид с наличием единичных сохранных участков базофильной зернистой цитоплазмы. Ядро, по-прежнему, находилось в центре и имело вытянутую овальную форму. По мере увеличения сроков эксперимента после прекращения токсического воздействия в группе 60 + 60 наблю- дались единичные клетки тиреоидэктомии (гипертрофированные тиротропоциты), что свидетельствовало о стимуляции процессов восстановления при сохранении признаков ХЭИ.
При сравнительном анализе данных морфометрического исследования происходило снижение ЯЦО за счет нарастания объема цитоплазмы на 7-е сутки после прекращения токсического воздействия и достигало максимума к 30-м суткам (рис. 3).
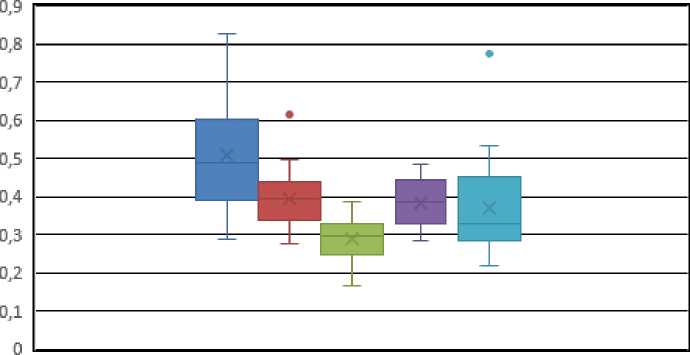
■ контроль ■ 60+7 ■ 60+30 ■ 60+60 ■ 60+90
Рис. 3. Морфометрические показатели ЯЦО тиротропоцитов аденогипофиза крыс на различных сроках эксперимента
В дальнейшем ЯЦО частично восстанавливалось с сохранением единичных клеток в состоянии вакуольной дистрофии, чем обу- словлено отклонение от показателей группы контроля. Так называемые «точки выброса» на сроке эксперимента 60 + 7 и 60 + 90 находятся в пределах значений контрольной группы, что свидетельствует о развитии компенсаторноприспособительных реакций клеток в условиях ХЭИ.
При сравнительном анализе морфологических изменений в ПВЯ гипоталамуса на ранних сроках эксперимента (60 + 7) наблюдалось сохранение общего плана строения клеток с увеличением ОД субстанции Ниссля, а также появлялись полиядрышковые клетки в ответ на длительное повреждение эндогенными токсическими соединениями, что свидетельствует об усилении белково-синтетических функций и может быть связано с адаптивными изменениями [6].
По мере увеличения сроков ХЭИ (60 + 30) наблюдалось относительное истощение нейронов ПВЯ с уменьшением ОД субстанции Ниссля, что может свидетельствовать о ее частичном растворении при развитии набухания, смещением ядра и истощением синтетического потенциала клеток, что подтверждалось морфометрическими показателями.
Интересен тот факт, что при таком же сроке ХЭИ, но при увеличении периода так назы-ваесмого «покоя» без введения препаратов (60 + 60), нейроны ПВЯ относительно восстанавливались, о чем свидетельствовало равномерное распределение субстанции Ниссля в цитоплазме клетки. Наличие компенсаторноприспособительных реакций нейронов ПВЯ к 90-м суткам на после прекращения токсического воздействия подтверждалось также морфометрическими данными, которые не имели отличий от показателей контрольной группы.
Проводя параллель с изменениями в аденогипофизе, можно заключить, что реакции ти-ротропоцитов в ответ на повреждение имели аналогичную тенденцию, где нарастали морфологические признаки неспецифического повреждения клеток в виде вакуольной дистрофии и появлением гипертрофированных тиро-цитов, популяция которых впоследствии снижалась к 90-м суткам после прекращения токсического воздействия.
Основываясь на полученных данных, можно предположить, что данные морфологические изменения гипоталамо-гипофизарной оси приводят к развитию гормонального дисбаланса, отмеченного при токсических воздействиях [7], с последующим влиянием на ЩЖ, и могут лежать в основе формирования тиреопатий, что требует дальнейшего изучения.
ЗАКЛЮЧЕНИЕ
Таким образом, на ранних сроках эксперимента после прекращения токсического воздействия нейросекреторные клетки ПВЯ гипоталамуса повышали свою биологическую активность, что проявлялось в увеличении морфометрических показателей структур, ответственных за синтетическую функцию. По мере увеличения срока эксперимента происходило относительное истощение физиологического резерва гормонов, о чем свидетельствовало снижение нейросекрета в клетках ПВЯ, а также появление клеток «тиреоидэктомии».
Следует отметить, что к 90-м суткам после прекращения токсического воздействия наблюдалась относительная компенсация белковосинтетических процессов в клетке при сохранении дистрофических изменений, что проявлялось в стабилизации ядерного комплекса и восстановлении содержания нейросекрета ПВЯ. Появление апоптотических телец, вакуольная дистрофия и изменение ЯЦО тиротропоцитов свидетельствует об изменении влияния на ЩЖ, что может лежать в основе перестройки тиреоидной паренхимы с формированием тиреопа-тий на фоне ХЭИ. Полученные данные свидетельствуют о том, что при отсутствии коррекции синдрома ХЭИ вероятно развитие морфологических изменений ЩЖ, которая находится под контролем гипоталамо-гипофизарной системы, с развитием гормонального дисбаланса периферического звена эндокринной системы.
Список литературы Роль морфологических изменений паравентрикулярного ядра гипоталамуса и тиротропоцитов аденогипофиза в формировании тиреопатий
- Полякова Л. В., Калашникова С. А., Фогель А. В. Морфогенез тиреопатий по данным экспрессии OCT3/4, PHH3, CK8, CK18 при хронической эндогенной интоксикации. Вестник Волгоградского государственного медицинского университета. 2023; 20(2):105-108. EDN: PDCXMZ
- Трошина Е. А., Мельниченко Г. А., Сенюшкина Е. С., Мокрышева Н. Г. Адаптация гипоталамо-гипофизарно-тиреоидной и гипоталамо-гипофизарно-надпочечниковой систем к новому инфекционному заболеванию - COVID-19 в условиях развития COVID-19-пневмонии и/или цитокинового шторма. Клиническая и экспериментальная тиреоидология. 2020;16(1 ):21-27. EDN: ISXSTK
- Hershman J. M., Beck-Peccoz P. Discoveries Around the Hypothalamic-Pituitary-Thyroid Axis. Thyroid. 2023;30. DOI: 10.1089/thy.2022.0258
- Mikofajczyk A., Zfotkowska D. Subclinical Lipopolysaccharide from Salmonella Enteritidis Induces Dysregulation of Bioactive Substances from Selected Brain Sections and Glands of Neuroendocrine Axes. Toxins (Basel). 2019;11(2):91.
- Новочадов В. В., Писарев В. Б. Эндотоксикоз: моделирование и органопатология: монография. Волгоград: Изд-во ВолгГМУ, 2005. 240 с. EDN: QMOLAL
- Смирнов А. В., Бисинбекова А. И., Файбисович Т. И. Морфофункциональные изменения головного мозга при сахарном диабете. Вестник Волгоградского государственного медицинского университета. 2022;19(3):3-8. EDN: KKQNGY
- Гуров Д. Ю., Туманов В. П., Смирнов А. В. и др. Морфологические изменения нейронов супраоптических ядер гипоталамуса крыс, предрасположенных к алкогольной зависимости. Современные проблемы науки и образования. 2019;2. URL: http://www.science-education.ru/ru/article/view?id=28750.
