Secular and age-related dynamics of biological characteristics in the rural population of Mongolia: a study of environmental adaptation

Author: Batsevich V.A., Yasina O.V., Sukhova A.V.
Journal: Archaeology, Ethnology & Anthropology of Eurasia @journal-aeae-en
Section: Anthropology and paleogenetics
Article in issue: 1 т.46, 2018.
Free access
Short address: https://sciup.org/145145350
IDR: 145145350 | DOI: 10.17746/1563-0102.2018.46.1.144-153
Text of the article Secular and age-related dynamics of biological characteristics in the rural population of Mongolia: a study of environmental adaptation
Answering the questions about how well human populations of various times and places are adapted to environment, and what factors affect the process of adaptation most, is the main, if not single, aim of anthropoecology. The former question can be gradually answered by collecting the data describing various adaptive changes in populations. The diversity of these changes is great, but not boundless. During the several last decades, dozens of populations have been studied from the points of view of morphology, physiology and biochemistry. The range of intergroup variation in Eurasia is generally determined by now (Alekseeva, 1986: 89–149; 1998: 112–196;
Antropoekologiya Tsentralnoy Azii, 2005: 85–237; Antropoekologiya Severo-Vostochnoy Azii, 2008: 238–312). But the possibility of determining the main factors affecting the adaptive process is the most important and the most challenging at the moment. It will be discussed in detail below.
The adaptation in human populations is specific. During early stages in the history of humanity, climatic factors dominated (Alekseeva, 1986: 150–154). But the role of collective labor and sociogenesis gradually increased in the course of human evolution (Alekseev, 1984: 348–383; Lambert, 1987; Khrisanfova, Perevozchikov, 1999: 108–119). The ultimate action of both abiotic and biotic environmental factors is strongly limited or even neglected by the economic
and cultural practices of human societies. According to T.I. Alekseeva, “…adaptation to environment at all stages of human society’s development has been brought about via two ‘channels’: biological and social, with the latter being dominant” (1972: 105). Further, Alekseeva outlines the following main features of well-adapted populations: equilibrium between the population and its environment, high survival rate, stability of morphofunctional status, and optimal accordance of economic and cultural customs with the ambient environment (Ibid.). These ideas were developed and detailed further by a number of authors (Okruzhayushchaya sreda…, 1979: 8; Prokhorov, 1991: 59).
In the modern world, the influence of humans on the transformation of the biosphere is growing at a pace exceeding the rate of biological evolution in living organisms, including humans. As a result, old ecological niches are disappearing, while new ones are being formed, and these latter are more difficult to adapt to. A conflict emerges between environment and adaptive features developed in previous generations but useless at present. In the last decades, both anthropologists and physicians have noted the widespread changes of somatic features emerging as results of developmental acceleration, as well as a sharp increase in the prevalence of diseases previously rare in youth: obesity, diabetes, allergies, autoimmune diseases, psychological dysfunctions (Auxology…, 2013: 110–129; Brüne, Hochberg, 2013).
In anthropoecology, attention is paid primarily to specific adaptive features. These include adaptive variation of morphological and physiological variables, particularly specifics of growth and development in populations occupying particular environments.
Owing to the large diversity of environmental factors, adaptive processes in human populations can only be evaluated through complex morphofunctional studies. When using other sources of data, special attention is also paid to the study of the tempos of growth, all stages of which are informative from the adaptive point of view, because all adaptive morphophysiological traits emerge and start functioning during ontogeny. The object of analysis in this study is traditional anthropometric variables, but also age-related changes of the hand bones that are identified and fixed, not only in childhood and adolescence, but also later in ontogeny (Rokhlin, 1936: 21–36, 181–190). The OSSEO technique developed for X-ray data can be successfully used for evaluation of the adaptive status of populations as well (Pavlovsky, 1987: 228–234; Pavlovsky, Maksinev, Batsevich, 1998; Batsevich et al., 2006; Kobyliansky, Livshits, Pavlovsky, 1995).
It was previously shown that in populations with increased longevity the disharmony of growth processes is absent, while maturation and aging of the hand bones occurs more slowly and age-related bone pathologies are rare (Batsevich et al., 2009, 2014; Kalichman et al., 2011). These data are in good agreement with the results of complex studies (demographical, socio-ethnographical, and biomedical) in centenarian populations of Abkhazia and Azerbaijan (Fenomen dolgozhitelstva, 1982: 59; Abkhazskoye dolgozhitelstvo, 1987: 94; Dolgozhitelstvo v Azerbaydzhane, 1989: 171). Judging by the results of the above-mentioned research, the populations with increased longevity can be confidently considered the most adapted groups of humans in terms of biology, demography, and socio-ethnography. These data, combined with knowledge about the morphophysiological peculiarities of the populations of particular regions (adaptive types), can be used as references for a comparative evaluation of the ecological status of the studied groups.
This study sets out to determine the adaptive status of rural populations of Mongolia, inhabiting various climates and keeping to traditional ways of life up to present time. To these ends, a comparative analysis of the rate of age-related skeletal changes was carried out in four samples of the adult Khalkha Mongols and one sample of the Khotons. The temporal dynamics of a number of morphophysiological features was studied in every population and in all Khalkha samples combined.
Materials and methods
Anthropometric and X-ray data were collected during the anthropoecological expeditions carried out by a joint team of the Research Institute and Museum of Anthropology (MSU) and the Institute of Biology of the Mongolian Academy of Sciences in 1986–1990. The studied area included Uver-Khangay (Bat-Ulziy and Burd somons, combined), Bayankhongor (Bayan-Lig village), Khubsugul (Zhargalant village), Ubsunur (Tarialan village), and Eastern (Khalkhgol village) aimags of Mongolia. The sample included 970 males and females from
18 to 80 years of age. The studied Mongolian populations are ethnically homogeneous, they all belong to the Khalkha Mongols, the largest ethnic group of the country. The Khotons from the Tarialan somon are descendants of migrants who were forcibly moved in the 18th century to what is now Western Mongolia (Grumm-Grzhimaylo, 1926: 274). From the point of view of physical anthropology, linguistics, and religion, the Khotons are similar to Central Asian groups of Eastern Turkestan, though their exact origins are still unknown. Originally, they were agriculturalists, but herding has gradually taken over. At the time of the expeditions, all studied populations shared the traditional nomadic and semi-nomadic economy of arid zones. As in the past, most food was produced domestically, while social differentiation between and within groups was minimal. Thus, the studied Mongolian populations occupy different geographical zones but share many cultural features, and therefore may serve as a good model for exploring anthropoecological associations.
All data were collected following the principles of bioethics. According to the law on personal data, all entries were anonymized before performing statistical analyses. Anthropometric data were collected following the standard protocol (Smirnova, Shagurina, 1981). The original OSSEO technique for the evaluation of age-related changes to the hand bones in individuals and populations was developed at the Research Institute and Museum of Anthropology (MSU) and described in a number of publications (Pavlovsky, 1987: 9–32; Pavlovsky, Maksinev, Batsevich, 1998; Batsevich et al., 2009). Some details of the technique, important for understanding the results of the present study, are outlined below.
The following variables (points) of the age osteomorphic status were evaluated:
A – the empirical age of the beginning of age-related changes in a population after full maturation of the hand skeleton. Late emergence of such changes is considered an advantage;
B – the empirical age at which all individuals in a population display at least some age-related changes to the hand bones. Again, this stage is achieved at an older age if the ontogenetic dynamic is optimal in the population;
B – A – the difference between the two ages (in years). Describes the period during which both individuals with age-related changes and without them are observed in a population. We hypothesize that the duration of this period might be proportional to the level of ontogenetic harmoniousness.
Standard samples were formed in each group for both sexes. The term “standard sample” in the OSSEO means individuals in the age range between A (see above) and 60 years. Such samples are used in order to exclude the possible influence of a difference in age distribution between samples on the results. The rationale behind this methodology is comprehensively described elsewhere (Antropoekologiya Tsentralnoy Azii, 2005: 214).
Tempo of age-related changes of the hand skeleton
First manifestations of age-related skeletal changes (the empirical point A) appear relatively early in both males and females of the studied populations, at 26– 28 years of age (Table 1). There is a marked intergroup variation in this variable among the populations, more pronounced in females. The Kalkha Mongols from Bayan-Lig and Zhargalant and the Khotons from Tarialan reach point A earlier.
The second post-definitive stage (point B), during which all individuals in a sample display manifestations of aging, begins at different ages in different populations (Table 1). In males, it ranges from 38 to 52 years, and in females from 35 to 55 years. This variation covers almost fully the range of values of this variable previously observed in 46 populations from various ecological conditions (Table 2). A relatively earlier appearance of age B was detected in the Khalkha Mongols from Bayan-Lig and the Khotons from Tarialan, while there was a later appearance in Bat-Ulziy, Zhargalant, and Khalkhgol, which is typical of populations with an optimal pace of ontogeny.
The interval between points A and B, the time of accumulation of age changes, also varies in duration (see Table 1). It is lengthy in Bat-Ulziy, Zhargalant, and Khalkhgol, where in some cases in females (max = 26 years) it approaches the world maximum typical for centenarian populations. In the Bayan-Lig population (Gobi zone), this interval is minimal among all Mongolian populations studied. If this population is excluded, the Khalkha Mongols in general differ from other populations in having a higher duration for this interval (see Table 2). The Khotons, both males and females, display intermediate values of this variable.
Table 1. Main sample statistics of the OSSEO variables in the studied populations
|
Population, aimag |
Sex |
n 1 |
Age, years (min–max) |
А |
В |
В – А |
n 2 |
a |
b |
|
Uver-Khangay, 1986 |
Male |
97 |
23–78 |
28 |
46 |
18 |
75 |
–6.25 |
0.218 |
|
Female |
99 |
18–72 |
31 |
55! |
24 |
53 |
–2.99 |
0.133 |
|
|
Bayankhongor, 1987 |
Male |
64 |
20–80 |
26 |
38 |
12 |
50 |
–5.67 |
0.269 |
|
Female |
79 |
19–73 |
24 |
35 |
11 |
68 |
–8.32 |
0.336 |
|
|
Khubsugul, 1988 |
Male |
76 |
22–73 |
26 |
41 |
15 |
63 |
–5.76 |
0.239 |
|
Female |
79 |
19–72 |
28 |
54 |
26 |
47 |
–2.47 |
0.125 |
|
|
Eastern, 1990 |
Male |
61 |
19–65 |
30 |
52 |
22 |
34 |
–7.50 |
0.251 |
|
Female |
81 |
18–66 |
32 |
51 |
19 |
42 |
–4.22 |
0.138 |
|
|
Ubsunur (Khotons), 1989 |
Male |
39 |
18–63 |
28 |
45 |
17 |
31 |
–9.55 |
0.365 |
|
Female |
56 |
19–54 |
27 |
41 |
14 |
25 |
–7.96 |
0.301 |
Note . n 1 – total size of the sample; n 2 – size of the “standard sample”, from the point A (the empirical age of the beginning of age-related changes in a population) to the age of 60; a – constant term of the linear regression equation in standard samples; b – coefficient of thelinear regression equation describing the rate of age-related changes in standard samples.
Table 2. Reference values of the OSSEO variables in 46 rural populations
|
Variable |
Males |
Females |
||||
|
Min |
Max |
М |
Min |
Max |
М |
|
|
А |
21.0 |
41.0 |
27.3 |
21.0 |
39.0 |
28.5 |
|
В |
28.0 |
57.0 |
41.1 |
31.0 |
66.0 |
43.7 |
|
В – А |
3.0 |
30.0 |
13.9 |
6.0 |
30.0 |
15.2 |
|
b * |
0.148 |
0.816 |
0.360 |
0.084 |
0.784 |
0.366 |
*See the note to the Table 1.
The rate of accumulation of age-related changes in the hand bones was assessed using regression analysis. The results of the analysis (Fig. 1) show that among all populations from Mongolia employed in this study, males and females of two groups (the Khalkha Mongols from Bayan-Lig (Gobi desert) and the Khotons from Tarialan) exhibit relatively high rates (Batsevich, Yasina, 2000). As compared to the previously studied populations from the former USSR, Israel, and Sinai Peninsula, all Mongolian populations display low to medium rates of skeletal aging (cf. Tables 1 and 2).
Thus, the adult population of Mongolia can be divided into two contrasting groups of populations in terms of their age-related osteomorphic status. The populations displaying an advantageous (delayed) rate of aging inhabit the central, northern, and eastern regions of the country (Khalkha Mongols of the Uver-Khangay, Khubsugul, and Eastern aimags). The adaptively disadvantageous, accelerated tempos of aging of the hand bones, for reasons still unclear, were observed in the Khalkha Mongols from the South (Bayankhongor aimag). Certain biological features of the latter (total body size, temporal stability of body morphology, decelerated maturation of children and adolescents, etc.) do not differ from other Khalkha populations. An influence from climatic, geographical, and geochemical environmental factors of the southern desert, that are different from the typical environment for Mongolian groups, can be hypothesized.
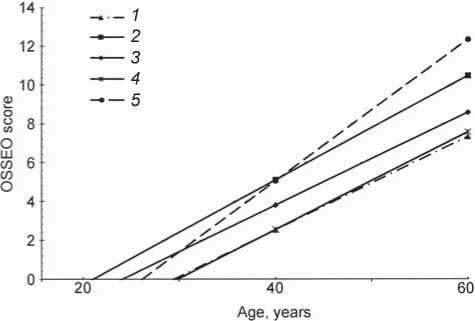
А
Fig. 1. Rates of age-related changes of the hand skeleton.
A – males; B – females.
1 – Bat-Ulziy and Burd; 2 – Bayan-Lig; 3 – Zhargalant; 4 – Khalkhgol; 5 – Tarialan (Khotons).
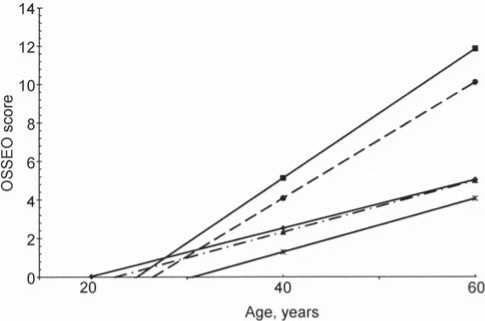
B
The Khotons of the western Ubsurun aimag display an accelerated tempo of ontogeny as well. This population was forced to migrate some 10– 12 generations ago, and changed not only their environment but also their subsistence strategy. Thus it can be reasonably suggested that adaptation in this group is not complete, and the high rate of age-related changes in the hand skeleton is a manifestation of that disadaptation. Similar examples of differences in the pace of skeletal aging between native groups and recent migrants in other regions were described in our previous studies (Batsevich et al., 2009; Batsevich, Kobyliansky, Yasina, 2013).
Age at menarche
The pace of ontogeny of females in adolescence can be assessed based on the age at puberty. The mean age at menarche and its dynamics during long time periods are an indicator of historical and social changes, and the health and well-being of the population (Lehmann, Scheffler, Hermanussen, 2010). Data on the age at menarche were collected in three Khalkha populations (excluding Uver-Khangay aimag) and in one of the Khotons. A preliminary exploration of these data has shown that this variable does not differ significantly among females of those populations. In this connection, pooled data for all the Mongolian females was used further in inter-group comparisons of age and temporal dynamics of sexual maturation rate.
The mean age at menarche of the Mongolian females was 15.8 years (n = 188, σ = 1.23). Its temporal changes have been undirected (Fig. 2, A), which is confirmed statistically (the p-values are shown in the figure). The tempos of sexual maturation in the Khoton females have been changing substantially during the same period of time (Fig. 2, B). The mean age at menarche was the highest in the 1940s (app. 18 years), and decreased during the following three decades. In the 1970s, it averaged 16 years, approaching the values typical for the Khalkha females. The mean age at menarche for the whole studied time span was 16.8 years (n = 64, σ = 1.18). The direct changes of this variable evidence active adaptive processes in this population due to changes in environment.
Temporal dynamics of anthropometric variables
For the study of temporal change of anthropometric variables, we employed a set of metric traits including main lengths and transverse skeletal dimensions of the body segments that were previously shown to change as a result of growth acceleration and the secular trend: lengths of body, torso and arm, and shoulder width. Each of these variables exhibits a specific growth trajectory. The variation of stature and torso length in long-term perspective can be affected by both secular and age-related factors. This issue was addressed in detail in our previous study (Batsevich, Yasina, 2015). But the shoulder width and arm length variation is not substantially affected by age-related changes after reaching maturity.
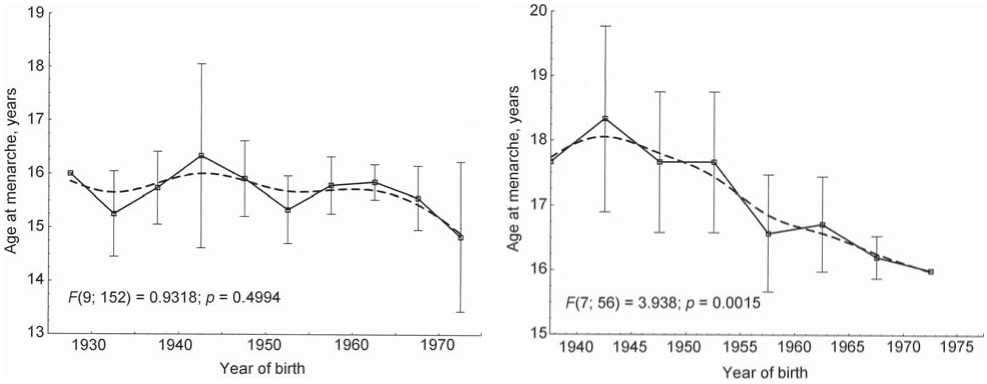
А
B
Fig. 2. Temporal dynamics of the age at menarche of the Khalkha ( A ) and Khoton ( B ) females.
In order to assess the direction and intensity of temporal changes in the anthropometric variables, correlation coefficients between their values and the birth dates of all individuals were calculated (Table 3). There were no significant associations found at the level of single populations in any of the Khalkha Mongolian groups, neither in males nor in females. Thus, it can be reasonably assumed that the somatic status of the rural Mongols was stable between the 1930s and the 1970s. In the Khotons, in the same time span, statistically significant changes in stature and shoulder width were detected in females, and in torso length in males. The increase in these and some other anthropometric variables is slight and does not usually reach the level of statistical significance. The growth acceleration and secular trend is
Table 3. The association between anthropometric variables and birth date in the Khalkha Mongolian and Khoton populations
|
Population, aimag |
Sex |
n |
Stature |
Shoulder width |
Torso length |
Arm length |
||||
|
Correlation coefficient |
p |
Correlation coefficient |
p |
Correlation coefficient |
p |
Correlation coefficient |
p |
|||
|
Uver-Khangay, 1986 |
Male |
141 |
–0.083 |
0.32 |
0.134 |
0.122 |
0.019 |
0.825 |
–0.103 |
0.221 |
|
Female |
152 |
0.102 |
0.21 |
0.049 |
0.547 |
0.012 |
0.880 |
0.073 |
0.371 |
|
|
Bayankhongor, 1987 |
Male |
54 |
–0.006 |
0.96 |
–0.228 |
0.091 |
0.020 |
0.882 |
0.007 |
0.958 |
|
Female |
71 |
0.118 |
0.31 |
0.134 |
0.249 |
0.187 |
0.109 |
0.113 |
0.327 |
|
|
Khubsugul, 1988 |
Male |
75 |
0.090 |
0.42 |
–0.106 |
0.342 |
0.026 |
0.815 |
0.093 |
0.405 |
|
Female |
98 |
0.109 |
0.27 |
–0.148 |
0.131 |
0.069 |
0.483 |
–0.053 |
0.591 |
|
|
Dornod, 1990 |
Male |
56 |
–0.147 |
0.25 |
0.058 |
0.650 |
–0.142 |
0.262 |
–0.131 |
0.301 |
|
Female |
78 |
0.058 |
0.60 |
0.044 |
0.695 |
–0.104 |
0.349 |
–0.115 |
0.303 |
|
|
Ubsunur (Khotons), |
Male |
48 |
0.079 |
0.56 |
0.087 |
0.503 |
0.273 |
0.033 |
0.212 |
0.100 |
|
1989 |
Female |
86 |
0.214 |
0.04 |
0.277 |
0.008 |
0.149 |
0.157 |
0.119 |
0.262 |
Note . Statistically significant coefficients are given in bold.
more pronounced in females, while in males it is commonly found in the form of tendencies. A similar picture of temporal changes in body morphology, accompanying social and economic changes, was previously observed in the Bashkirs. The observed morphological differences between the two sexes are probably related to the gender role differences typical of the ethnic culture (Ibid.).
It was previously demonstrated by T.P. Chizhikova and N.S. Smirnova (2011) that the intergenerational change in stature in a number of populations (Russians, Abkhazians, Chuvash, Bashkirs) was determined by the level of stability of social and cultural conditions. Weakly pronounced or absent changes in body morphology are found when a tradition way of life is kept. A similar association between skeletal maturation and aging rates on one hand and the stability of ecological conditions on the other hand was observed previously (Batsevich et al., 2009).
Another way of assessing temporal changes in body morphology is to compare the results of researches carried out in various years in the past. The data for stature from the literature are
Table 4. Published data on stature of the adults of rural populations of the Khalkha Mongols studied in various years
|
Source |
Population, aimag |
|||||||||||
|
tn О о ^ |
5 5 о О |
о с о -С с го го СП |
ГО с го -С ^ ф ZD |
О) ф 5 |
го с го го < |
с го О) СП |
го "с ф о |
о о о со |
о О го ш |
о го °? ф 5 |
с ф го ш |
|
|
Males |
||||||||||||
|
(Talko-Gryntsevich, 1902) |
161.8 (36) |
|||||||||||
|
(Vlcek, 1965) |
164.8 (59) |
|||||||||||
|
(Nyamdorj, 1988) |
165.1 (170) |
165.4 * (427) |
||||||||||
|
(Zolotareva, 1990) |
164.8 (92) |
164.3 (105) |
166.1 (106) |
166.4 (76) |
164.9 (95) |
164.2 (104) |
165.0 (93) |
164.6 (68) |
165.1 (99) |
|||
|
(Tumen, 1992) |
165.9 (50) |
165.9 (35) |
163.7 (53) |
162.6 (73) |
||||||||
|
(Antropoekologiya Tsentralnoy Azii, 2005) |
164.6 (54) |
165.5 (69) |
165.7 (75) |
165.2 (56) |
||||||||
|
Females |
||||||||||||
|
(Vlcek, 1965) |
151.3 (49) |
|||||||||||
|
(Nyamdorj, 1988) |
154.9 (120) |
153.9 * (707) |
||||||||||
|
(Tumen, 1992) |
153.4 (76) |
153.7 (43) |
151.2 (66) |
153.8 (72) |
||||||||
|
(Antropoekologiya Tsentralnoy Azii, 2005) |
154.1 (71) |
153.5 (70) |
154.4 (98) |
154.5 (78) |
||||||||
Note. Sample size is given in parentheses.
*Pooled data for South Gobi and East Gobi aimags.
presented in Table 4. Taking into account the dates of publications and age distribution of the samples, the temporal dynamic of stature in Mongolia during the last hundred years can be traced (though some adjustment on the number and geographic distribution of the studied samples is certainly needed). The mean stature of the Khalkha Mongolian males ranges from 161.8 to 166.1 cm irrespective of the population’s location and year of study (Table 4). The minimal value of this variable, 161.8 cm, was reported in the earliest study by Talko-Gryntsevich (1902), carried out in Urga (present day Ulan-Bator) in the middle of the 1890s. The author of that study emphasized that his sample employed young males retaining a potential for further growth. The delayed biological maturation of the Mongols in the early 20th century, which could last until 25–26 years of age, was also reported by Rokhlin (1936: 162). The mean stature of the Khalkha Mongolian females varies from 151.2 to 154.9 cm (Table 4), which, like that of the Mongolian males, fits into the range of values typical for related populations.
Notably, stature in other ethnic groups of Mongolia studied during the 1980s was similar to that in the Khalkha populations. D. Tumen, in his dissertation (1992), reports stature values in 22 ethnic and subethnic groups. No substantial differences in total body size relating to linguistic, ethnic, or geographic differences among the samples were detected. Thus, it can be reasonably hypothesized that the similarity in some anthropometric variables among Mongolian populations is due to the influence of similar and stable environmental conditions.
The temporal dynamic of a number of anthropometric and also physiological variables is a result of the change in the pace of ontogeny due to the influence of changing social and economic factors. During 50 years of research in Mongolia, accelerated growth was only observed in children and adolescents of Ulan-Bator city (Tsolmon, 1994; Otgon, Efimova, Korytov, 2009; Uranchimeg, 2011; and others). An exploration of the data collected in various years demonstrates a gradual increase in total body size in all age cohorts. But the tempo of these changes is not the same in different periods. For instance, a steep acceleration of growth and its differentiation according to social status was observed in Ulan-Bator at the turn of the 21st century (Uranchimeg, 2011). The complex influence of urban stresses on the biological features of citizens is well studied, and the consequences of such influence are broadly similar in various parts of the world (Nikityuk, 1989; Godina, Miklashevskaya, 1990; Purundzhan, 1990; Bogin, 1988). In this respect, the urban dwellers from Mongolia repeat the path of the population of other countries that experienced an early urbanization.
The research on growth and development of rural children of 8 aimags and 11 somons, carried out in 1987–1998, revealed a similar rate of maturation in various climatic and geographic zones of Mongolia (Antropoekologiya Tsentralnoy Azii, 2005: 127–153; Erdene, 2007). The definite body size of male and female adolescents of older age cohort do not differ from that of adults of previous generations. This suggests that the secular trend and growth acceleration are absent in these populations in the studied period.
Conclusions
The results of the present study show that the adult rural Khalkha Mongolian populations, leading a traditional way of life, retain the optimal adaptive balance with their environment. No temporal changes in the age at menarche or total body size in both sexes were observed in the studied samples. Our X-ray data revealed a delayed rate of aging of the hand bones in adults. The migrant Khotons, who have changed their traditional culture and economy, exhibit manifestations of an accelerated pace of ontogeny as compared to the adapted native population. A similar situation was found in migrant groups in Transcaucasia and Israel (Batsevich et al., 2009; Batsevich, Kobyliansky, Yasina, 2013). In the Khotons, an increase in stature and transverse bodydimensions was observed as well. Their morphology is similar to that of other local Mongolian ethnic groups of the Ubsunur aimag: the Bayads and Derbets. Apparently, the Khoton population is gradually approaching the biological adaptive norm typical of the native population of this region.
Judging by a number of biological variables explored in this study, the Khalkha Mongolian populations described here can be considered well-adapted to the complex of environmental factors. In terms of their level of adaptation, they are most similar to the centenarian groups of the Caucasus. Such a high level of adaptation of the modern Mongolian population can be explained in most cases by the longterm socio-economic stability of their habitat. The situation observed in the Khotons seems to confirm the above-mentioned conclusion.
Acknowledgement
This study was supported by the Russian Science Foundation (Project No. 14-50-00029).
