Секреторная функция желудка при действии мышечной нагрузки в условиях фармакологической блокады М-холино- и -адренорецепторов

Автор: Кузнецов Александр Павлович, Смелышева Лада Николаевна, Архипова Ольга Алексеевна, Московкин Алексей Сергеевич, Сажина Нина Витальевна
Журнал: Человек. Спорт. Медицина @hsm-susu
Рубрика: Физиология
Статья в выпуске: S1 т.19, 2019 года.
Бесплатный доступ
Цель: изучение влияния действия мышечной нагрузки на секреторную функцию желудка у человека в условиях частичной фармакологической блокады М-холино- и β-адренорецепторов при ацидификации двенадцатиперстной кишки. Материалы и методы. У 22 лиц мужского пола 18-23 лет методом гастрального и гастродуоденального зондирования исследовалась секреторная функция желудка в покое при действии 30-минутной велоэргометрической нагрузки объемом 36 900 кгм в сочетании с частичной фармакологической блокадой М-холинорецепторов (1,5 мг/кг массы тела) и β-адренорецепторов (0,6 мг обзидана на кг массы тела). Результаты. Блокада М-холинорецепторов вызывала в условиях мышечного покоя выраженное снижение валового содержания электролитов (p ˂ 0,01), ферментов (p ˂ 0,001), HCl (p ˂ 0,001) и протеолитической активности (p ˂ 0,001) в желудочном соке. Сочетанное действие атропина и физической нагрузки в условиях ацидификации двенадцатиперстной кишки еще более выраженно тормозило объем секрета до 49,9 ± 11,1 %, HCl до 29,7 ± 9,4 %, пепсиногена до 19,8 ± 7,4 % (p ˂ 0,001). При блокаде β-адренорецепторов секреторная функция желудка в условиях базальной секреции и ацидификации двенадцатиперстной кишки достоверно снижалась. При использовании в качестве стимулятора желудочной секреции 10 % отвара сухой капусты объем возрастал до 141,1 ± 12,2 %, дебит HCl до 186,3 ± 21,9 %, пепсиноген до 188,5 ± 20,4 %. Аналогичная картина наблюдалась при выполнении велоэргометрической нагрузки в условиях блокады β-адренорецепторов. Заключение. При действии мышечной нагрузки на секреторный аппарат желудка существенно возрастает роль симпатического отдела вегетативной нервной системы. При ацидификации двенадцатиперстной кишки в условиях блокады β-адренорецепторов тормозный эффект усиливается, а при стимуляции капустным отваром секреторной функции желудка отмечается ее усиление.
Секреторная функция желудка, м-холино- и β-адренорецепторы, велоэргометрическая нагрузка
Короткий адрес: https://sciup.org/147233555
IDR: 147233555 | УДК: 612.34 | DOI: 10.14529/hsm19s107
Stomach secretory function under muscle load in the conditions of pharmacological blocade of M-cholinoreceptors and -adrenoceptors
Aim. The article deals with establishing the effect of muscle load on the stomach secretory function in the conditions of partial pharmacological blockade of M-cholinoreceptors and β-adrenoceptors in patients with duodenal acidification. Materials and methods. 22 males aged 18-23 years participated in the study. The stomach secretory function at rest under a 30-minute cycle ergometer load of 36 900 kgm combined with a partial pharmacological blockade of M-cholinoreceptors (1.5 mg per kg of body mass) and β-adrenoceptors (0.6 mg obsidan per kg of body mass) was studied using gastric and gastroduodenal intubation. Results. The blockade of M-cholinoreceptors at muscle rest provoked a noticeable decrease in the total content of electrolytes (p
Текст научной статьи Секреторная функция желудка при действии мышечной нагрузки в условиях фармакологической блокады М-холино- и -адренорецепторов
Введение. В последние годы наблюдается рост числа работ отечественных и зарубежных исследователей, посвященных регуляции секреторной функции пищеварительных желез [3, 5, 11, 12, 15]. Это в первую очередь обусловлено тем, что сохраняется высокий процент людей с различными заболеваниями пищеварительной системы [1, 13, 14, 17, 18, 19]. Для выявления причин этого явления необходимы знания об участии в регуляции желудочно-кишечного тракта центральной нервной и эндокринной систем, гормонов желудочно-кишечного тракта, циклических нуклеотидов, посредством которых осуществляется влияние практически всех гормонов [2, 4, 7, 16, 20]. Однако следует отметить, что в основном все исследования проводились в условиях относительного мышечного покоя, когда уровень двигательной активности сведен до минимума. В повседневной жизни че- ловек очень часто испытывает значительные мышечные и эмоциональные напряжения. Специалистами спортивной медицины установлено, что у людей, имеющих высокий уровень повседневной двигательной активности, в условиях мышечного покоя наблюдаются специфические особенности регуляторных механизмов, обеспечивающих экономичность функций организма [6, 7]. Особенно проявление регуляторных механизмов наблюдается в процессе выполнения мышечной нагрузки и после ее окончания у людей, адаптированных и неадаптированных к действию физической нагрузки [5, 9]. Все эти изменения накладывают отпечаток на различные функции систем организма, в том числе и на желудочнокишечный тракт [10, 21].
Перед исследователем, который ставит перед собой задачу изучения механизмов регуляции секреторной функции пищеваритель- ных желез, возникает методическая трудность, суть которой состоит в том, что при описании единых нейрогуморальных механизмов регуляции пищеварительных желез он вынужден условно расчленять этот процесс, выделяя в нем нервные и гуморальногормональные механизмы.
Для изучения влияния парасимпатического и симпатического отделов вегетативной нервной системы использовали фармакологическую блокаду М-холинорецепторов (1,5 мг атропина на кг массы тела, подкожно) и β-адре-норецепторов (0,6 мг обзидана на кг массы тела, внутрь). Фармакологические препараты вводились за 15–20 мин до начала зондирования, а при выполнении дозированной нагрузки – за 15–20 мин до ее начала. В качестве дозированной нагрузки использовалась работа на велоэргометре в течение 30 мин общим объемом 36 900 кгм при частоте педалирования 60 об/мин.
Результаты исследования и их обсуждение. Классическими работами И.П. Павлова и многочисленными последующими исследователями было убедительно продемонстрировано, что в регуляции желудочной секреции ведущая роль принадлежит блуждающему нерву [3, 5, 15].
В табл. 1 представлены показатели концентрации электролитов, ферментов и протеолитической активности желудочного сока в условиях мышечного покоя, частичной блокады М-холинорецепторов (атропин 1,5 мг/кг массы тела, подкожно) и при введении атропина с последующим выполнением 30-минутной дозированной велоэргометрической нагрузки объемом 36 900 кгм.
В условиях блокады М-холинорецепторов атропином выявлены существенные изменения в концентрации ферментов и электролитов за исключением достоверного снижения
Таблица 1
Table 1
Влияние атропина и велоэргометрической нагрузки объемом 36 900 кгм на концентрацию электролитов, ферментов и протеолитическую активность желудочного сока (M ± m) (n = 10)
Effect of atropine and a cycle ergometer load of 36 900 kgm on the content of electrolytes, enzymes, and proteolytic activity of gastric juice (M ± m) (n = 10)
|
Исследуемые показатели Parameters |
В покое At rest |
Атропин Atropine |
Атропин + нагрузка Atropine + load |
|||
|
I |
II |
I |
II |
I |
II |
|
|
Хлориды, ммоль/л Chlorides, mmol/l |
94 ± 5,4 |
87 ± 7,7 |
101 ± 9,3 |
105 ± 7,3 |
107 ± 8,4 |
113 ± 9,02* |
|
HCl, мкмоль/л HCl, μmol/l |
30 ± 8,4 |
31 ± 6,0 |
27 ± 8,0 |
11 ± 4,8 |
19 ± 3,2 |
26 ± 3,4 |
|
Натрий, ммоль/л Sodium, mmol/l |
38 ± 3,6 |
41 ± 4,1 |
38 ± 4,3 |
62 ± 7,1 |
32 ± 4,1 |
38 ± 4,4 |
|
Калий, ммоль/л Potassium, mmol/l |
15 ± 0,9 |
13 ± 0,5 |
10 ± 0,5*** |
9 ± 1,1 |
8,2 ± 0,9** |
9 ± 0,91* |
|
Пепсиноген, мкг/мл Pepsinogen, mkg/ml |
345 ± 36 |
390 ± 40 |
214 ± 27** |
129 ± 39 |
211 ± 26** |
178 ± 21*** |
|
Протеолитическая активность, мкг/мл Proteolytic activity, mkg/ml |
90 ± 10,1 |
120 ± 14,4 |
84 ± 11,2* |
45 ± 18 |
35 ± 5,3* |
40 ± 5,8*** |
|
Липаза, ед./мл Lipase, unit/ml |
0,5 ± 0,05 |
0,5 ± 0,05 |
0,55 ± 0,07 |
1,05 ± 0,08 |
0,89 ± 0,1** |
0,7 ± 0,08 |
Примечание. Здесь и в табл. 2 I – базальная секреция, II – стимулированная секреция. В условиях стимулированной секреции в двенадцатиперстную кишку вводился 0,5 % (30 мл) раствор хлористоводородной кислоты. Достоверность по отношению к исходному уровню в покое: * – p ˂ 0,05; ** – p ˂ 0,01; *** – p ˂ 0,001.
Note. Here and in table 2 I – basal secretion, II – stimulated secretion. In the conditions of stimulated secretion, a 0.5 % (30 ml) hydrochloric acid solution was injected into the duodenum. Reliability in relation to the initial level at rest: * – p ˂ 0.05; ** – p ˂ 0.01; *** – p ˂ 0.001.
При выполнении 30-минутной велоэрго-метрической нагрузки в сочетании с блокадой М-холинорецепторов атропином выявили достоверное снижение в базальном секрете протеолитической активности, концентрации пепсиногена и калия. Исключение составили и показатели концентрации липазы в желудочном секрете, которые достоверно (p ˂ 0,01) возрастали по сравнению с показателями в покое. При совместном влиянии велоэргомет-рической нагрузки и атропина в условиях ацидификации двенадцатиперстной кишки раствором хлористоводородной кислоты обнаружили снижение в базальном желудочном секрете протеолитической активности и концентрации пепсиногена и калия. При этом в условиях атропинизации снижение концент- рации пепсиногена и калия были более выражены, чем при сочетанном влиянии мышечной нагрузки и атропина.
Достоверные изменения выявили при блокаде М-холинорецепторов как раздельно, так и в сочетании с действием физической нагрузки в показателях протеолитической активности, дебит-часе ферментов и электролитов в желудочном соке (табл. 2). Фармакологическая блокада М-холинорецепторов вызывала снижение исследуемых показателей желудочной секреции кроме валового выделения липазы. В условиях введения в двенадцатиперстную кишку раствора хлористоводородной кислоты наблюдалось выраженное торможение желудочной секреции. Объем желудочного сока снижался до 49,9 ± 11,1 %, дебит-час пепсиногена – до 19,8 ± 7,4 %, хлористоводородной кислоты – до 29,7 ± 9,4 %, суммарная протеолитическая активность – до 22,9 ± 12,1 %. Не изменялся только один показатель – дебит-час липазы. Очевидно, повышение pH желудочного сока в этих условиях является благоприятным фактором повышения активности липазы, которая наиболее активна в нейтральной среде.
04 CM
ГО Ф
н ю го
|
(ГО 43 я + + я я -я К & о 5 < |
-X- НН |
4Г о" НН 40^ ГН |
-х-оо о" НН |
-X- -X-о" НН о |
* о" НН |
* o^ o" HH ^ o" |
* * ГН НН 00^ |
* * * о" НН °\ |
ГН НН 04 ГН |
|
|
-х- -х-СП ГП НН ID си |
о" НН ГП |
* о" НН оо^ СП |
* -х- о" НН о" |
* -х-гн о" НН |
* o^ o" o" |
* -х-ID СО НН °\ 40" |
о" НН |
* ГН пГ НН 40 ГН |
||
|
Я О S .S 1g с < |
-х- ID НН 00 |
-X- -X-•D о" НН Ог |
о" НН о^ |
ГН о" НН |
СП гн^ о" НН 40^ ГН |
* o" HH ID^ o" |
-х-сп о^ си" НН 40^ оо" |
* НН 00^ ГН |
о" НН |
|
|
гн UH НН ип оо" ID |
ГН о" НН оо^ ГН |
оо Ог о" НН оо^ •D |
о" НН ^ |
сп^ о" НН ГН ГН |
04 o^ o" HH ^ o" |
О^ сп" НН о |
НН 00^ |
4о" НН ГН СП |
||
|
CD _ О GO Й О О ^ m ^ |
оо^ оо" НН оо |
ГН о" НН ГН ГН |
ОО^ о" НН •D |
ГП о" НН СП |
сп^ о" НН ^ си" |
ГН гн^ о" -H |
НН m |
40 04 о" НН ГН |
оо^ 4о" НН |
|
|
1— |
о" НН оо |
40 О^ о" НН ГН |
40^ о" НН |
о" НН ГН СП |
оо ГН о" НН о^ си" |
о" НН |
ГН НН СП СП |
НН оо^ 04" |
НН ГН |
|
|
Я О CD С Id Я (ГО CD П О |
о я § ^ g 0 81 |
к О- |
ч 2 g в и Й и |
^ и и К к |
11 5S g н 45 Щ CZ) |
е 'о g 11 )S СЛ S g § О « л |
я Ы>2 S Щ г. CD 0 ^ CD О ад G О. С рц |
ьР" S ад CD -rt Я <ГО £ .У о Я " Д « 2 К ^ я |
1^ 5J СО CD я .g- е=; и |
|
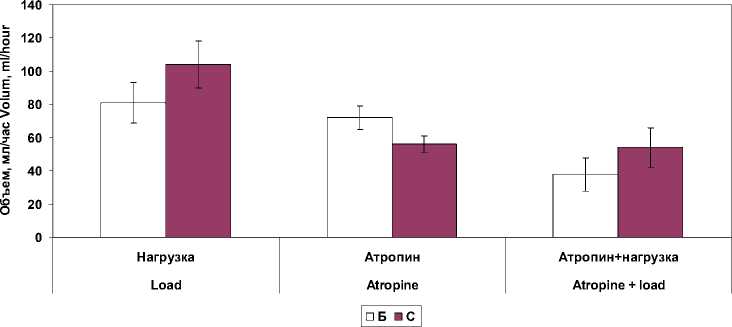
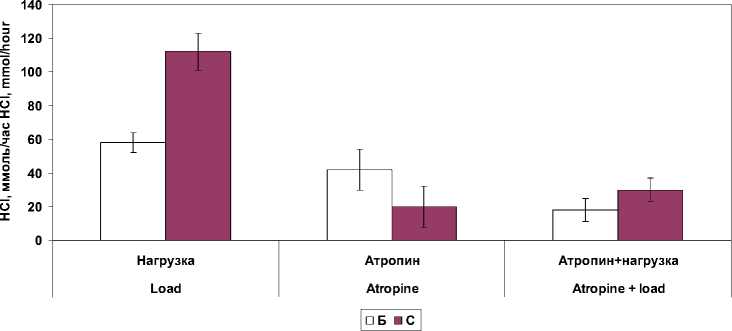
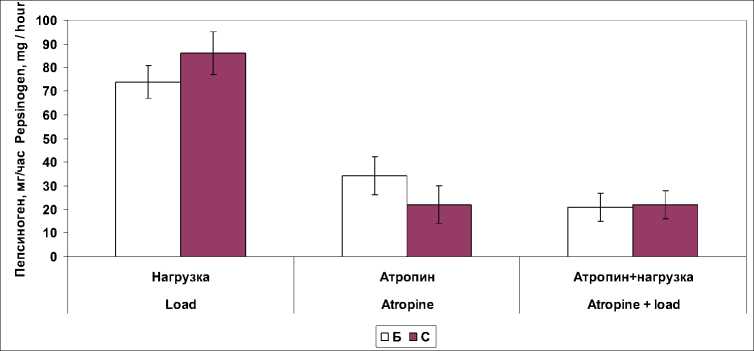
Б – в условиях базальной секреции; С – в условиях ацидификации двенадцатиперстной кишки раствором хлористоводородной кислоты Fig. 1. Stomach secretory function with isolated and combined effect of atropine and a 30-minute physical activity (in% of the baseline level):
B – basal secretion; C – in the conditions of duodenal acidification with hydrochloric acid
пепсиногена – до 70,8 ± 8,9 %, протеолитическая активность желудочного секрета снижалась до 67,6 ± 8,3 %.
В условиях мышечного покоя влияние симпатического отдела вегетативной нервной системы на регуляцию желудочной секреции незначительно, хотя и установлено наличие на мембранах париетальных клеток адренорецепторов. Регулирующие эффекты в основном обеспечиваются через действие гистамина, ацетилхолина и гастрина (В.Т. Ивашкин,
Список литературы Секреторная функция желудка при действии мышечной нагрузки в условиях фармакологической блокады М-холино- и -адренорецепторов
- Ивашкин, В.Т. Метаболическая организация функций желудка / В.Т. Ивашкин. - Л.: Наука, 1981. - 215 с.
- Климов, П.К. Физиология желудка. Механизмы регуляции / П.К. Климов, Г.М. Барашкова. - Л.: Наука, 1991. - 256 с.
- Коротько, Г.Ф. Физиология системы пищеварения / Г.Ф. Коротько. - Краснодар: Изд-во ООО БК «Группа Б», 2009. - 608 с.
- Коротько, Г.Ф. Постпрандиальная секреция поджелудочной железы / Г.Ф. Коротько. - Краснодар: Изд-во «ЭДВИ», 2017. - 115 с.
- Кузнецов, А.П. Желудочно-кишечный тракт и стресс / А.П. Кузнецов, А.В. Речкалов, Л.Н. Смелышева. - Курган: Изд-во Курган. гос. ун-та, 2004. - 254 с.
