Синхронизация охоты и приживляемость эмбрионов у коров черно-пестрой породы при гормональном индуцировании суперовуляции

Автор: Рамирес Родригес Р., Сергеев Н.И., Порфирьев И.А.
Журнал: Сельскохозяйственная биология @agrobiology
Рубрика: Экологические проблемы в животноводстве
Статья в выпуске: 6 т.39, 2004 года.
Бесплатный доступ
На коровах и телках черно-пестрой породы исследовали эффективность использования гормональных препаратов с целью синхронизации охоты. Оценивали влияние различных факторов на оплодотворяемость в период индуцированной охоты и при суперовуляции, а также приживляемость эмбрионов при пересадке. Рассматривается возможность овариостимуляции для ускоренного завершения послеродового периода и восстановления репродуктивной функции коров - доноров эмбрионов.
Короткий адрес: https://sciup.org/142133024
IDR: 142133024 | УДК: 636.02:636.082.451:57.089.32:577.17
Synchronization of hunting and embryo survival in cows of the black-and-white breed at the hormonal induction of superovulation
The authors investigated the efficiency of the use of hormonal preparations for hunting synchronization in cows and heifers of the Black-and-White breed. The influence of different factors on the fertilization during inducible hunting and superovulation and also on the embryo survival after transplantation was estimated. It was shown, that gonadotropins in combination with prostaglandin need to be applied for synchronization of sexual cycle in donors and recipients. The use of synthetic gonadoliberin corrects the possible disturbances in ovulation, that permit to obtain the embryos from high productive cows in more early period after calving. At that the variation of ovulation number, fertility of ovule, number and stage of development of embryos ready for transplantation in superovulated cows-donors depend on duration of hunting period. The optimal concentrations of progesterone and estradiol in cow's blood under the apply of prostaglandins are 3,1 ng/ml and 28,9 pg/ml, respectively. The possibility of ovary stimulation for accelerated completion of post-natal period and recovery of reproductive function of cows-donors of embryos is considered.
Текст научной статьи Синхронизация охоты и приживляемость эмбрионов у коров черно-пестрой породы при гормональном индуцировании суперовуляции
У крупного рогатого скота лимитирующим фактором успешного применения метода трансплантации эмбрионов, способствующего интенсификации воспроизводства, служат нарушения репродуктивной функции коров (1-5). Существенным ограничением трансплантации эмбрионов является также низкая оплодотворяемость, отклонения в развитии яйцеклеток после оплодотворения и высокое число погибших эмбрионов. Результативность пересадки повышается по мере усовершенствования методов трансплантации и устранения воздействия негативных внешних факторов. Однако причины эмбриональной смертности изучены недостаточно.
Эффективность трансплантации эмбрионов в практике селекции молочного скота во многом зависит от правильного подбора коров-доноров по способности производить достаточное число качественных эмбрионов после гормональной обработки и искусственного осеменения. Разработка объективных тестов прогнозирования эмбрио-продуктивности доноров имеет важное теоретическое и практическое значение для улучшения воспроизводства высокопродуктивных коров. На результаты трансплантации эмбрионов оказывают также влияние физиологические особенности доноров и реципиентов и, кроме того, квалификация специалистов, осуществляющих процедуры (6-8).
В задачу нашей работы входила синхронизация охоты у коров-доноров и телок-реципиентов посредством индуцирования суперовуляции при использовании препаратов простагландина, фолликулостимулирующего гормона и гона-долиберинов, оценка эффективности комбинированного применения биологически активных соединений, показателей суперовуляции в зависимости от продолжительности охоты и содержания гормонов в крови, а также приживляемо-сти эмбрионов при пересадке телкам-реципиентам.
Методика. В качестве доноров эмбрионов использовали высокопродуктивных племенных коров и телок черно-пестрой породы из опытного хозяйства «Щапово», ферм «Кузенево» и «Лукошкино» экспериментального хозяйства «Кленово-Чегодаево» Всероссийского НИИ животноводства (ВИЖ). Реципиентами служили 417 телок черно-пестрой породы случного возраста живой массой 350-380 кг.
Для синхронизации охоты доноров и реципиентов и стимуляции овариальной активности применяли синтетические препараты простагландина F2α (PgF2α) и эстрофан как раздельно, так и в сочетании с гипофизарными гормонами — фолликулостимулирующим (ФСГ), фоллитропином и гонадотропином сыворотки жеребых кобыл (ГСЖК), а также гонадолиберинами — диригистра- ном и сурфагоном. Использование сурфагона способствовало коррекции гормонального равновесия, а также ослаблению или устранению последствий нарушения последнего.
Для сравнительной оценки эффективности комбинированного применения эстрофана в сочетании с ГСЖК и сурфагоном было сформировано три группы животных (табл. 1). Перед введением гормонов проводили 2-кратную инъекцию тетравита (по 10 мл) по общепринятой схеме: в день обработки и через 48 ч после применения PgF2 α . Животным II и III групп гормоны вводили при наличии выраженного желтого тела в одном из яичников, коровы III группы дополнительно получали сурфагон в дозе 15 мкг перед первым осеменением.
1. Доза и время введения различных гормональных препаратов при индуцировании охоты у коров-доноров черно-пестрой породы (схема первого опыта)
|
Группа животных |
n |
Препарат (доза) |
Время введения препарата, сутки полового цикла |
|
I (контроль) |
30 |
Эстрофан (500 мкг) |
10-12-е |
|
II |
30 |
Эстрофан (500 мкг) + ГСЖК (500 ИЕ) |
7-9-е и 13-15-е |
|
III |
30 |
Эстрофан (500 мкг) + ГСЖК (500 ИЕ) + сурфагон (15 мкг) |
7-9-е и 13-15-е |
Для стимуляции овуляции у коров-доноров после отела воздействовали препаратами фоллитропин, ГСЖК в сочетании с сурфагоном, а также PgF2 α (табл. 2). Наличие желтого тела и развитие фолликула выявляли в ходе клинического обследования. Гонадолиберины и лютеинизирующий гормон вводили через 48 ч после применения PgF2 α .
2. Доза и время введения различных гормональных препаратов при стимуляции послеродовой овариальной активности у коров черно-пестрой породы (схема второго опыта)
|
Группа животных |
n |
Препарат (доза) |
Время введения препарата, сутки после отела |
|
I (контроль) |
50 |
– |
– |
|
II |
30 |
Фоллитропин (500 ИЕ) |
5-е |
|
Сурфагон (15 мкг) |
12-е |
||
|
III |
50 |
ГСЖК (1000 ИЕ) |
5-е |
|
Сурфагон (15 мкг) |
12-е |
||
|
IV |
30 |
Фоллитропин (1000 ИЕ) + PgF 2 α (500 мкг) |
12-е |
|
Сурфагон (15 мкг) |
14-е |
||
|
V |
50 |
ГСЖК (1000 ИЕ) |
11-е |
|
PgF 2 α (500 мкг) |
13-е |
||
|
Сурфагон (15 мкг) |
15-е |
На 7-е сут после первого осеменения эмбрионы извлекали, оценивали их качество и проводили подготовку к пересадке по методикам ВИЖ.
Результаты. И н д у ц и р о в а н и е о х о т ы у к о р о в - д о- н о р о в. По данным первого опыта, общее количество животных, пришедших в охоту, составляло в I, II и III группах соответственно 90,0; 86,6 и 90,0 %, в том числе соответственно 70,0; 66,7 и 83,4 % через 48 ч после применения препаратов (табл. 3). Следовательно, при совместной инъекции эстрофана и гонадотропина эффективность синхронизации охоты повышается, а гонадолиберин корректирует возможные нарушения овуляции.
3. Оценка синхронизации охоты у коров-доноров черно-пестрой породы при комбинированном введении гормонов
|
Показатель |
Группа животных |
|||||
|
I (контроль) |
II |
III |
||||
|
n |
% |
n |
% |
n |
% |
|
|
Общее количество животных |
30 |
100 |
30 |
100 |
30 |
100 |
|
Из них пришло в охоту: |
27 |
90,0 |
26 |
86,7 |
27 |
90,0 |
|
в том числе через: |
||||||
|
24 ч |
3 |
10,0 |
2 |
6,7 |
2 |
6,7 |
|
48 ч |
21 |
70,0 |
20 |
66,7 |
25 |
83,3 |
|
72 ч |
3 |
10,0 |
4 |
13,3 |
– |
– |
Не проявляли признаков охоты
Пригодно к индуцированию суперовуляции
3 10,0
90,0
4 13,3 3 10,0
24 80,0 27 90,0
П р и м е ч а н и е. Схема первого опыта представлена в таблице 1.
Восстановление в послеродовой период воспроизводительной функции у коров является одной из серьезных проблем зооветеринарной службы. Чаще всего нарушения этой функции возникают при задержании последа, атонии матки или заболеваниях яичников (персистентное желтое тело, гипофункция, киста и др.). Для профилактики послеродовых осложнений используют различные лекарственные и гормональные препараты, но надежно гарантировать восстановление воспроизводительной функции за короткое время не удается, так как применяемые препараты не создают тот гормональный фон, который необходим для активации полового цикла.
С т и м у л я ц и я о в у л я ц и и у к о р о в - д о н о р о в п р и и с п о л ь з о в ан и и ф о л л и кул о с ти м ул ир у ю -щ е г о г о р м о н а в с о ч е т а н и и с р и л и з и н г - г о р-м о н а м и. По данным второго опыта, первая овуляция после отела у коров II-V групп наступала на 2,0-3,7 сут раньше, чем в контроле (табл. 4). Более выраженная стимуляция овуляции выявлена у животных IV и V групп, оплодотворяемость которых после первого осеменения была выше, чем в контроле, — соответственно 62,00-63,33 и 52,00 %; индекс осеменения составлял 1,65; продолжительность сервис-периода — 72,367,5 сут (против соответственно 1,95 и 85,0 сут в контроле). Следовательно, совместное введение фоллитропина, сурфагона, ГСЖК и PgF2 α в различных комбинациях вызывает суперовуляцию, что позволяет использовать корову в качестве донора в более ранние сроки после отела по сравнению с интактными особями.
4. Показатели овариальной активности коров-доноров черно-пестрой породы при введении различных гормональных препаратов в разные сроки после отела
|
Группа животных |
Число животных, имеющих в яичниках: |
Время после отела, сут |
Количество коров по времени первой овуляции после отела |
||||
|
менее 15 сут |
более 15 сут |
||||||
|
желтое тело |
желтое тело и фолликул или только фолликул |
n |
% |
n |
% |
||
|
I (контроль) |
27 |
23 |
20,1 ± 3,4 |
10 |
20,0 |
40 |
80,0 |
|
II |
12 |
18 |
18,1 ± 4,0 |
6 |
20,0 |
24 |
80,0 |
|
III |
27 |
23 |
17,4 ± 3,4 |
19 |
38,0 |
31 |
62,0 |
|
IV |
16 |
14 |
16,4 ± 2,1 |
11 |
36,7 |
19 |
63,3 |
|
V |
26 |
24 |
16,4 ± 3,1 |
20 |
40,0 |
30 |
60,0 |
П р и м е ч а н и е. Схема второго опыта представлена в таблице 2.
П р о д о л ж и т е л ь н о с т ь о х о т ы, с т а д и я р а з -в и т и я и к а ч е с т в о э м б р и о н о в. Известно, что овуляция индуцированных фолликулов затягивается по времени, например, на 7-8-е сут после осеменения обнаруживают эмбрионы на разных стадиях развития. После индуцирования суперовуляции фоллитропином в синхронизированный (I группа) и спонтанный (II группа) циклы ( n = 51) у некоторых коров-доноров охота продолжалась на 2-е и 3-и сут (табл. 5). Животных в I и II группах осеменяли соответственно 3 и 4 раза с интервалом 12 ч на протяжении всей охоты, что способствовало увеличению числа овуляций соответственно на 1,2 и 1,5 (на одного донора). Наивысшая оп-лодотворяемость яйцеклеток отмечена при 3-кратном осеменении; при продолжительности охоты 36 ч и 4-кратном осеменении этот показатель снижался.
5. Оплодотворяемость яйцеклеток у коров-доноров черно-пестрой породы в зависимости от продолжительности охоты и кратности осеменения
|
Продолжительность охоты, ч |
Количество коров |
Кратность осеменения |
Число овуляций на одного донора |
Оплодотворяемость яйцеклеток, % |
|
|
n |
% |
||||
|
С и н |
х р о н и з и р о в а н н ы й п о л о в о й ц и к л (I группа) |
||||
|
12 |
4 |
20 |
2 |
7,5 |
85,0 |
|
24 |
10 |
50 |
3 |
3,0 |
93,2 |
|
36 |
6 |
30 |
4 |
8,7 |
72,7 |
|
С п о н т а н н ы й п о л о в о й |
ц и к л (II группа) |
||||
|
12 |
8 |
25,8 |
2 |
6,9 |
85,0 |
|
24 |
16 |
51,6 |
3 |
7,3 |
93,6 |
|
36 |
7 |
22,6 |
4 |
8,4 |
84,6 |
Уменьшение продолжительности охоты до 12 ч и увеличение до 36 ч не оказывало влияния на число клеточных структур в расчете на одного донора, но число пригодных для трансплантации эмбрионов снижалось (табл. 6). При этом количество пригодных для трансплантации эмбрионов на близких стадиях развития (поздние морулы и ранние бластоцисты) составляло 49,1 -51,9 %, а на стадии ранней и поздней морулы и поздней бластоцисты — снижалось до 11,1-23,1 %.
6. Показатели качества эмбрионов в зависимости от стадии их развития и продолжительности охоты у коров-доноров черно-пестрой породы
|
Продолжительность охоты |
Число доноров |
Стадия развития эмбриона |
Число клеточных структур |
Количество эмбрионов, пригодных для трансплантации |
|||
|
общее |
на одного донора |
шт. |
% |
на одного донора, шт. |
|||
|
Менее 12 ч |
11 |
Поздние морулы и ранние бластоцисты |
57 |
5,2 ± 1,3 |
28 |
49,1 |
2,6 ± 0,6 |
|
3 |
Ранние и поздние морулы и поздние бластоцисты |
26 |
8,7 ± 1,1 |
6 |
23,1 |
2,0 ± 0,5 |
|
|
12-24 ч |
23 |
Поздние морулы и ранние бластоцисты |
128 |
5,6 ± 1,4 |
111 |
86,7 |
4,8 ± 0,6 |
|
5 |
Ранние и поздние морулы и поздние бластоцисты |
43 |
8,6 ± 1,6 |
10 |
23,3 |
2,0 ± 0,3 |
|
|
Более 24 ч |
11 |
Поздние морулы и ранние бластоцисты |
52 |
4,7 ± 0,9 |
27 |
51,9 |
2,5 ± 0,4 |
|
10 |
Ранние и поздние морулы и поздние бластоцисты |
72 |
7,2 ± 1,2 |
8 |
11,1 |
0,8 ± 0,2 |
|
Большинство эмбрионов от доноров, у которых продолжительность охоты составляла 12-24 ч, находилось на стадии поздней морулы и ранней бластоцисты, причем количество эмбрионов, пригодных для трансплантации, было почти в 2 раза больше, чем у остальных коров. Следовательно, у коров-доноров при суперовуляции стадия развития и число пригодных для трансплантации эмбрионов зависят от продолжительности охоты.
С и н х р о н и з а ц и я о х о т ы и м о р ф о ф у н к - ц и о н а л ь н о е с о с т о я н и е я и ч н и к о в. Для синхронизации охоты коровам ( n = 90) вводили эстрофан в дозе 500 мкг на разных стадиях полового цикла: 7-9-, 10-12-и 13-15-е сут (при наличии в одном из яичников желтого тела) — соответственно I, II и III группы. Перед введением препарата отбирали пробы крови для определения концентрации прогестерона и эстрадиола. Большинство животных II группы (10-12-е сут полового цикла) пришло в охоту через 48 ч после инъекции эстрофана (табл. 7). До 24 ч и после 48 ч пришли в охоту по три особи, признаки охоты не отмечены у 10 % коров. После введения эстрофана у всех пришедших в охоту животных (90 %) на 8-10-е сут индуцированного цикла было выявлено желтое тело при отсутствии пальпируемого фолликула, что служило основным показателем индукции суперовуляции.
7. Оценка синхронизации охоты у коров-доноров черно-пестрой породы в зависимости от стадии полового цикла и исходной концентрации гормонов в крови
|
Группа животных |
Время начала охоты после введения гормона, ч |
Количество животных, пришедших в охоту |
Исходная концентрация в крови |
Количество животных, пригодных к суперовуляции |
|||
|
n |
% |
прогестерона, нг/мл |
эстрадиола, пг/мл |
n |
% |
||
|
I ( n = 30) |
24 |
4 |
13,3 |
0,5 ± 0,3 |
18,0 ± 1,4 |
– |
– |
|
48 |
5 |
16,7 |
1,8 ± 0,2 |
19,6 ± 0,1 |
4 |
13,3 |
|
|
72 |
10 |
33,3 |
0,8 ± 0,1 |
20,6 ± 0,1 |
1 |
3,3 |
|
|
II ( n = 30) |
24 |
3 |
10,0 |
1,9 ± 0,2 |
31,9 ± 2,0 |
3 |
10,0 |
|
48 |
21 |
70,0 |
3,1 ± 0,3 |
28,9 ± 1,7 |
21 |
70,0 |
|
|
72 |
3 |
10,0 |
1,5 ± 0,3 |
26,0 ± 2,0 |
3 |
10,0 |
|
|
III ( n = 30) |
24 |
4 |
13,3 |
0,7 ± 0,3 |
30,1 ± 3,0 |
2 |
6,6 |
|
48 |
11 |
36,7 |
2,0 ± 0,2 |
27,0 ± 2,1 |
9 |
30,0 |
|
|
72 |
4 |
13,3 |
0,7 ± 0,1 |
21,6 ± 2,3 |
– |
– |
|
|
Всего |
65 |
72,2 |
– |
– |
43 |
47,7 |
|
П р и м е ч а н и е. Животные I, II и III групп находились соответственно на 7-9-, 10-12- и 13-15-х сут полового цикла.
Время прихода в охоту коров в разных группах значительно варьировало; не проявляли признаков охоты 26,7 % животных (см. табл. 7). В I и III группах доля коров, пригодных для использования в качестве доноров, составляла соответственно 16,6 и 36,6 %, то есть в 5 и 2,5 раза меньше, чем во II группе. Концентрация прогестерона в крови животных I и III групп перед введением эстрофана составляла 0,5-0,7 нг/мл, что свидетельствует об отсутствии возможности быстрого лизиса желтого тела под действием экзогенного PgF2 α . Однако такое функциональное состояние характерно для соответствующих периодов цикла, так как только у 16,7 и 36,7 % коров (соответственно I и III группы) концентрация прогестерона (соответственно 1,8 и 2,0 нг/мл) обеспечивала приход в охоту, что позволяло стимулировать суперовуляцию.
У коров II группы приход в охоту через 48 ч после введения эстрофана характеризовался высокой концентрацией прогестерона и низкой — эстрадиола в середине лютеальной стадии цикла; до 24 ч — наоборот; до 72 ч — низкой концентрацией как эстрадиола, так и прогестерона.
У животных I и III групп приход в охоту через 48 ч и наибольшее число пригодных для последующего использования доноров отмечены при близкой к средней (для соответствующих стадий цикла) концентрации эстрадиола и достаточно высокой — прогестерона, что вызывало секреторную активность желтого тела. При увеличении или уменьшении концентрации эстрадиола и низкой концентрации прогестерона наблюдалось варьирование сроков прихода в охоту у самок I и III групп либо отсутствие охоты у таковых во всех трех группах. Следовательно, общее снижение результативности синхронизации охоты у коров с 7-х по 15-е сут спонтанного цикла при наличии хорошо выраженного желтого тела в одном из яичников происходит при недостаточной концентрации гормонов в крови при введении эстрофана на 7-9-е и 13-15-е сут полового цикла. При этом подавляющее большинство коров, пригодных для индуцированной суперовуляции, приходило в охоту через 48 ч после обработки. Применение PgF 2 α вызывало лизис желтого тела в течение нескольких часов, а затем овуляцию фолликула. При этом большинство особей приходило в охоту через 48 ч после инъекции препарата. PgF2 α оказывал лютелитическое действие в период с 5-6-х по 15-16-е сут спонтанного полового цикла. Однако введение простагландина в период до 10-х и после 12-х сут цикла даже при наличии в яичниках типичного желтого тела не всегда сопровождалось полноценным и синхронным лизисом последнего у некоторых коров, что приводило к варьированию сроков прихода в охоту или отсутствию последней.
Концентрация гормонов в крови перед синхронизацией половых циклов коров-доноров препаратами простагландина служит диагностическим тестом: оптимальной концентрацией прогестерона и эстрадиола можно считать соответственно 3,1 нг/мл и 28,9 пг/мл.
Суперовуляторную реакцию яичников и качество эмбрионов в зависимости от концентрации гормонов в крови доноров оценивали не менее чем через 2 мес после отела у коров как со спонтанным (I группа), так и индуцированным РgF2 α + ГСЖК (II группа) половым циклом. Для этого животным вводили фоллитропин + РgF2 α в течение 4 сут, начиная с 10-х сут цикла. Через 48 ч после последней инъекции коров в период охоты осеменяли ректоцервикальным методом с интервалом в 12 ч замороженно-оттаянной спермой одного и того же производителя в двойной дозе. Перед инъекцией определяли концентрацию прогестерона и эстрадиола в крови. При оценке влияния исходной концентрации прогестерона и эстрадиола на суперовуляторную реакцию яичников и качество эмбрионов были выделены четыре степени реакции суперовуляции по числу желтых тел и извлеченных эмбрионов, пригодных для пересадки: первая — соответственно 12 и 9 шт. (и более), вторая — 8-11 и 5-8 шт., третья — 4-7 и 1-4 шт., четвертая — 3 шт. (и менее) и отсутствие пригодных для пересадки эмбрионов (табл. 8). У коров II группы каждой степени суперовуляции соответствовала определенная исходная концентрация гормонов в крови (особенно прогестерона). Так, при повышении концентрации прогестерона число овуляций и нормальных эмбрионов увеличивалось соответственно от 3 до 12 и от 0 до 9. Аналогичная корреляция наблюдалась и у животных I группы, которым вводили гормоны при спонтанном цикле. При этом исходная концентрация прогестерона в крови животных II группы была выше, чем у таковых I группы, что, по-видимому, связано с увеличением массы лютеальной ткани при используемой схеме синхронизации.
Результативность гормонального индуцирования суперовуляции обусловлена оптимальной активностью желтого тела при концентрации прогестерона в крови доноров от 2,9 до 4,0 нг/мл. Отступления от выявленной тенденции связаны с индивидуальными особенностями животных. Выявлена корреляция между числом овулировавших и неовулировавших фолликулов при минимальной концентрации прогестерона. Зависимости числа клеточных структур от исходной концентрации эстрадиола не наблюдалось. Следовательно, определение исходной концентрации прогестерона в крови коров-доноров позволит в определенной степени прогнозировать эмбриопродуктивность отбираемых животных.
8. Оценка суперовуляторной реакции яичников и качества эмбрионов в зависимости от исходной концентрации гормонов в крови коров-доноров черно-пестрой породы и синхронизации полового цикла
|
Показатель |
Ι группа (спонтанный цикл) |
ΙΙ группа (синхронизированный цикл, РgF2 α + ГСЖК) |
||||
|
n |
Концентрация |
n |
Концентрация |
|||
|
прогесте рона, нг/мл |
эстрадиола, пг/мл |
прогестерона, нг/мл |
эстрадиола, пг/мл |
|||
|
Число овуляций (желтых тел): 12 15 3,1 ± 1,2 29,7 ± 2,6 26 4,0 ± 1,7 23,5 ± 1,6 8-11 25 2,9 ± 0,3 24,7 ± 2,8 40 3,2 ± 1,7 19,8 ± 1,4 4-7 30 2,6 ± 0,3 29,0 ± 2,0 30 2,9 ± 0,7 18,3 ± 1,2 3 12 1,2 ± 0,3 26,7 ± 2,3 16 1,2 ± 0,8 19,1 ± 1,2 Число эмбрионов, пригодных для трансплантации: 9 8 8,4 ± 0,2 33,6 ± 5,0 12 9,0 ± 1,9 21,5 ± 5,7 5-8 15 2,9 ± 0,4 23,8 ± 2,3 23 3,4 ± 0,7 20,6 ± 1,5 1-4 15 2,5 ± 0,4 26,0 ± 2,4 17 2,8 ± 0,3 17,3 ± 1,2 |
||||||
Продолжительность охоты и качество эмбрион о в п р и в в е д е н и и с и н т е т и ч е с к и х г о н а д о л и б е р и н о в и л ю т е и н и з и р у ю щ е г о г о р м о н а. Для оценки суперовуляции, индуцируемой введением в течение 4 сут ФСГ + РgF2α, было сформировано четыре группы коров: I (контроль) — не вводили синтетические гонадолиберины; II, III и IV — через 48 ч после введения простагландина перед первым осеменением вводили соответственно диригистран (Чехия) в дозе 15 мкг, сурфагон в дозе 15 мкг и овогон-ТИО (1000 ИЕ). Осеменение во II-IV группах коров проводили 2кратно с интервалом 12 ч, в контроле — через 48 ч после введения простагландина на протяжении всего периода охоты с интервалом 12 ч. Продолжительность охоты у коров I, II, III и IV групп составляла соответственно 28,8; 20,7; 18,0 и 19,0 ч (табл. 9).
Хотя клеточных структур в расчете на одного донора было больше у животных I группы, число пригодных для трансплантации эмбрионов у коров II-IV групп было выше. При этом следует отметить снижение числа дегенери-рованных эмбрионов и неоплодотворенных яйцеклеток, доля которых в контроле более чем в 2 раза превышала таковую в опыте. Доля эмбрионов, пригодных для трансплантации, у животных II, III и IV групп составляла соответственно 74,6; 74,6 и 81,1 % против 47,3 % в контроле.
9. Оценка продолжительности охоты и качества эмбрионов при введении коровам-донорам черно-пестрой породы синтетических гонадолиберинов и лютеинизирующего гормона
|
Показатель |
Группа животных |
|||
|
Ι (контроль) |
ΙΙ (диригистран) |
ΙΙΙ (сурфагон) |
IV (овогон-ТИО) |
|
|
Количество обработанных коров: гол. |
13 |
11 |
20 |
13 |
|
% |
100 |
100 |
100 |
100 |
|
Количество положительных доноров: гол. |
13 |
11 |
20 |
13 |
|
% |
100 |
100 |
100 |
100 |
|
Продолжительность охоты, ч |
28,8 ± 2,3 |
20,7 ± 2,2 |
18,0 ± 2,4 |
19,0 ± 2,4 |
|
Получено клеточных структур: шт. (всего) |
93 |
67 |
134 |
74 |
|
% |
100 |
100 |
100 |
100 |
|
шт. (на одного донора) |
7,1 ± 0,6 |
6,1 ± 1,0 |
6,7 ± 0,7 |
5,7 ± 0,8 |
|
Количество пригодных для трансплантации эмбрионов: шт. (всего) |
44 |
50 |
100 |
60 |
|
% |
47,3 |
74,6 |
74,6 |
81,1 |
|
шт. (на одного донора) |
3,4 ± 0,8 |
4,5 ± 0,6 |
5,0 ± 0,7 |
4,6 ± 0,5 |
|
Количество дегенерированных эмбрионов: шт. (всего) |
26 |
8 |
18 |
7 |
|
% |
28,0 |
11,9 |
13,4 |
9,5 |
|
шт. (на одного донора) |
2,0 ± 0,8 |
0,7 ± 0,2 |
0,9 ± 0,4 |
0,5 ± 0,5 |
|
Количество неоплодотворенных яйцеклеток: шт. (всего) |
23 |
9 |
16 |
7 |
|
% |
24,7 |
13,5 |
11,9 |
9,5 |
|
шт. (на одного донора) |
1,8 ± 0,7 |
0,8 ± 0,3 |
0,9 ± 0,3 |
0,5 ± 0,1 |
Следовательно, включение синтетических гонадолиберинов или лютеинизирующего гормона в схему индуцирования суперовуляции позволяет корректировать возможные нарушения овуляции и повышает число эмбрионов, пригодных для трансплантации.
Для выбора оптимального срока осеменения при суперовуляции у 19 коров-доноров определяли время между инъекцией простагландина, началом эструса и максимальной концентрацией лютеинизирующего гормона:
Препарат
ФСГ + РgF 2 α α
Фоллитропин + РgF 2 α α
n Инъекция простагландина— Инъекция простагландина—максимальная начало эструса, ч концентрация лютеинизирующего гормона, ч
9 41,3 ± 2,3 48,5 ± 3,0
10 42,0 ± 5,5 48,1 ± 3,9
Одновременно проводили регулярную ректальную пальпацию яичников сразу после инъекции простагландина на протяжении всего периода эструса. Через 41-42 ч после инъекции РgF 2 α наступала течка, а через 48 ч концентрация лютеинизирующего гормона достигала максимума, после чего через 20-24 ч происходила овуляция. Поэтому время осеменения необходимо выбирать так, чтобы спермии, попавшие в половые органы самки, завершали процесс капаци-тации к началу овуляции, то есть первое осеменение следует проводить через 48 ч после инъекции простагландина.
К о н ц е н т р а ц и я г о р м о н о в в к р о в и к о- р о в - д о н о р о в п р и с т и м у л я ц и и о в у л я ц и и р и л и- з и н г - г о р м о н а м и. Причиной неудовлетворительной суперовуляции, индуцированной в первый половой цикл после родов, являлась низкая концентрация прогестерона в крови перед введением фоллитропина + + РgF 2 α вследствие функциональной неполноценности желтого тела, что в свою очередь вызвано незавершенной инволюцией матки и общей послеотель-ной перестройкой (рис, а). Гормональное индуцирование активности яичников в по-слеотельный период позволило получить удовлетворительные показатели суперовуляции (три-пять эмбрионов на одного донора) в индуцированный половой цикл и положительные результаты (более пяти эмбрионов) в последовавший спонтанный половой цикл (см. рис., б, в).
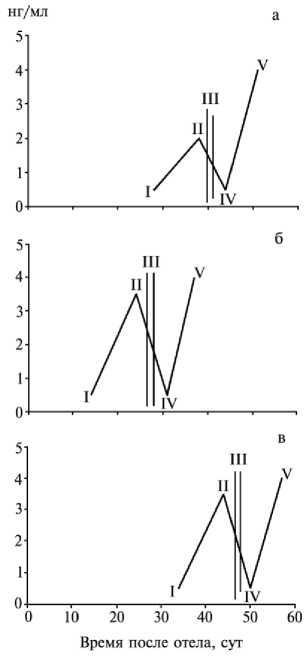
Концентрация прогестерона (нг/мл) в крови коров-доноров черно-пестрой породы при индуцировании суперовуляции в послеродовой период посредством введения фоллитропина + простагландина (PgF 2 α ) в первый спонтанный (а), индуцированный (б) и спонтанный после индуцированного (в) половые циклы: I — охота; II — инъекция гормонов; III — овуляция; IV — осеменение; V — извлечение эмбрионов.
Считается, что индукция суперовуляции в первый после отела цикл оказывается безуспешной. Наши исследования свидетельствуют о возможности положительной суперовуляторной реакции. Направленная стимуляция активности функциональной деятельности яичников приводила к активации восстановления репродуктивной функции в целом, о чем свидетельствует концентрация прогестерона, приближающаяся к среднестатистическим показателям, которые являются нормой для соответствующего периода спонтанных циклов после окончательного завершения послеотельного периода. Пригодность коров для индуцирования суперовуляции определялась концентрацией прогестерона в крови, причем показанием к введению гормонов служит неуклонное повышение последней.
Непосредственной причиной нарушения суперовуляции являются преждевременная или поздняя овуляции, а также отсутствие таковой б о льшего или меньшего числа фолликулов и сочетание этих процессов, что в свою очередь обусловливает послеовуляторные сдвиги в гормональном балансе. Своевременно зафиксированный гормональный дисбаланс позволяет нормализовать оплодотворение и развитие эмбрионов посредством введения гонадолиберина или лютеинизирующего гормона. Следовательно, по показателям суперовуляции, вызванной в спонтанный после индуцированного цикл, можно корректировать овариальную функцию в ранний период после отела. Достигаемая за счет послеродовой овариостимуляции стабилизация функциональной деятельности яичников ускоряет восстановительные процессы в матке.
П р и ж и в л я е м о с т ь э м б р и о н о в в зависимос-ти от действия ра з л и ч н ы х ф а к т о р о в. На основании 274
пересадок телкам-реципиентам свежеполученных эмбрионов определяли степень приживляемости последних в зависимости от показателей суперовуляции коров-доноров и качества вымываемых эмбрионов. Эмбрионы извлекали за 2-3 ч до пересадки у 38 коров-доноров, которым в течение 4 сут вводили ФСГ в количестве 45 мг (инъекции по убывающим дозам). Через 3 мес после пересадки ректально оценивали стельность реципиентов. При увеличении степени суперовуляции достоверно снижалось число эмбрионов хорошего и отличного качества (табл. 10). При этом приживляемость эмбрионов при высоком числе овуляций была меньше, чем при низком и среднем.
10. Приживляемость эмбрионов у коров черно-пестрой породы в зависимости от степени суперовуляции и показателей качества зародышей
|
Показатель |
Число овуляций |
|
0-4 5-8 9-12 13-16 |
|
|
Число коров-доноров Получено эмбрионов, шт.: всего на одного донора Доля эмбрионов от числа овуляций, % |
6 7 13 12 17 52 86 119 2,8 7,4 6,6 9,9 70,0 ± 11,1 93,0 ± 3,5 55,0 ± 5,4 62,0 ± 4,5 |
Количество извлеченных клеточных структур, %: эмбрионы:
Таким образом, при индуцировании охоты с целью синхронизации полового цикла у коров-доноров и телок-реципиентов следует вводить гонадотропины в сочетании с простагландином, что способствует стимуляции овариальной активности в течение 7-15 сут цикла. Синтетический гонадолиберин корректирует возможные нарушения овуляции, что существенно повышает эффективность синхронизации охоты. Введение в схемы овариостимуляции препаратов гонадолиберина на 14-15-е сут после отела стимулирует первую овуляцию и регулярность половых циклов, что позволяет получать эмбрионы от высокопродуктивных лактирующих коров в более ранние сроки и максимально приблизить межотельный период у коров-доноров к экономически обоснованному, а также может служить лечебно-профилактическим средством при отклонениях функциональной деятельности яичников и эндометрия, способствуя ускоренному восстановлению репродуктивной функции коров после индуцирования суперовуляции. При этом варьирование числа овуляций, оплодотво-ряемость яйцеклеток, количество и стадия развития эмбрионов, пригодных для трансплантации, у суперовулировавших коров-доноров зависят от продолжительности охоты. Так, оплодотворяемость яйцеклеток повышается при увеличении продолжительности охоты от 12 до 24 ч и кратности осеменения до 3. Диагностическим тестом перед проведением синхронизации половых циклов у коров-доноров при введении препаратов простагландина может служить концентрация гормонов в крови: оптимальная концентрация прогестерона — 3,1 нг/мл, эстрадиола — 28,9 пг/мл. Определение исходной концентрации прогестерона в крови коров-доноров позволяет также оценить овариальную активность после отела и уточнить данные клинического контроля перед синхронизацией охоты и индуцированием суперовуляции, что дает возможность прогнозировать эмбриопродуктивность отбираемых животных. Эффективность индуцирования суперовуляции существенно возрастает при введении синтетических гонадолиберинов или лютеинизирующего гормона; первое осеменение следует проводить через 48 ч после инъекции простагландина. Наибольшее количество эмбрионов отличного и хорошего качества и достаточно высокие показатели приживляемости отмечены у коров, эмбриопродуктивность которых составляла 5-8 овуляций. При трансплантации замороженно-оттаянных эмбрионов наиболее высокой приживляемостью характеризуются эмбрионы, полученные от коров-доноров с низкой и средней эмбриопродуктивностью. При трансплантации особо ценных эмбрионов следует использовать телок-реципиентов, у которых в яичнике диагностировано отличное или хорошее функциональное состояние желтого тела.
Л И Т Е Р А Т У Р А
-
1. Х и л к е в и ч С., Т я п у г и н Е. Экономические аспекты трансплантации эмбрионов в молочном скотоводстве. Молочное и мясное скотоводство, 1992, 5-6: 16-17.
-
2. Б а л к о в о й И.И., И н о з е м ц е в В.П., Х р и с т о ф о р о в В.Н. и др. Милк-терапия мастита у коров. Ветеринария, 1997, 3: 46-47.
-
3. С е р г е е в Н.И., О в ч и н н и к о в А.В., С м ы с л о в а Н.И. Взаимодействие между донором, эмбрионом и реципиентом при нехирургической трансплантации. Животноводство, 1987, 6: 52-54.
-
4. Ш и п и л о в В.С. Состояние и перспективы развития ветеринарного акушерства и гинекологии. Ветеринария, 1991, 7: 3-6.
-
5. Р е ш е т н и к о в а Н.М., М о р о з Т.А., М а л и н о в с к и й А.М. и др. Проблема интенсификации воспроизводства в племенном скотоводстве. В сб.: Современные аспекты селекции, биотехнологии, информатизации в племенном животноводстве. М., 1997: 121-130.
-
6. С а м о д е л к и н А.Г., Г а в р и к о в А.М., С е р г е е в Н.И. Трансплантация эмбрионов в мясном скотоводстве. М., 1996.
-
7. Э р н с т Л.К., С е р г е е в Н.И. Трансплантация эмбрионов сельскохозяйственных животных. М., 1989.
-
8. A k e L o p e z J.R., A l f a r o G a m b o a M.E., H o l y L. Respuesta superovulatoria en ganado bos indicus y bos taurus bajo condiciones tropicales, y efecto del desarrollo de la calidad del embri o n sobre el porcentaje de la gestaci o n. Veterinaria de M e xico, l995,26,3: 189-193.
Российский университет дружбы народов, Поступила в редакцию 6
117198, Москва, ул. Миклухо-Маклая, 8/1; февраля 2002 года
Всероссийский государственный НИИ животноводства, Московская обл., п/о Дубровицы
SY NCHRONIZATION OF HUNTING AND EMBRYO SURVIVAL
IN COWS OF THE BLACK-AND-WHITE BREED AT THE HORMONAL INDUCTION OF SUPEROVULATION
R. Ramires Rodriges, N.I. Sergeev, I.A. Porfir’ev
S u m m a r y
The authors investigated the efficiency of the use of hormonal preparations for hunting synchronization in cows and heifers of the Black-and-White breed. The influence of different factors on the fertilization during inducible hunting and superovulation and also on the embryo survival after transplantation was estimated. It was shown, that gonadotropins in combination with prostaglandin need to be applied for synchronization of sexual cycle in donors and recipients. The use of synthetic gonadoliberin corrects the possible disturbances in ovulation, that permit to obtain the embryos from high productive cows in more early period after calving. At that the variation of ovulation number, fertility of ovule, number and stage of development of embryos ready for transplantation in superovulated cows-donors depend on duration of hunting period. The optimal concentrations of progesterone and estradiol in cow’s blood under the apply of prostaglandins are 3,1 ng/ml and 28,9 pg/ml, respectively. The possibility of ovary stimulation for accelerated completion of post-natal period and recovery of reproductive function of cows-donors of embryos is considered .
