Синхронность в сроках половой активности самок и самцов вуалевого песца в условиях звероводческого хозяйства

Автор: Баранов В.А., Халилова Г.Х., Шумилина Н.Н.
Статья в выпуске: 3 т.251, 2022 года.
Бесплатный доступ
Звероводство, как и все отрасли сельского хозяйства, на настоящий момент требует оптимизации непосредственно в отношении подхода к физиологическим циклам зверей. Отрасль относительно молода, доместикация зверей ещё продолжается, и моноцикличность зверей имеет свои определённые нюансы, что приводит к необходимости подстраиваться под условия рынка. В статье представлены результаты проведенных в республике Татарстан (ЗАО «Бирюли») исследований по изучению влияния даты начала и характера эструса на показатели воспроизводства самок песцов. Изучена работа самцов по результатам воспроизводства покрытых ими самок. Получены данные, которые свидетельствуют о том, что лучшие показатели воспроизводства у самок, покрытых самцами в ранние сроки эструса. Знания в отношении физиологии зверей и возможность контролирования сроков эструса самок и половой активности самцов, могут помочь посмотреть на возможности производства, более комплексно.
Звероводство, песец, гон, эструс, воспроизводство
Короткий адрес: https://sciup.org/142235689
IDR: 142235689 | УДК: 591.16: | DOI: 10.31588/2413_4201_1883_3_251_28
Synchronicity in the timing of sexual activity of females and males of the veiled arctic fox in the conditions of fur farming
Animal husbandry, like all agriculture branches, currently requires optimization directly in relation to the approach to the physiological cycles of animals. The industry is relatively young, the domestication of animals is still ongoing, and the semelparity of animals has its own certain nuances, which leads to the need to adapt to market conditions. The article presents the results of the researches conducted in the Republic of Tatarstan (CJSC «Biryuli») on the study of the influence of the start date and the nature of estrus on the reproduction indicators of female Arctic foxes. The work of males based on the results of reproduction of females mated by him was studied. The data obtained indicate that the best reproduction rates are in females mated by males in the early stages of estrus. Knowledge about the physiology of animals and the ability to control the timing of female estrus and sexual activity of males, can help to look at the possibilities of production, more comprehensively.
Текст научной статьи Синхронность в сроках половой активности самок и самцов вуалевого песца в условиях звероводческого хозяйства
Звероводство, как и все отрасли сельского хозяйства, на настоящий момент требует оптимизации непосредственно в отношении подхода к физиологическим циклам зверей. Отрасль относительно молода, доместикация зверей ещё продолжается, и моноцикличность зверей имеет свои определённые нюансы, что приводит к необходимости подстраиваться под условия рынка. В статье представлены результаты проведенных в республике Татарстан (ЗАО «Бирюли») исследований по изучению влияния даты начала и характера эструса на показатели воспроизводства самок песцов. Изучена работа самцов по результатам воспроизводства покрытых ими самок. Получены данные, которые свидетельствуют о том, что лучшие показатели воспроизводства у самок, покрытых самцами в ранние сроки эструса. Знания в отношении физиологии зверей и возможности контролировать сроки эструса самок и половой активности самцов, могут помочь посмотреть на возможности производства более комплексно.
В настоящих условиях рынка пушнины, у производителей нет возможности вести производство по пути, который не наращивает интенсивность, не оптимизируется из срока в срок. Так, международная конференция по клеточному пушному звероводству осветила актуальные вопросы, в том числе и выступление Хеннинга Отте Хансена, на тему: «Мировое производство и торговля мехом в 2015-2016 гг.» и одним из вопросов было влияние изменений цен шкурок зверей на производительные обороты хозяйств. Было бы ошибочно полагаться на выводы учёного, если бы эта картина не повторялась вновь и вновь, как после Копенгагенского аукциона 2018 года и взлёта цены, так и после падения спроса в 2016 году и снижения оборотов зверохозяйств в предыдущий год. Поэтому хозяйства, следующие всеобщим тенденциям, остаются в проигрыше, тогда как постоянное получение стабильного количества пушнины хорошего качества, возможно, станет путём к успеху [1, 5].
Оптимизация цикла производства пушнины не останавливается на создании лучших условий и наличия лишь самых здоровых зверей, а также на обеспечении новыми прогрессивными и рациональными типами кормления [8, 9].
Возможность выявления периода наиболее высокой воспроизводительной способности самок, способствует минимизации проведения различных внутрихозяйственных манипуляций работниками зверофермы в период гона, отсюда следует уменьшение трудозатрат. Результаты покрытия самок самцами зависят, как правило, как от активности самца, так и от срока плодотворного покрытия, поскольку есть данные результатов исследований свидетельствующие о помётах с большим количеством щенков к моменту забоя [7, 12, 13, 14]. В свою очередь, селекция по размеру песцов показала хорошие результаты, таким образом, этот признак можно увеличивать не только путем селекции по длине тела, но и сбитости туловища. Однако, на фермах Финляндии, плодовитость самок отрицательно коррелировала с размером щенков к периоду убоя [2, 4, 6, 8, 10]. А также стоит учитывать возраст песцов при содержании в племенном ядре, так как отмечено, что по результатам покрытий самок самцами в возрасте 3-4 лет количество плодотворных спариваний было меньше, чем самцами с возрастом более 4 лет [3, 11].
Материал и методы исследований. Исследования были проведены на песцовой ферме в ЗАО «Бирюли» Высокогорского района Республики Татарстан, в условиях кафедры мелкого животноводства Московской ГАВМ и Б – МВА имени К.И. Скрябина и кафедры технологии животноводства и зоогигиены Казанской ГАВМ им. Н.Э. Баумана.
С целью углубиться в изучаемую проблему 99 самок вуалевого песца были разделены на три группы согласно срокам эструса – с ранним, средним и с поздним сроками проявления признаков течки. В отличие от стандартных примеров определения сроков у самок, а именно: ранний – в случае прихода в гон до 13 марта, средний – срок прихода в гон до 31 марта, поздний – не дальше 10 апреля, с целью облегчения подсчёта были взяты иные временные рамки, отличающиеся от стандарта на несколько дней. Такой вариант расчёта позволяет смотреть в будущее и размышлять о сдвиге сроков на более ранние, для удобства проведения гона, а также, отмечая экономическую выгоду, от более старших по возрасту зверей к моменту забоя, по причине раннего срока рождения.
Характер эструса у песцов был учтен у 56 особей. У самок первого поколения эструс учитывали на дату первого спаривания второго сезона размножения, тогда как у самок второго и третьего поколения дата первого спаривания второго сезона размножения соответствовала срокам их матерей. Эструс стабильный, в случае, если со второго сезона размножения самка приходит в гон в те же сроки с максимальным отклонением в ±10 дней. Эструс начинают определять со второго сезона, по причине стабилизации сроков гона лишь во второй сезон, в первые же сроки могут быть более ранние или поздние, то есть сроки могут быть изменены. Статичный эструс – при отклонении сроков гона со второго сезона размножения в разрыве ±15 дней, а динамичный при отклонении последующих сроков гона на ±30 дней.
Самцы (37 гол.) изучались в соответствии с данными, представленными в племенных картах, для анализа были взяты результаты гона и щенения самок, покрытых каждым из них в течение гона, причем дата покрытия фиксирована в карточках.
Статистическую обработку исследуемого материала проводили на персональном компьютере с помощью программ Microsoft Office.
Результат исследований. По воспроизводительным качествам самки вуалевого песца были распределены по трём группам; самки со средним сроком прихода в гон представляют большинство (37 гол.), с немного увеличенным периодом беременности (51,75±0,06), по сравнению с самками с ранними сроками начала гона (50,58±0,2), но при этом большим количеством мёртвых щенков, при увеличенном количестве покрытий в сравнении с двумя другими группами. Хотелось бы уточнить, что период прихода в гон с 20 марта по 4 апреля хоть и даёт достаточно хорошие результаты по продуктивности самок, но в эти сроки покрывать самок не желательно, так как эти щенки не вырастут к срокам забоя также, как те щенки, что рождены раньше, а, как известно, убой происходит при полной зрелости волосяного покрова, медлить и ждать роста щенков нет возможности. Исходя из этого, хозяйство теряет в результатах зачета по качеству.
Таблица 1 – Воспроизводительные качества самок вуалевого песца, в зависимости от сроков гона
|
Показатель |
Ед. изм. |
Группа зверей по датам гона |
|||
|
16.02.-3.03. |
4.03.-19.03. |
20.03.-4.04. |
|||
|
Поголовье самок в родительском стаде |
n |
31 |
37 |
6 |
|
|
Продолжительность беременности |
дни |
50,58±0,2** |
51,75±0,06 |
52,00±0,35 |
|
|
Родилось щенков |
живых |
гол. |
13,20±0,10 |
12,50±0,07 |
13,70±0,32 |
|
мертвых |
гол. |
0,80±0,02 |
1,46±0,05 |
0,83±0,15 |
|
|
всего |
гол. |
14,00±0,08 |
14,05±0,05 |
14,50±0,33 |
|
|
Количество покрытий |
3,32±0,02 |
3,54±0,02 |
3,50±0,17 |
||
** Р>0,99
Таблица 2 – Воспроизводительные качества самок вуалевого песца, в зависимости от характера эструса
|
Показатель |
Ед. изм |
Характер эструса |
|||
|
стабильный |
статичный |
динамичный |
|||
|
Поголовье самок в родительском стаде |
n |
6 |
7 |
46 |
|
|
Продолжительность беременности |
дни |
47,6±3,3 |
51,8±1,0 |
51,6±0,6 |
|
|
Родилось щенков |
живых |
гол. |
14,5±1,62 |
13,28±1,12 |
12,83±0,32 |
|
мертвых |
гол. |
2,3±0,4 |
1,5±0,33 |
1,92±0,36 |
|
|
всего |
гол. |
15,66±1,12 |
14,14±1,14 |
14,09±0,29 |
|
|
Количество покрытий |
3,3±0,2 |
3,3±0,3 |
3,5±0,1 |
||
В таблице 2 приводятся данные большого разрыва по срокам гона из года в год, а именно ±30 дней у 46 самок, хотя показатели их воспроизводства весьма достойны (кроме малозначительного увеличения количеств покрытий, а именно 3,5±0,1, по сравнению с 3,3±0,2 у самок со стабильным эструсом), но как было описано выше, щенки не достигнут максимального роста к моменту убоя, так как от года к году самка может прийти в гон в разные сроки с колебанием в месяц. А разрывы в сроках гона не только ухудшают показатели производства, но и затрудняют работу с самками, а именно, выявление начала охоты, и продлевает сроки гона, так как колебание прихода самок в гон существенно велико по срокам.
Анализ классификации воспроизводительных способностей самок песцов в зависимости от срока и характера эструса, наглядно показывает, что у щенков самок с динамичным характером эструса, вне зависимости от сроков, имеется высокий процент смертности щенков (ранний – 9,72, средний – 7,94), хоть генеральная совокупность исчисляется из большого количества самок. В то же время самок со стабильным и статичным эструсом гораздо меньше, но их результаты перекликаются, однако недостаточное количество данных не позволяет говорить о выявлении закономерности в воспроизводственных показателях между самками разных эструсов и одинакового периода гона и наоборот. Но данные таблицы 3 дают возможность предположить, что высокий процент смертности может быть связан со снижением активности самцов, по причине увеличения светового дня, так как именно солнце сигнализирует о смене времён года, что и является одной из причин завершения гона у моноэстричных хищников.
В отношении самцов провести анализ, основанный на изучении родословной при помощи карточек, явилось невозможным, по причине отсутствия данных об их родителях. Поэтому нами были взяты данные действующих самцов и результаты за последний гон.
Определяя периоды наибольшей активности самцов, были соотнесены даты совершения покрытий самцами в стаде с результатами щенения, а именно с количеством полученных от них щенков. Используя в работе данные о сроках наибольшей активности самцов (Рисунок 1) возможно максимально исключить прохолостение самок по вине низкой потентности самцов.
Таблица 3 – Воспроизводительные способности самок песцов в зависимости от срока и характера эструса
|
Характер эструса |
n |
>Я я я Я о О я я о ^ |
>я я н о я о я я о ^ |
я я Я ч Я о Я я ю |
я я н о н я С |
X я . Я я * 2 Я й н о о и Я Я о |
я я Я Н о Е X О Я й я |
я о Н о о о И |
X |
|
Стабильный ранний |
5 |
14 |
3,4±0,27 |
47,0±0,44 |
5,0±0,01 |
70 |
7 |
77 |
9,09 |
|
Стабильный средний |
1 |
2 |
3,0±0,36 |
51,0±1,00 |
8,5±0,01 |
17 |
0 |
17 |
0 |
|
Статичный ранний |
6 |
15 |
3,3±0,36 |
51,3±1,00 |
5,3±0,01 |
79 |
6 |
85 |
7,06 |
|
Статичный средний |
1 |
2 |
3,0±0,36 |
55,0±1,00 |
7,0±0,01 |
14 |
0 |
14 |
0 |
|
Динамичный ранний |
24 |
57 |
3,3±0,17 |
51,0±1,15 |
5,2±0,01 |
297 |
32 |
329 |
9,72 |
|
Динамичный средний |
19 |
41 |
3,2±0,16 |
52,3±0,44 |
6,2±0,01 |
255 |
22 |
277 |
7,94 |
|
Всего |
56 |
131 |
3,6±0,09 |
51,2±0,62 |
5,6±0,01 |
732 |
67 |
799 |
8,38 |
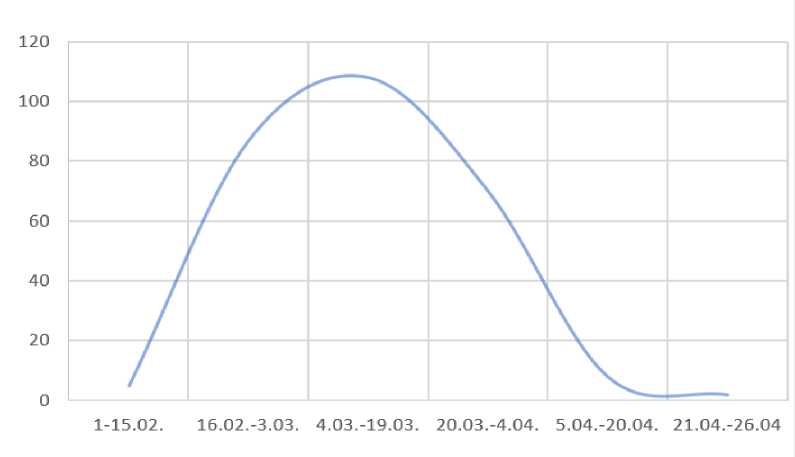
Рисунок 1 – Активность самцов в период гона
Так, мы видим, что покрытия с рождением наибольшего количества шенков (108 гол.) в помете приходятся на самок с ранними сроками прихода в гон (4.03. - 19.03.) и идёт на сильный спад в срок после 4.04. Это дает нам возможность судить о потентности самцов (через призму сроков гона самок), а именно мы видим, что самцы лучше кроют самок с ранним эструсом. Исходя из этого, в стаде необходимы самки, которые имеют статичный и стабильный характер эструса, то есть приходят в охоту с разницей в ±10
или ±15 дней от первого спаривания второго сезона размножения.
Кроме того, нами было рассчитано корреляционное отношение между датой покрытия самки и количеством рождённых щенков, которое составило 0,11, что свидетельствует о наличие слабой зависимости, но она есть.
Все выше приведённые данные взяты в совокупности за два исследуемых года, так как по каждому году имеется отчётность в виде результатов гона и щенения.
Таблица 4 – Результаты гона и щенения песца по ЗАО «Бирюли» в первый год исследования
|
Показатель |
Ед. изм. |
Молодые |
Взрослые |
Итого |
|
|
Пущено в гон |
гол. |
58 |
148 |
206 |
|
|
Прохолостело |
гол. |
5 |
1 |
6 |
|
|
% |
8,6 |
0,7 |
2,9 |
||
|
пропустовало |
гол. |
5 |
3 |
8 |
|
|
% |
8,6 |
2,0 |
3,9 |
||
|
НБР, аборт |
гол. |
5 |
5 |
10 |
|
|
% |
8,6 |
3,4 |
4,8 |
||
|
Пало самок до щенения |
гол. |
- |
- |
- |
|
|
% |
- |
- |
- |
||
|
Всего самок, не давших приплода |
гол. |
15 |
9 |
24 |
|
|
% |
25,8 |
6,08 |
11,6 |
||
|
Благополучно ощенилось |
гол. |
43 |
139 |
182 |
|
|
% |
74,1 |
93,9 |
88,3 |
||
|
У благополучно ощенившихся самок |
родилось щенков всего в т.ч. живых мертвых |
гол. гол. гол. |
546 516 30 |
1656 1605 51 |
2202 2121 81 |
|
плодовитость |
гол. |
12,7 |
11,9 |
12,09 |
|
|
количество мертвых |
гол. |
30 |
51 |
81 |
|
|
% |
5,81 |
3,17 |
3,81 |
||
|
пало щенков до регистрации |
гол. |
36 |
276 |
312 |
|
|
% |
7,0 |
17,2 |
14,7 |
||
|
Зарегистрировано |
всего щенков |
гол. |
480 |
1329 |
1809 |
|
на благополучно щенившуюся самку |
гол. |
11,1 |
9,56 |
9,93 |
|
|
на основную самку |
гол. |
8,27 |
8,97 |
8,78 |
|
Таблица 5 – Результаты гона и щенения песца по ЗАО «Бирюли» за второй год исследования
|
Показатель |
Ед.изм. |
Молодые |
Взрослые |
Итого |
|
|
Пущено в гон |
гол. |
79 |
175 |
254 |
|
|
Прохолостело |
гол. |
9 |
5 |
14 |
|
|
% |
11,4 |
0,3 |
5,5 |
||
|
пропустовало |
гол. |
7 |
7 |
14 |
|
|
% |
8,9 |
4 |
5,5 |
||
|
НБР, аборт |
гол. |
11 |
9 |
20 |
|
|
% |
13,9 |
5,1 |
7,8 |
||
|
Пало самок до щенения |
гол. |
- |
- |
- |
|
|
% |
- |
- |
- |
||
|
Всего самок, не давших приплод |
гол. |
27 |
21 |
48 |
|
|
% |
34,2 |
12 |
18,9 |
||
|
Благополучно ощенилось |
гол. |
52 |
154 |
206 |
|
|
% |
65,8 |
88 |
81,1 |
||
|
У благополучно ощенившихся самок |
родилось щенков всего в т.ч. живых мертвых |
гол. гол. гол. |
616 543 73 |
2076 1780 296 |
2592 2323 269 |
|
плодовитость |
гол. |
12,3 |
12,8 |
17,7 |
|
|
количество мертвых |
гол. |
73 |
296 |
269 |
|
|
% |
13,4 |
16,6 |
11,6 |
||
|
пало щенков до регистрации |
гол. |
112 |
402 |
517 |
|
|
% |
18,2 |
19,4 |
19,8 |
||
|
Зарегистрировано |
всего щенков |
гол. |
431 |
878 |
1809 |
|
на благополучно щенившуюся самку |
гол. |
8,29 |
8,95 |
8,78 |
|
|
на основную самку |
гол. |
5,4 |
7,8 |
7,1 |
|
По результатам первого года исследований (Таблица 4) наибольший процент холостых самок был выявлен у молодых (8 %), а по всему поголовью лишь 2,9 %. Такая же ситуация и с пропустовавшими самками – большинство из них молодые, а общий процент пропустовавших самок по стаду составил 3,9 %. Кроме того, по стаду высокий уровень абортов (4,8 %). При этом основным показателем гона, можно считать количество зарегистрированных щенков на основную самку. Так у взрослых самок этот показатель составил 8,97, а у молодых – 8,27 голов.
Во второй год исследования результаты гона (Таблица 5) намного ниже, так НБР и аборты, у молодых самок 13,7 %. Кроме того, в этом году выявлен недостаток таурина, вследствие чего у зверей отмечались признаки легочной недостаточности. Недостаток аминокислоты таурина был связан с отмеченным уже нами ранее, малым количеством углеводов или же рацион является неполноценным по белку.
Заключение. Таким образом, основное стадо состоит из самок с ранними сроками гона, но изучая характеры эструса, данные указали на большие разбросы в сроках гона (±30 дней), так как большинство самок имеет динамический характер эструса. Динамичный характер эструса мы считаем нежелательным, по причине трудности прогнозирования прихода самок в гон. Динамичный эструс нежелателен и в отношении самцов, так как лучшие показатели воспроизводства получают от самок, покрытых самцами в ранние сроки, а в случае разброса (±30) дней прихода самок в охоту, синхронизировать сроки проведения плодотворной случки является крайне тяжёлым занятием.
Резюме
Звероводство, как и все отрасли сельского хозяйства, на настоящий момент требует оптимизации непосредственно в отношении подхода к физиологическим циклам зверей. Отрасль относительно молода, доместикация зверей ещё продолжается, и моноцикличность зверей имеет свои определённые нюансы, что приводит к необходимости подстраиваться под условия рынка. В статье представлены результаты проведенных в республике Татарстан (ЗАО «Бирюли») исследований по изучению влияния даты начала и характера эструса на показатели воспроизводства самок песцов. Изучена работа самцов по результатам воспроизводства покрытых ими самок. Получены данные, которые свидетельствуют о том, что лучшие показатели воспроизводства у самок, покрытых самцами в ранние сроки эструса. Знания в отношении физиологии зверей и возможность контролирования сроков эструса самок и половой активности самцов, могут помочь посмотреть на возможности производства, более комплексно.
Список литературы Синхронность в сроках половой активности самок и самцов вуалевого песца в условиях звероводческого хозяйства
- Баранов, В. А. Улучшение производственного цикла при отборе самок на племя с желательным сроком и характером эструса / В. А. Баранов, Г. Х. Халилова, Р. Х. Равилов // Учёные записки КГАВМ. - 2019. - Т. 238. - С. 3234.
- Бекетов, С. В. Факторы, влияющие на плодовитость у пушных зверей промышленного разведения / С. В. Бекетов, А. Р. Корешков, К. С. Карпов // Кролиководство и звероводство. - 2016. -№ 5. - C. 13-17.
- Дормидонова, О. Ю. Продуктивные качества гибридов, полученных при гибридизации лисиц и песцов / О. Ю. Дормидонова // Санкт-Петербург, 2009. - С. 22.
- Сырников, Н. И. Воспроизводительная способность песцов в зависимости от упитанности, веса и длины тела / Н. И. Сырников, Е. М. Вальтман, Н. А. Петрова // Вопросы повышения воспроизводительной способности пушных зверей и кроликов: науч. тр. НИИПЗК. - Родники. - 1977. - Т. XV. - С. 193-197.
- Хенинг, О. Х. Мировое производство и торговля мехом в 2015-2016 гг. / О. Х. Хенинг // XVII Международная конференция по клеточному пушному звероводству. - 2016. - С. 56-59.
- Peura, J. Genetics of litter size, age at fi rst insemination and animal size in blue fox (Alopex lagopus) / J. Peura, I. Stranden, K. Smeds // Sdentifur. - 2004. - V. 28. - № 3. - P. 206-210.
- Balakirev, N. A. Development and application of iron supplements, and principles of iron deficiency anemia therapy in fur farming / N. A. Balakirev, V. I. Maksimov, A. A. Deltsov // Conf. Series: Earth and Environmental Science 848. - 2021. - P. 564569.
- Balakirev, N. A. Ways to develop and improve a pharmacologically ac 00082 / N. A. Balakirev, V. I. Maksimov, A. A. Deltsov // Fies. - 2020. - P. 17-21.
- Balakirev, N. A. Peculiarities of breeding work with red foxes obtained as a result of dominant color mutations / N. A. Balakirev, N. N. Shumilina, E. E. Larina, A. A. Galatskaya // Conf. Series:Earth and Environmental Science. - 2021. - P. 45-52.
- Fawcett, D. W. The mammalian spermatozoon / D. W. Fawcett // Dev. Biol. 1975. - V. 44(2). - P. 394-436.
- Miki, K. Glyceraldehyde 3-phosphate dehydrogenase-S, a sperm-specific glycolytic enzyme, is required for sperm motility and male fertility / K. Miki, W. Ou. E. Goulding [et al.] // Proc. Natl. Acad. Sci. USA. - 2004. - V. 101:16501-6. - P. 1116-1138.
- Schatten, H. The role of centrosomes in mammalian fertilization and its significance for ICSI. / H. Schatten, Q.-Y. Sun // Molecular Human Reprod. - 2009. - V. 15(9) . - P . 834-843.
- Sutovsky, P. Interactions of sperm perinuclear theca with the oocyte: implications for oocyte activation, anti-polyspermy defense, and assisted reproduction / P. Sutovsky, G. Manandhar, A. Wu, R. Oko // Microsc. Res. Tech. - 2003. - V. 61(4). - P. 324-352.
- Yanagida, K. Successful pregnancy after ICSI with strontium oocyte activation in low rates of fertilization / K. Yanagida, K. Morozumi, H. Katayose [et. al.] // Reprod. Biomed Online - 2006. - V. 13(6). - P. 12241231.
