Совершенствование системы оценки генетических и средовых факторов при составлении родительских пар в молочном скотоводстве

Автор: Шендаков А.И., Шендакова Т.А., Ханина Т.И., Климова С.П.
Журнал: Биология в сельском хозяйстве @biology-in-agriculture
Рубрика: Селекция и генетика в животноводстве
Статья в выпуске: 1 т.1, 2013 года.
Бесплатный доступ
Одна из задач современного животноводства - совершенствование системы управления селекции, особенно это касается биологических факторов (наследуемости, генетической корреляции, эффекта гетерозиса и пр.). В статье приведены результаты анализа влияния генетических и средовых факторов на молочную продуктивность скота, быков-производителей на генетические корреляции селекционных признаков у дочерей, наследования предрасположенности к болезням, влияния сезона оплодотворения на рождение двоен у чёрно-пёстрых коров, а также результаты инбридинга симментальского скота. В работе изучена динамика концентрации аллелей групп крови у быков разного происхождения и молочная продуктивность коров с разными генотипами по гену каппа-казеина.
Генетические и средовые факторы, селекция, чёрно-пёстрый и симментальский скот, инбридинг, аддитивная и неаддитивная изменчивость, наследуемость
Короткий адрес: https://sciup.org/14770266
IDR: 14770266 | УДК: 636.082.2:57.017.003.12
Текст научной статьи Совершенствование системы оценки генетических и средовых факторов при составлении родительских пар в молочном скотоводстве
ми [15, 17, 20], в том числе изучена генетическая корреляция между признаками бурого швицкого [17], голштинского [20], симментальского и чёрнопёстрого скота [13]. В настоящее время ведётся модернизация и оптимизация селекционных программ [3, 10], изучено иммуногенетическое сходствo большинства молочных пород, наиболее распространенных в России [11]. Однако следует признать, что в молочном скотоводстве постсоветского пространства оценке генетических и средовых факторов при составлении родительских пар и повышению эффективности подбора в стадах уделяется мало внимания, особенно мало внимания уделяется изучению генетико-статистических параметров при разных вариантах подбора, структуры генетической изменчивости и величины генетических корреляций не только удоев и жирности молока, но и других селекционных признаков. Данные факты объясняют актуальность наших исследований.
Целью наших исследований была разработка системы оценки генетических и средовых факторов в стадах молочного крупного рогатого скота при составлении родительских пар. В соответствии с целью исследований были поставлены задачи изучения сле- дующих величин и процессов: 1) распределения превосходства дочерей над матерями по жирности молока; 2) структуры генетической изменчивости жирности молока; 3) результатов увеличения молочной продуктивности чёрно-пёстрых коров, полученных при разной степени гетерогенности подбора; 4) наследования по матерям предрасположенности к заболеваниям; 5) влияния быков-производителей на генетические корреляции селекционных признаков у дочерей; 6) зависимости соотношения полов в потомстве от живой массы при первом оплодотворении и возраста коров; 7) влияния сезона оплодотворения на рождение двоен у чёрно-пёстрых коров; 8) детерминации роста чёрно-пёстрых тёлок генетическими и средовыми факторами; 9) зависимости интенсивности роста симментальских тёлок от степени возрастания гомозиготности; 10) динамики иммуногенетического полиморфизма у быков-производителей; 11) концентрации аллелей и генотипов по гену каппа-казеина, а также его ассоциаций с признаками молочной продуктивности.
Материалы и методы исследований
Для решения поставленных задач нами была начата комплексная работа по модернизации селекции молочного скота Орловской области. Были разработаны селекционно-генетические планы для 17 ведущих хозяйств Орловской области. Эффективность подбора коров по молочной продуктивности была изучена в ОПХ «Стрелецкое», СПК имени Мичурина и ООО «Фатнево». Интенсивность роста чёрнопёстрых тёлок – в ОПХ «Красная Звезда», ОАО «Агрофирма Мценская» и СПК имени Мичурина. Влияние генетических и средовых факторов на воспроизводительные качества коров изучались в ОПХ «Стрелецкое», СПК имени Мичурина, ЗАО «Куракинское», ОПХ «Красная Звезда» и др.
Генетическая корреляция между удоем и жирностью молока определялась по формулам Хейзеля:
rG
r ■ r xд yм yдxм или rG =
(г +г к 2
x д yм y д xм , где r –
фенотипические коэффициенты корреляции между признаками x и y у дочерей и матерей.
Коэффициент инбридинга (возрастания гомозиготности) вычисляли по формуле Райта-Кисловского: F x = Е [(2) n 1 + n 2 - 1 ■ (1 + f a )] ■ 100, где n i и П 2 — ряд, в котором находится общий предок с материнской и отцовской стороны, f a – величина коэффициента общего инбредного предка.
Величина концентрации эритроцитарных антигенов была определена как по методике, изложенной Е. К. Меркурьевой с соавт. в учебнике «Генетика» (1991), так и с целью выявления некоторых законо- мерностей – по формуле: ni , где ni – число жи-pi NA вотных, несущих в генотипе данный антиген, NА – сумма обнаруженных антигенов у всех животных.
Теоретическая основа исследований строилась на формуле: y=µ+G+B+е , где y – абсолютная фенотипическая ценность, µ – среднее значение признака в популяции (стаде), G – генетические факторы, B – систематические средовые факторы, е – случайные факторы.
Методика вычисления частоты встречаемости аллелей была построена на основе формулы Харди-Вайнберга (pA+qa)2=(p2AA+2pqAa+q2aa).
В работе были использованы каталоги фирм «Semex» и «Spermex Top Sires» (2010). Генетикостатистический анализ был проведён в компьютерной программе «Microsoft Excel».
Результаты собственных исследований
Наши исследования подтвердили наличие сложных генетических процессов при подборе и закреплении быков-производителей к стадам. Сила увеличения удоев в группах при разных вариантах подбора часто была слабо прогнозируемой, при этом точность прогноза по смежным признакам молочной продуктивности могла быть ещё более затруднительной. В этой ситуации интересным стало изучение генетикостатистических параметров при моделировании под- г ■ г xдxм yдyм
. r ■ r x д x м y д y м
Градации по жирности молока, +/-%
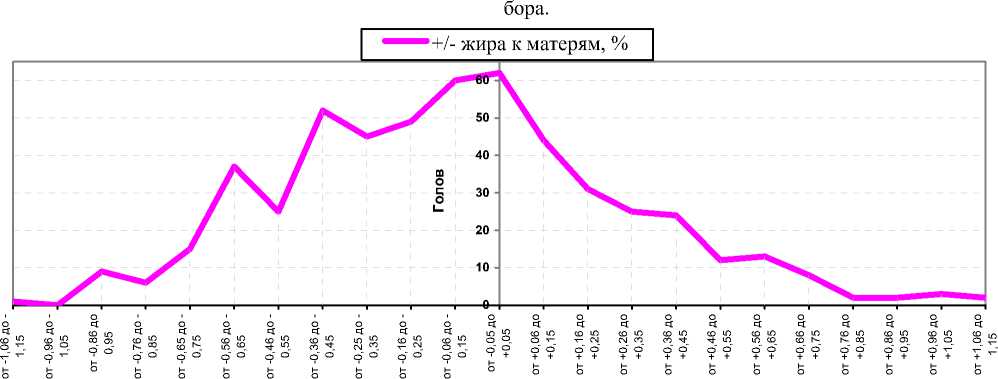
Рисунок 1. – Распределение эффективности подбора чёрно-пёстрых голштинизированных коров по жирности молока в ОПХ «Стрелецкое», +/-% к матерям (σ=0,29%, n=527)
Так, из рисунка 1 следует, что в стаде коров ОПХ «Стрелецкое» прослеживалось ассиметричное распределение превосходства дочерей над матерями по жирности молока (от -1,15 до +1,15%, или ±3,96σ, за пределами ±3σ находилось 0,95% особей). При этом наибольшее количество коров (n=62) практически не превысили и не понизили жирности молока в сравнении с матерями (около -0,05–+0,05%). Вместе с тем, достаточно большой массив дочерей (n=166) превзошёл своих матерей по жирности молока (от +0,06 до +1,15%, n=166).
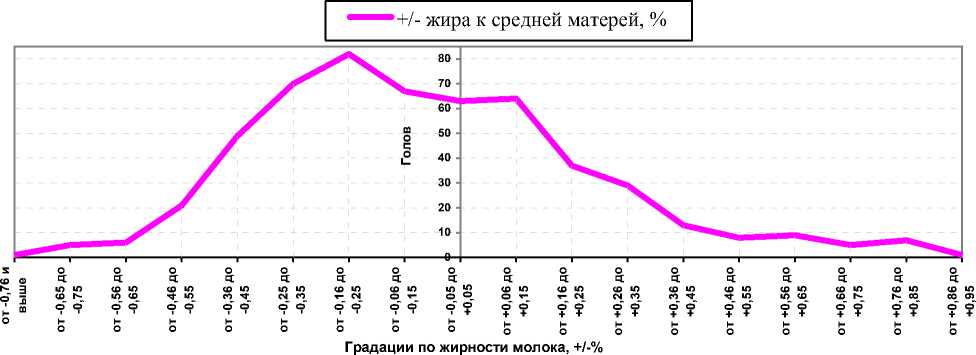
Рисунок 2. – Распределение превосходства чёрно-пёстрых голштинизированных дочерей над средней жирностью молока матерей по первой лактации, +/-% к средней жирности молока матерей (3,85%)
Наряду с этим распределение превосходства жирности молока дочерей над средней жирностью молока матерей стада также проявляло асимметрию (см. рисунок 2), т. е. большинство коров-дочерей имели меньшую жирность молока, чем средняя жирность у матерей, а наибольшая группа (от -0,16 до -0,25) была представлена 82 головами. Однако 81 корова имела превосходство над средней жирностью молока матерей в пределах от +0,16 до +0,55%.
Таблица 1. – Структура наследуемости селекционных признаков первой лактации у чёрно-пёстрых коров ОПХ «Стрелецкое» при сравнении дочерей и матерей по удою за 305 дней лактации
|
S к S и g § к ч с О |
н „ о ^ ^ ч 2 н о m К |
Генетические эффекты |
|||
|
В группе, превышающей удой матерей (n=384) |
В группе, не превышающей удой матерей (n=143) |
||||
|
h 2A |
d 2 |
h 2A |
d 2 |
||
|
Удой, кг |
0,150 |
-0,006 |
0,156 |
-0,243 |
0,393 |
|
Жир, % |
0,066 |
0,094 |
-0,028 |
-0,095 |
0,161 |
|
Жир, кг |
0,152 |
0,046 |
0,106 |
-0,187 |
0,339 |
Из таблицы 1 следует, что в группе, превышающей удой матерей (n=384), при общей наследуемости по удою h2=0,150 неаддитивные эффекты составляли 0,156 (15,6%). Однако влияние неаддитивной генетической изменчивости на суммирующие действие генов было, судя по всему, отрицательным по той причине, что у быков-производителей, давших случайные эффекты, были нежелательные гены. В данном случае основная доля неаддитивной наследуемости была полезной для селекции. В противовес данному явлению, в группе, не превышающей удой матерей (n=143), действие неаддитивной генетической изменчивости было негативным: по сути, её величина 0,393 показы- вает влияние препотентных быков-ухудшателей на признак.
В отличие от удоев, наследуемость жирности молока в лучшей группе коров (n=384) происходило в основном за счёт суммирующего действия генов. При этом неаддитивные эффекты, вызванные голштинскими быками, по жирности молока были отрицательными (-0,028). В худшей по удоям группе, напротив, за счёт неаддитивной наследуемости происходило увеличение жирности молока. По количеству молочного жира в лучшей по удоям группе коров прослеживался смешанный тип наследования. Следовательно, на увеличение или ухудшение удоев, уменьшение жирности и количества молочного жира в стаде существенное влияние оказывали случайные генетические эффекты (эпистаз, сверхдоминирование и пр.). Также возможно существенное влияние препо-тентных (доминантных по количественным признакам) отцов и сочетания генотипов матерей и отцов.
Таблица 2. – Структура наследуемости селекционных признаков первой лактации у чёрно-пёстрых коров ОПХ «Стрелецкое» при сравнении дочерей и матерей по жирности молока
|
ч с О |
н о 5 з i^ S 5 « м а 5 мп ю ч 2 II О g 3 О CQ К |
Генетические эффекты |
|||
|
В группе, превышающей жирность молока матерей (n=198) |
В группе, не превышающей жирность молока матерей (n=329) |
||||
|
h2 A |
d2 |
h2 A |
d2 |
||
|
Удой, кг |
0,150 |
0,039 |
0,111 |
0,182 |
-0,032 |
|
Жир, % |
0,066 |
0,126 |
-0,060 |
0,194 |
-0,128 |
|
Жир, кг |
0,152 |
0,040 |
0,102 |
0,146 |
0,006 |
Из таблицы 2 следует, что в группе, превышающей жирность молока матерей (n=198), при общей наследуемости по удою 0,150 неаддитивные эффекты составляли 0,111 (11,1%), прогнозируемые – 0,039 (3,9%). В группе, не превышающей жирность молока матерей (n=329), было очевидным негативное действие неаддитивной генетической изменчивости на удой, что объяснимо обычной закономерностью отрицательной корреляции между удоем и жирностью молока в популяциях крупного рогатого скота. Однако жирность молока в лучшей группе повышалась в основном за счёт суммирующего действия генов (h2A=0,126). При этом и в лучшей, и в худшей группе быки-производители, через которых проявлялись случайные факторы, имели негативное влияние на жирность молока (это связано ещё и с тем, что в последние годы жирность молока в стаде достигала 4,004,20%).
Таблица 3. – Генетическая корреляция между удоем и жирностью молока первой лактации у чёрно-пёстрых голштинизированных коров ОПХ «Стрелецкое» в разных группах
|
Генетическая группа |
n |
Фенотипическая корреляция, r |
r G |
|||
|
Удой матерей – удой дочерей |
Жир матерей – жир дочерей |
Удой матерей – жир дочерей |
Жир матерей – удой дочерей |
|||
|
Всё стадо |
542 |
0,075±0,043 |
0,033±0,043 |
0,055±0,043 |
0,059±0,043 |
0,003 |
|
Лучше средней жирности матерей (3,85%) |
198 |
0,019±0,071 |
0,063±0,071 |
0,103±0,070 |
0,005±0,071 |
0,001 |
|
Хуже средней жирности матерей (3,85%) |
323 |
0,091±0,055 |
0,097±0,055 |
-0,058±0,055 |
0,014±0,056 |
≈0 |
|
Лучше среднего удоя матерей (3782 кг) |
384 |
-0,003±0,051 |
0,047±0,051 |
0,032±0,051 |
0,011±0,051 |
– |
|
Хуже среднего удоя матерей (3782 кг) |
143 |
-0,121±0,083 |
-0,048±0,084 |
0,027±0,084 |
0,143±0,082 |
– |
|
Коровы по превосходству над средним удоем матерей от -500 до +500 кг (от 3782 кг) |
224 |
0,108±0,066 |
0,021±0,067 |
0,139±0,066 |
0,053±0,067 |
0,004 |
|
Коровы по превосходству над средним удоем матерей от -500 до +1000 кг (от 3782 кг) |
342 |
0,112±0,053 |
0,022±0,054 |
-0,055±0,054 |
0,033±0,054 |
≈0 |
Из таблицы 3 следует, что во всём исследуемом стаде ОПХ «Стрелецкое» в 2011 году (n=542) прослеживалась почти нулевая корреляция между удоем и жирностью молока (+0,003). Положительная генетическая корреляция между удоем и жирностью молока была только в двух группах: в группе, где коровы превышали средние значения матерей по жирности молока (n=198), и в группе, состоявшей из коров, средних по превосходству над матерями по удоям (от -500 до +500 кг молока). В первом случае это объяснимо более высокой аддитивной наследуемостью жирности молока, во втором – общей аддитивной наследуемостью двух коррелирующих признаков. При этом в группе с удоями, крайне отклоняющимися от среднего, прослеживались неаддитивные эффекты не только по удоям, но и по жирности молока. Отцы способствовали проявлению в стаде случайных эффектов при скрещивании.
Таблица 4. – Влияние гомогенного и гетерогенного подбора на продуктивные качества чёрно-пёстрых коров по первой лактации в СПК им. Мичурина
|
Удой матерей, кг |
Средняя разница в удоях матерей отцов и матерей, кг |
n |
Продуктивные признаки |
|||
|
Удой за 305 дней, кг |
Жир, % |
Жир, кг |
Живая масса, кг |
|||
|
более 5000 |
536 |
14 |
3361 ±155 |
3,73 ±0,05 |
125,3 ±3,5 |
444 ±6 |
|
40004999 |
3259 |
51 |
3743 ±78* |
3,73 ±0,02 |
140,3 ±2 4*** |
443 ±4 |
|
30003999 |
4934 |
173 |
3868 ±52** |
3,72 ±0,01 |
144,5 ±2,1*** |
446 ±3 |
|
20002999 |
5672 |
83 |
3787 ±48* |
3,70 ±0,02 |
140,1 ±2,3*** |
438 ±4 |
|
менее 2000 |
6042 |
7 |
4171 ±198** |
3,69 ±0,05 |
154,6 ±4,8*** |
468 ±7* |
Примечание: *– p<0,05, **– p<0,01.
Схожие тенденции нами были получены в СПК им. Мичурина Орловской области. При этом в данном хозяйстве (см. таблицу 4) прослеживалась высокая эффективность гетерогенного подбора высокоценных быков к коровам-матерям с удоями менее 2000 кг молока, в результате чего их дочери дали по первой лактации в среднем 4171 кг молока и достоверно превзошли группу, полученную методом гомогенного подбора, на 810 кг. В данном хозяйстве неверным подходом было закрепление к коровам-рекордисткам быков с невысоким потенциалом (50005500 кг молока). Это привело к уменьшению удоев в группе на 1500-2000 кг молока и более.
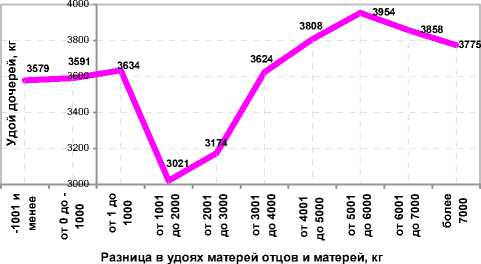
Рисунок 3. – Влияние гетерогенности подбора по молочной продуктивности на удой чёрно-пёстрых коров СПК им. Мичурина по первой лактации (n=335)
Вместе с тем, превосходство матерей отцов над матерями коров выше 6000 кг молока вело к снижению удоев за 305 дней первой лактации в стаде с 3954 до 3775 кг (см. рисунок 3). Худшие удои по первой лактации были получены при превосходстве матерей отцов над матерями на уровне от 1001 до 3000 кг (3021-3174 кг), однако гомогенный подбор также дал относительно неплохие результаты для данного хозяйства – до 3591-3634 кг молока.
Следовательно, грамотно построенный подбор может не только увеличить удой за одно поколение на 1000-2000 кг молока и более, но и существенно повлиять на генетико-статистические параметры, в частности на структуру генетической изменчивости и генетические корреляции между селекционными признаками. Гомогенный подбор является наиболее сложным с этой точки зрения, поскольку при нём сложно получить эффект гетерозиса.
Таблица 5. – Фенотипическая и генетическая корреляция между удоем и жирностью молока у дочерей чёрно-пёстрых и голштинских быков-производителей в ОПХ «Стрелецкое»
|
Кличка и № быка |
n |
Фенотипическая корреляция, r |
r G |
|||
|
удой матерей – удой дочерей |
жир матерей – жир дочерей |
удой матерей – жир дочерей |
жир матерей – удой дочерей |
|||
|
Риголетто 9862 |
77 |
0,222 ± 0,112 |
0,077 ± 0,115 |
0,093 ± 0,115 |
0,065 ± 0,115 |
0,595* |
|
Левкой 110 |
13 |
0,128 ± 0,299 |
-0,316 ± 0,286 |
0,339 ± 0,284 |
-0,276 ± 0,299 |
– |
|
Хезелден 474459 |
30 |
0,277 ± 0,181 |
0,107 ± 0,188 |
-0,161 ± 0,186 |
0,017 ± 0,189 |
-0,418* |
|
4267 |
18 |
-0,037 ± 0,250 |
0,173 ± 0,246 |
-0,325 ± 0,236 |
0,063 ± 0,249 |
– |
|
Эксперт 661277 |
15 |
0,244 ± 0,269 |
0,613 ± 0,219 |
-0,317 ± 0,263 |
0,243 ± 0,269 |
-0,310* |
|
Жамес 016 |
49 |
0,284 ± 0,114 |
0,227 ± 0,142 |
0,237 ± 0,142 |
-0,038 ± 0,146 |
0,392* |
|
Элеватор 482077 |
15 |
-0,006 ± 0,278 |
-0,147 ± 0,274 |
-0,069 ± 0,277 |
0,465 ± 0,245 |
– |
|
Тино 379457 |
16 |
0,088 ± 0,266 |
0,304 ± 0,255 |
-0,044 ± 0,267 |
-0,222 ± 0,260 |
0,604** |
|
Эльвио 9942 |
26 |
-0,337 ± 0,192 |
0,341 ± 0,192 |
-0,071 ± 0,204 |
-0,006 ± 0,204 |
– |
|
Лель 23682 |
17 |
-0,077 ± 0,257 |
-0,103 ± 0,257 |
-0,415 ± 0,235 |
0,056 ± 0,258 |
– |
|
2970 |
15 |
-0,108 ± 0,276 |
-0,370 ± 0,258 |
0,158 ± 0,274 |
-0,008 ± 0,277 |
– |
|
2944 |
12 |
-0,178 ± 0,311 |
-0,312 ± 0,300 |
-0,187 ± 0,311 |
-0,326 ± 0,299 |
– |
|
Резонанс 203 |
49 |
0,138 ± 0,144 |
-0,032 ± 0,146 |
-0,028 ± 0,146 |
-0,252 ± 0,141 |
– |
|
Пир 695 |
39 |
0,096 ± 0,164 |
-0,052 ± 0,164 |
0,068 ± 0,164 |
-0,065 ± 0,164 |
– |
|
Персуадер 13064124 |
51 |
0,068 ± 0,142 |
0,201 ± 0,140 |
0,096 ± 0,142 |
-0,045 ± 0,143 |
0,218* |
Примечание: *– p<0,05, **– p<0,01.
Из таблицы 5 следует, что в ОПХ «Стрелецкое», кроме того, у дочерей только четырёх быков прослеживалась положительная генетическая корреляция между удоем и жирностью молока (от 0,218 до 0,604). У дочерей Хезелдена и Эксперта была получена отрицательная связь (-0,418 и -0,310 соответственно). Для большинства быков было характерно отсутствие генетической корреляции между удоем и жирностью молока. При этом не всегда самый лучший бык, согласно традиционным способам оценки, способствовал одновременному улучшению и удоя, и жирности молока. В этой ситуации мы отдавали предпочтение быкам, чьи дочери отличались высокими продуктив- ными показателями и положительной генетической корреляцией.
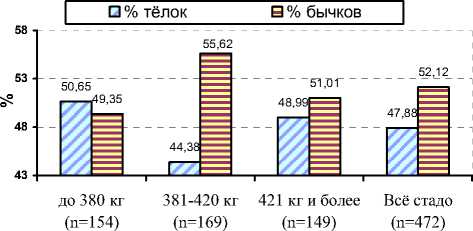
Согласно нашим исследованиям, на соотношение полов в потомстве чёрно-пёстрых коров некоторое влияние оказала живая масса тёлок при первом осеменении (см. рисунок 4).
Так, наибольшее количество бычков (55,62%) рождалось в группе матерей-первотёлок, имеющих при первом осеменении живую массу 381-420 кг. При этом с возрастанием живой массы тёлок при первом осеменении с 380 до 420 кг и более процент мёртворожденных телят уменьшался с 6,67 до 2,61%.
Живая масса тёлок при первом осеменении
Рисунок 4. – Влияние живой массы чёрно-пёстрых тёлок при первом осеменении на соотношение полов в потомстве в ОПХ «Стрелецкое»
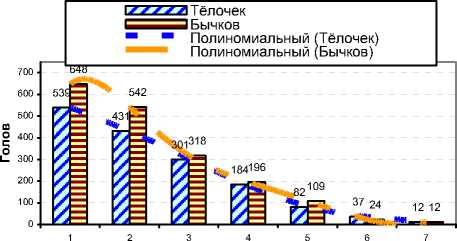
Лактации
Рисунок 5. – Количество родившихся тёлочек и бычков в хозяйствах в период проведения исследований (n=3435)
Существенного влияния возраста коров (по лактациям) на соотношение полов получено не было (см. рисунок 5), однако практически во все отёлы рождалось больше бычков – до 57,07%. При этом % рождения тёлочек с каждой последующей лактацией имел тенденцию к выравниванию.
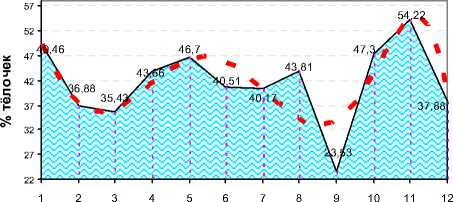
Месяцы оплодотв орения
Рисунок 6. – Проценты родившихся тёлочек в хозяйствах Орловской области в период проведения исследований (n=3435)
Из рисунка 6 следует, что по месяцам оплодотворения, начиная с января, наблюдалась существенная флуктуация соотношения полов. Наименьший процент тёлочек был в стадах при оплодотворении в сентябре – всего 23,53%, наибольший – при оплодотворении в ноябре (54,22%). Однако соотношение полов скорее имело случайную форму проявления.
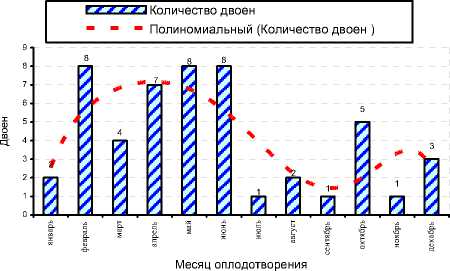
Рисунок 7. – Количество двоен у чёрно-пёстрых гол-штинизированных коров по месяцам оплодотворения в хозяйствах Орловской области (всего 1936 живых телят)
Была намечена тенденция влияния месяца оплодотворения на рождение двоен (см. рисунок 7). Так, наибольшее количество двоен в исследованных хозяйствах было получено при оплодотворении в период с февраля по июнь, а количество телят-двоен ко всему полученному поголовью по весенним месяцам оплодотворения составляло до 6,5-7,0%.
Таблица 6. – Влияние продолжительности сервис-периода на формирование пола телёнка в ОПХ «Стрелецкое»
|
Отёл |
Сервис-период, дней |
Живых телят, голов |
% тёлок |
Мёртвых телят, голов |
|
второй |
до 80 |
74 |
52,70 |
1 |
|
81-120 |
102 |
43,14 |
4 |
|
|
121-160 |
74 |
39,19 |
3 |
|
|
161 и более |
178 |
44,94 |
7 |
|
|
третий |
до 80 |
80 |
50,00 |
2 |
|
81-120 |
79 |
54,43 |
0 |
|
|
121-160 |
38 |
60,53 |
0 |
|
|
161 и более |
128 |
56,25 |
4 |
Из таблицы 6 следует, что во второй отёл продолжительность предшествующего сервис-периода незначительно влияла на соотношение полов. Так, при увеличении сервис-периода до 160 дней родилось 39,19% тёлочек. По третьему отёлу рождались преимущественно тёлочки – от 50 до 60,53%. Очевидно, к третьему отёлу коровы ослабевают, и, согласно теории Trivers и Willard (1973), им проще выносить тёлочку. Рождение большего количества бычков по второму отёлу при сервис-периоде до 160 дней, очевидно, обусловлено тем, что корова перед плодотворным осеменением длительное время накапливает резервы питательных веществ. Увеличение сервис-периода до 200 дней сопровождается болезнями и ослаблением здоровья, что снова косвенно способствует измене- нию физиологических процессов в организме и способствует повышению вероятности рождения тёлочки.
В целом, можно сделать вывод, что в скотоводческих хозяйствах, разводящих чёрно-пёструю и симментальскую породу, существуют предпосылки улучшения воспроизводительных качеств коров за счёт селекции и планирования подбора по месяцам. В проведённых исследованиях инбридинг не оказывал существенного влияния на мёртворождаемость. Процент генов голштинской породы также оказывал влияние на распределение полов в потомстве. Для уменьшения количества мёртворожденных телят было целесообразно рекомендовать интенсивное выращивание тёлок (720-750 г в сутки в среднем) и осеменение их в 17-18 месяцев при живой массе 421 кг и более. С возрастом, при истощении резервов организма коровы чаще рождали тёлочек, чем бычков, поэтому важно создавать приходящим в охоту коровам благополучные условия содержания для нормального соотношения полов. Следовательно, в селекции молочного скота существуют некоторые предпосылки планирования пола даже без применения сексирован-ного семени. Средовые факторы играют важную роль в селекционно-генетической работе.
Мастит
Гинекологические заболевания Болезни конечностей Лейкоз
Трудные отёлы Яловость Прочее
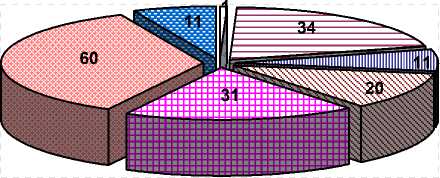
Рисунок 8. - Причины выбытия коров из стада при выбраковке их матерей из-за гинекологических заболеваний (n=168)
Мастит
Гинекологические заболевания
Болезни конечностей
Лейкоз
Трудные отёлы
Яловость
Прочее
Рисунок 9. - Причины выбытия коров из стада при выбраковке их матерей из-за яловости (n=97)
Анализ наследования причин выбытия коров в ОПХ «Стрелецкое» показал, что при выбраковке ко- ров по причине гинекологических заболеваний у их дочерей наблюдались различные заболевания, по причине которых они также рано выбывали из стада (см. рисунок 8). Особенно у дочерей проявлялись трудные отёлы (n=31), яловость (n=60) и гинекологические заболевания (n=34).
При выбраковке коров по причине яловости (n=97) почти половина их дочерей (n=44, или 45,4%) также выбывала по причине яловости (см. рисунок 9).
При этом большое количество голов выбывало из-за гинекологических болезней и трудных отёлов (n=14 и 10 соответственно). Необходимо отметить, что при трудных отёлах у матерей до 74% дочерей проявляли яловость, а мастит и болезни конечностей наследовались, судя по полученным данным, не столь сильно, как гинекологические заболевания и яловость. Следовательно, в перспективе это можно учитывать при составлении родительских пар.
Таблица 7. – Влияние генетических и средовых факторов на интенсивность роста чёрно-пёстрых тёлок в зависимости от процента генов голштинской породы и линейной принадлежности
|
Группа |
Хозяйство |
В среднем по хозяйствам |
||||||||||
|
ОПХ «Красная Звезда» |
ОАО «Агрофирма Мценская» |
СПК имени Мичурина |
||||||||||
|
n |
r w |
1- r w |
n |
r w |
1- r w |
n |
r w |
1- r w |
n |
r w |
1- r w |
|
|
Процент генов голштинской породы (% HF) |
||||||||||||
|
0 |
- |
- |
- |
229 |
0,324 |
0,676 |
72 |
0,057 |
0,943 |
301 |
0,191 |
0,809 |
|
12,5 |
10 |
0,173 |
0,827 |
20 |
0,433 |
0,567 |
72 |
0,652 |
0,348 |
102 |
0,419 |
0,581 |
|
25,0 |
34 |
0,525 |
0,475 |
76 |
0,330 |
0,670 |
17 |
0,306 |
0,694 |
127 |
0,387 |
0,613 |
|
37,5 |
60 |
0,140 |
0,860 |
110 |
0,093 |
0,907 |
55 |
0,242 |
0,758 |
225 |
0,158 |
0,842 |
|
50,0 |
50 |
0,461 |
0,539 |
61 |
0,116 |
0,884 |
62 |
0,230 |
0,770 |
173 |
0,268 |
0,731 |
|
62,5 |
35 |
0,433 |
0,567 |
- |
- |
- |
11 |
0,521 |
0,490 |
46 |
0,477 |
0,523 |
|
75,0 |
15 |
0,151 |
0,849 |
- |
- |
- |
- |
- |
- |
15 |
0,151 |
0,849 |
|
в т. ч.инбридинг |
18 |
0,378 |
0,622 |
- |
- |
- |
- |
- |
- |
18 |
0,378 |
0,622 |
|
Отцовские линии |
||||||||||||
|
А. Адема |
- |
- |
- |
240 |
0,320 |
0,680 |
- |
- |
- |
240 |
0,320 |
0,680 |
|
Х. Адема |
- |
- |
- |
128 |
0,292 |
0,708 |
- |
- |
- |
128 |
0,292 |
0,708 |
|
Орла |
- |
- |
- |
17 |
0,456 |
0,544 |
51 |
0,119 |
0,881 |
68 |
0,287 |
0,713 |
|
Р. Соверинг |
15 |
0,393 |
0,607 |
126 |
0,026 |
0,974 |
84 |
0,626 |
0,374 |
225 |
0,348 |
0,652 |
|
М. Чифтейн |
64 |
0,140 |
0,860 |
- |
- |
- |
- |
- |
- |
64 |
0,140 |
0,860 |
|
В. Б. Айдиал |
109 |
0,218 |
0,782 |
- |
- |
- |
128 |
0,243 |
0,757 |
237 |
0,231 |
0,769 |
|
Все линии |
222 |
0,301 |
0,699 |
496 |
0,273 |
0,727 |
289 |
0,333 |
0,667 |
1007 |
0,282 |
0,718 |
Из таблицы 7 следует, что в хозяйствах Орловской области сила влияния генетических и средовых факторов на интенсивность роста чёрно-пёстрых тёлок в зависимости от их кровности по голштинам и линейной принадлежности по отцу может быть разной. Так, в ОПХ «Красная Звезда» по влиянию генетических факторов отличался генотип с кровностью 25,0% по голштинам и тёлки линии Р. Соверинг (r w =0,525 и 0,393 соответственно), в ОАО «Агрофирма Мценская» – животные с 12,5% генов голштинской породы и тёлки линии Орла (rw=0,433 и 0,456 соответственно), в СПК им. Мичурина также отличались низкокровные тёлки и тёлки линии Р. Соверинг (rw=0,652 и 0,626 соответственно). В этих генетических группах влияние средовых факторов (1- rw) было наименьшим, а по всем хозяйствам наибольшее влияние генотипа на интенсивность роста было у тёлок с кровностью 62,5% по голштинам, среди линий сложно было выделить наиболее удачные по устойчивости роста. Влияние генов на реализацию живой массы составило в среднем 20-50%, а паратипических условий – 50-80%. Учитывая наличие вариаций результативности скрещивания, можно сделать вывод о возможности ведения подбора на увеличение устойчивости роста молодняка.
В инбредном поголовье симментальского и сим-ментал-голштинского скота ООО «Фатнево» (см. таблицу 8) наибольшей живой массой при рождении об- ладали 2 тёлки, полученные в результате кровосмешения (32,5 кг), однако тёлки (n=30), полученные при топбридинге и топкроссбридинге (т.е. с применением инбредных отцов в степени IV-IV и V-V), также дали более высокую живую массу – 31,2 кг. Начиная с первого месяца выращивания, наибольшую живую массу имели тёлки, полученные при кровосмешении и умеренном родстве: живая масса от 1 до 12 месяца возросла у них от 57,0 и 53,5 до 268,0 и 266,0 кг соответственно. Причём в 7, 8, 9 и 10 месяцев тёлки с Fx=25,0% превосходили тёлок с Fx=0,781% на 24,8, 31,0, 41,7 и 43,2 кг соответственно (p<0,1-0,05). Впоследствии животные этих групп смогли набрать к 18 месяцам живую массу, необходимую для нормального оплодотворения, а тёлки, полученные в результате боттомкроссинга (т.е. при оплодотворении инбредных матерей семенем аутбредных быков), к 12 месяцам дали худший показатель – всего 211,5±5,7 кг, что уступило тёлкам с Fx=25,0% 56,5 кг. Следует отметить, что вариабельность живой массы у всего инбредного поголовья (n=87) во все периоды выращивания была достаточно высокой для данного признака – от 6,64 до 18,42%, однако в отдельные периоды в разных группах были замечены тенденции ещё большего увеличения или, напротив, уменьшения фенотипической изменчивости. Так, низкий Сv был у 8 тёлок, полученных методом ин-энд-инбридинга, т. е. инбридинга в нескольких поколениях, а следовательно, с высокой степенью возрастания гомозиготности (с 4 по 12 месяц Сv был в пределах 5,6-8,1%). Схожую тенденцию дали инкроссбридинг и инбредлайнкроссинг в период от рождения до 5 месяцев (Сv=3,3-9,9%). Это означает, что тёлки в этих группах в данные периоды выращивания набирали живую массу ровно, в то время как высокая изменчивость признака была у тёлок, полученных методом боттомкоросинга, инбредлайн-кроссинга и инкроссбридинга, в периоды 7-8 и 8-10 месяцев соответственно (до 25,1%). В целом следует отметить, что инбридинг дал парадоксальные результаты: селекционные мероприятия, направленные на понижение инбредной депрессии (боттомкроссинг), не принёс ожидаемых положительных результатов, а целенаправленное кровосмешение привело к закреплению интенсивного роста тёлок, хотя при инбридинге это встречается крайне редко. Выводы можно завершить тем, что инбредная группа тёлок (n=87) уступала средним показателям стада (n=350) в разные периоды выращивания от 5 до 15 кг живой массы.
Таблица 8. – Влияние вариантов родственного спаривания на интенсивность роста симментальских тёлок от рождения до 12 месяцев в ООО «Фатнево»
|
Разновидность инбридинга |
n |
Параметры |
Живая масса по месяцам (от рождения до 12), кг |
||||||||||||
|
при рожд. |
1 |
2 |
3 |
4 |
5 |
6 |
7 |
8 |
9 |
10 |
11 |
12 |
|||
|
Кровосмешение (клозебридинг) F x =25,0% |
2 |
M |
32,5 |
57,0 |
84,5 |
108,0 |
125,0 |
133,0 |
149,0 |
172,0 |
196,5 |
224,5 |
241,0 |
253,5 |
268,0 |
|
±m |
0,5 |
5,0 |
8,5 |
12,1 |
14,0 |
11,1 |
14,0 |
18,1 |
16,5 |
8,51 |
9,0 |
11,6 |
12,1 |
||
|
σ |
0,7 |
7,1 |
12,0 |
17,0 |
19,8 |
15,6 |
19,8 |
25,5 |
23,3 |
12,0 |
12,7 |
16,3 |
17,0 |
||
|
C v, % |
2,1 |
12,4 |
14,2 |
15,7 |
15,8 |
11,7 |
13,3 |
14,8 |
11,9 |
5,3 |
5,2 |
6,4 |
6,3 |
||
|
Ин-энд-инбридинг (т.е. в нескольких поколениях) |
8 |
M |
28,2 |
45,1 |
64,2 |
86,0 |
98,1 |
110,4 |
124,4 |
138,1 |
150,6 |
167,7 |
190,0 |
205,6 |
221,6 |
|
±m |
0,2 |
2,1 |
3,3 |
4,1 |
2,5 |
2,1 |
3,6 |
2,9 |
2,8 |
4,1 |
4,7 |
4,3 |
4,4 |
||
|
σ |
0,7 |
6,0 |
9,4 |
11,5 |
7,0 |
5,9 |
10,1 |
8,1 |
7,8 |
11,5 |
13,3 |
12,2 |
12,4 |
||
|
C v, % |
2,5 |
13,3 |
14,6 |
13,4 |
7,1 |
5,3 |
8,1 |
5,9 |
5,2 |
6,9 |
7,0 |
5,9 |
5,6 |
||
|
Умеренное родство (IV-IV) F x =0,781% |
6 |
M |
28,8 |
53,5 |
77,8 |
96,2 |
112,3 |
124,5 |
139,2 |
147,2 |
165,5 |
182,8 |
197,8 |
215,8 |
266,0 |
|
±m |
0,3 |
2,7 |
4,9 |
2,5 |
5,0 |
7,3 |
10,6 |
10,4 |
10,0 |
9,8 |
10,4 |
10,1 |
10,2 |
||
|
σ |
0,8 |
6,7 |
12,1 |
6,1 |
12,3 |
17,9 |
25,9 |
25,4 |
24,43 |
24,00 |
25,52 |
24,84 |
25,01 |
||
|
C v, % |
2,8 |
12,5 |
15,6 |
6,3 |
10,9 |
14,4 |
18,6 |
17,2 |
14,8 |
13,1 |
12,9 |
11,5 |
9,4 |
||
|
Боттомкроссинг (т.е. инбредные матери) |
35 |
M |
29,9 |
50,9 |
70,0 |
86,6 |
100,9 |
111,4 |
121,5 |
132,4 |
147,0 |
162,2 |
178,3 |
194,7 |
211,5 |
|
±m |
0,3 |
1,0 |
1,8 |
2,2 |
2,8 |
3,3 |
4,0 |
4,8 |
5,3 |
5,4 |
5,5 |
5,6 |
5,7 |
||
|
σ |
1,5 |
6,1 |
10,7 |
12,8 |
16,5 |
19,5 |
23,8 |
28,4 |
31,2 |
31,7 |
32,3 |
33,0 |
33,7 |
||
|
C v, % |
5,2 |
12,0 |
15,3 |
14,8 |
16,4 |
17,5 |
19,6 |
21,5 |
21,2 |
19,6 |
18,1 |
16,9 |
15,9 |
||
|
Топбридинг и топкроссбридинг (т.е. инбредные отцы) |
30 |
M |
31,2 |
51,2 |
71,8 |
89,7 |
101,9 |
113,9 |
125,7 |
139,2 |
155,1 |
174,0 |
190,6 |
205,4 |
221,0 |
|
±m |
0,4 |
1,0 |
1,6 |
1,6 |
1,9 |
2,3 |
3,1 |
4,0 |
4,4 |
5,0 |
6,0 |
6,4 |
6,5 |
||
|
σ |
2,4 |
5,3 |
8,6 |
9,0 |
10,3 |
12,8 |
17,0 |
22,1 |
23,9 |
27,7 |
32,8 |
35,2 |
35,9 |
||
|
C v, % |
7,7 |
10,4 |
12,0 |
10,0 |
10,1 |
11,2 |
13,5 |
15,9 |
15,4 |
15,9 |
17,2 |
17,1 |
16,2 |
||
|
Инбредлайнкрос-синг и инкроссб-ридинг |
6 |
M |
29,2 |
47,2 |
68,0 |
88,8 |
102,7 |
107,3 |
111,3 |
117,7 |
130,8 |
152,5 |
173,8 |
190,8 |
217,0 |
|
±m |
0,4 |
1,9 |
2,7 |
2,3 |
2,6 |
4,3 |
6,7 |
9,4 |
13,0 |
15,6 |
14,4 |
13,6 |
13,7 |
||
|
σ |
1,0 |
4,7 |
6,5 |
5,7 |
6,0 |
10,5 |
16,4 |
23,0 |
32,0 |
38,3 |
35,4 |
33,3 |
33,5 |
||
|
C v, % |
3,3 |
9,9 |
9,6 |
6,4 |
5,9 |
9,8 |
14,8 |
19,5 |
24,4 |
25,1 |
20,4 |
17,4 |
15,5 |
||
|
Всё инбредное поголовье |
87 |
M |
30,1 |
50,4 |
70,7 |
88,9 |
102,4 |
113,2 |
124,1 |
135,9 |
151,2 |
168,8 |
185,8 |
201,4 |
218,7 |
|
±m |
0,2 |
0,7 |
1,1 |
1,2 |
1,4 |
1,7 |
2,2 |
2,7 |
3,0 |
3,2 |
3,4 |
3,5 |
3,6 |
||
|
σ |
2,0 |
6,1 |
10,1 |
11,1 |
13,5 |
15,9 |
20,7 |
25,0 |
27,8 |
30,0 |
31,7 |
32,2 |
33,5 |
||
|
C v, % |
6,6 |
12,2 |
14,3 |
12,5 |
13,2 |
14,1 |
16,7 |
18,4 |
18,4 |
17,8 |
17,1 |
16,0 |
15,3 |
||
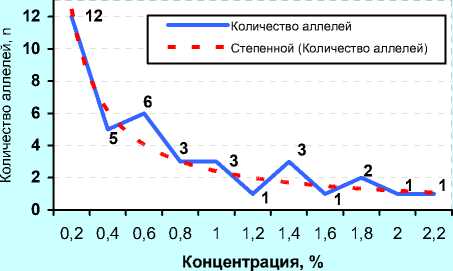
Из рисунка 10 (А) следует, что у чёрно-пёстрых голштинов «ЦСИО» в 2001 году 12 антигенов имели популяционный вес менее 0,2%, было обнаружено 5 аллелей с концентрацией от 0,2 до 0,4% и 6 аллелей с концентрацией от 0,41 до 0,6%. С увеличением частоты аллеля происходило уменьшение количества аллелей, имеющих такую же частоту, т. е. высокочастотных аллелей в стадах всегда меньше, чем низкочастотных. Однако данная тенденция не проявляла линейной зависимости, а спадала по гиперболе первого порядка, которую мы выразили математической формулой (А): у = 0,23511 + 3,13837 , где х — это частота xx аллеля. Расчёт по данной формуле для прогноза количества эритроцитарных антигенов, имеющих опреде- лённую частоту, коррелировала с фактическими показателями (r=0,963).
В 2008 году у быков, семя которых реализовывала ОАО «ЦСИО», увеличилось количество аллелей с популяционным весом 0,2, 1,2, 1,4, 1,6, 1,8 и 2,2%. С одной стороны, это объяснимо усилением иммуноге-нетического разнообразия, с другой – накоплением желательных аллелей, являющихся, очевидно, в некоторой степени маркерами каких-либо продуктивных качеств. В данном случае можно было прогнозировать количество аллелей по формуле (Б):
- 7.3S21S7 „ , у = 0 09367+ 2,38218' с высокой точностью (r=0,967). x
x
Концентрация, %
А) ФГУП «ЦСИО» (2001 г., n=39)
Г) ОАО «ЦСИО» (2008 г., n=82)
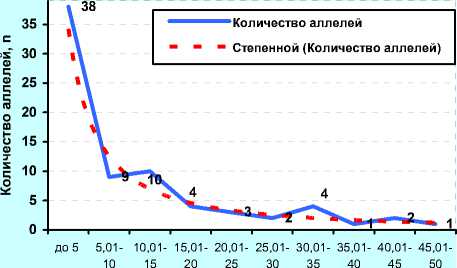
Концентрация, %
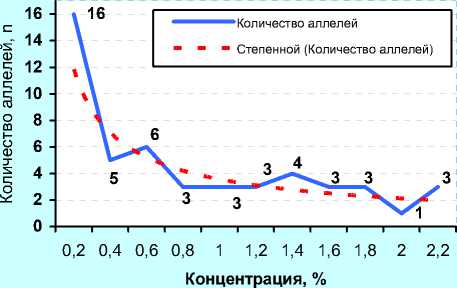
Б) ОАО «ЦСИО» (2008 г., n=82)
Д) ОАО «Орловское» (2011 г., n=41)
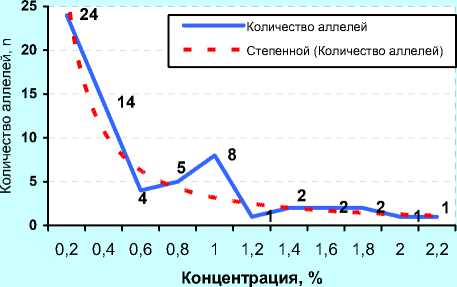
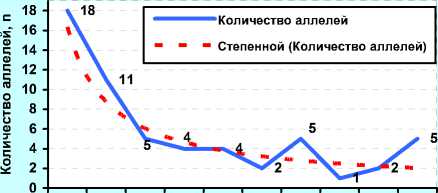
до 5 5,01- 10,0 15,0 20,0 25,0 30,0 35,0 40,0 45,0
10 1-15 1-20 1-25 1-30 1-35 1-40 1-45 1-50
Концентрация, %
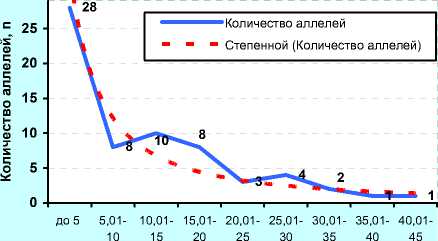
В) ОАО «Орловское» (2011 г., n=41)
Е) ОПХ «Стрелецкое» (коровы, 2010 г., n=33)
Рисунок 10. – Иммуногенетическое разнообразие по группам аллелей с разной концентрацией у чёрно-пёстрых голштинских быков-производителей в зависимости от происхождения и у коров племенного ядра ОПХ «Стрелецкое» Орловской области
В отличие от ОАО «ЦСИО», быки ОАО «Орловское» были более разнообразны по иммуногенетиче-ским факторам: 24 аллеля имели популяционный вес около 0,2, т. е. в 2 раза выше, чем у быков ОАО «ЦСИО».
При этом 14 и 8 антигенов имели вес 0,4 и 1,0 соответственно. В данном случае прогноз по уравнению гиперболы первого порядка (В)
У = 0,6755 +2,81925 сопровождался высокой точно-xx стью (r=0,949).
Вместе с тем, и у быков ОАО «ЦСИО», и у быков в ОАО «Орловское» были стабильно сохраняющиеся антигены, выходящие за пределы графиков (с популяционным весом 6,0, 8,0 и даже 14% от общего количества суммированных аллелей у всех быков-производителей), но они были единичны. А следова- тельно, во всех частотах после 2,2% можно ожидать не более 1-2 антигенов, после 4-5% – только один ан- тиген.
Расчёт частоты аллелей по методике Е. К. Меркурьевой (см. рисунок 10, Г, Д и Е) показал аналогичные тенденции. При этом следует заметить, что у голштинских быков-производителей, семя которых реализовывало ОАО «Орловское» в 2011 году было 38 эритроцитарных антигенов с концентрацией менее 0,05, или 5%, в то время как у голштинских быков-производителей ОАО «ЦСИО» в 2008 году данную концентрацию имели лишь 28 антигенов. Спад количества аллелей с концентрацией от 5 до 20% у них проходил не так интенсивно, как у быков-производителей ОАО «Орловское» по племенной работе, у которых, кроме того, высокую концентрацию имели антигены H', Х 2 , W, Q', А 1 , E, Z, Y 2 , Е' 1 , С 1 , G 2 , L, I 2 , O 1 , S 1 и C 2 .
По частотам эритроцитарных антигенов у чёрнопёстрых голштинизированных коров в ОПХ «Стре- лецкое» можно сделать вывод о том, что, очевидно, коровы одного стада более однообразны в отличие от быков-производителей, семя которых используют при чистопородном разведении и, особенно, при скрещивании, поскольку быки-производители, происходящие из разных стран и разных племенных объединений, могут существенно усиливать генетическую изменчивость в стадах не только отдельных областей, но и целых регионов.
A1
S1
J/1
Y/
P/2
G/
L
Z
M
I
C/
O/
J
X1
I//1
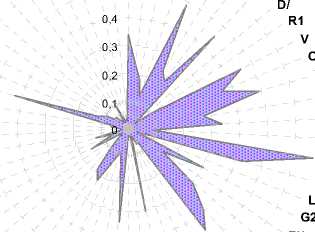
0,4
0,2
D/
R1
V
Y3Q/
O2C1
K/C//
A/2H//
A/2 T1 V/ H//
G3 B/ O1 U1 W K
U//
Рисунок 11. - Концентрация эритроцитарных антигенов у чёрно-пёстрых голштинизированных коров племенного ядра ОПХ «Стрелецкое» (2010 г.)
Также из рисунка 11 следует, что при расчёте по методике Е. К. Меркурьевой (1991) наибольшую концентрацию в ОПХ «Стрелецкое» (33 коровы-рекордистки) имели эритроцитарные антигены H' – 0,666 (66,6%), А' 1 и Y 2 – 0,4848, Х 2 – 0,4545, С'' – 0,4545 и I'' – 0,4242. Антиген F был у всех коров-рекордисток.
Анализ каталога быков-производителей фирмы «Semex» показал интересные результаты (см. таблицу 9). Так, у дочерей быков с генотипом ВВ индекс племенной ценности (EBV) был достоверно выше, чем у коров с генотипом АА, на 469 кг по удою и на 19 кг по молочному жиру (p<0,001 и p<0,01). При этом коэффициенты фенотипической изменчивости (Сv) индекса племенной ценности у быков большинства генотипов были достаточно велики – до 64-68%. По 1 голове в каталоге имели генотип ВЕ и ЕЕ по каппа-казеину.
Таблица 9. - Молочная продуктивность дочерей голштинских быков канадской фирмы «Semex» в зависимости от генотипа по гену каппа-казеина
|
С S о |
m о ю й |
>s О Ч |
Индекс племенной ценности, EBV |
|||||
|
Удой, кг |
С v |
Жир, кг |
С v |
Белок, кг |
С v |
|||
|
АА |
80 |
162786 |
+1046±75 |
64 |
+37±3 |
68 |
+32±2 |
52 |
|
АВ |
35 |
42029 |
+1069±104 |
57 |
+36±4 |
62 |
+37±3 |
51 |
|
ВВ |
2 |
262 |
+1515±40*** |
4 |
+40±9 |
32 |
+51±5** |
15 |
|
АЕ |
8 |
20831 |
+859±186 |
61 |
+47±7 |
42 |
+31± 5 |
47 |
|
ВЕ |
1 |
56 |
+1275 |
0 |
+41 |
0 |
+41 |
0 |
|
ЕЕ |
1 |
39 |
+511 |
0 |
-12 |
0 |
+15 |
0 |
Примечание: **–p<0,01, ***–p<0,001.

АА АВ ВВ А В
Рисунок 12. – Частота генотипов и аллелей каппа-казеина у швицких быков-производителей немецкого происхождения («Spermex Top Sires 2010»)
Из рисунка 12 следует, что у швицких быков-производителей, семя которых было предложено для реализации в немецком каталоге «Spermex Top Sires 2010», генотип ВВ имел наибольшую частоту – 0,474 (или 47,4%), в то время как на генотип АА приходилось всего 0,105 (или 10,5%). Частота аллеля В при этом достигала 0,685, т. е. 68,5%. Это было обусловлено, в первую очередь, комбинированным направлением продуктивности швицкой породы.
Таблица 10. – Молочная продуктивность дочерей швицких быков-производителей, семя которых реализуется в Германии (2010 год)
|
С S о |
m о И |
>s л У о |
Племенная ценность, +/– |
||
|
Удой, кг |
Жир, кг |
Белок, кг |
|||
|
АА |
4 |
342 |
596,5±99,7 |
24,7±4,3 |
22,0±4,7 |
|
АВ |
16 |
1577 |
811,6±87,2 |
32,2±2,9 |
26,2±1,9 |
|
ВВ |
18 |
47432 |
698,6±70,9 |
21,3±3,2 |
24,5±1,8 |
|
Все |
38 |
49351 |
735,4 |
26,3 |
24,9 |
Из таблицы 10 следует, что дочери швицких быков-производителей с генотипами АВ и ВВ превосходили по удоям дочерей с генотипом АА на 215,1 и 102,1 кг молока (в первом случае p<0,1). Превосходство по количеству молочного белка по этим генотипам составило 3,8 и 2,5 кг соответственно. Большое количество дочерей (n=47432) позволило сделать вывод о высокой ценности животных швицкой породы с генотипом ВВ.
Таким образом, на основании проведённых исследований можно предложить схему, которая позволит учитывать при составлении родительских пар прямые и косвенные факторы, влияющие на эффективность подбора по молочной продуктивности и селекционно-технологическим качествам дойных коров (см. рисунок 13). Полученные данные позволяют выделить наиболее важные генетические и внешние факторы, на которые следует обращать внимание при модернизации подбора в стадах.
Повышение эффективности подбора на увеличение признаков молочной продуктивности и селекционно-технологических качеств
т
|
Генетические факторы |
Негенетические факторы |
1 |
1 |
Оценка эффективности подбора родительских пар по селекционным признакам в стадах
Рисунок 13. – Схема повышения эффективности подбора молочного скота
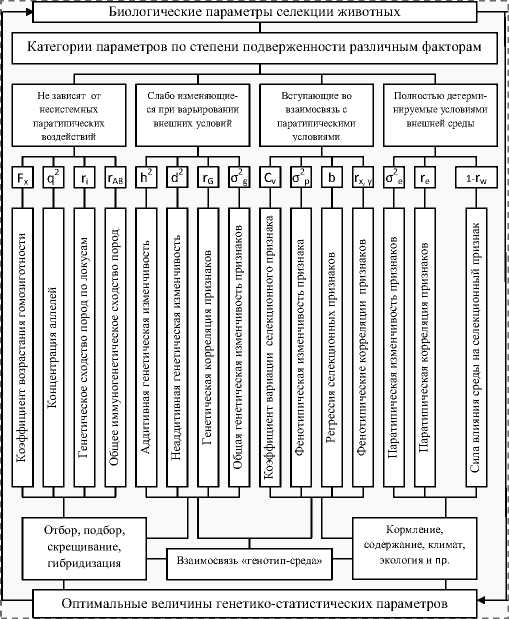
Рисунок 14. – Схема управления биологическими параметрами селекции в популяциях ности генетическим и внешним факторам в четыре категории (см. рисунок 14).
Согласно этим исследованиям, можно выделить параметры, полностью зависящие от генетических факторов, слабо изменяющиеся при варьировании внешних условий, активно вступающие в взаимодействие со средой и зависящие только от внешних условий. На каждый из перечисленных в схеме параметров существуют силы воздействия, а следовательно, на эффективность селекции.
Выводы
В целом, проведённые исследования позволяют подвести итог и сделать вывод, что в группах коров, в разной степени превышающих своих матерей и их средние величины признаков, аддитивная и неаддитивная наследуемость может существенно отличаться, однако структура генетической изменчивости удоя и жирности молока находятся в зависимости друг от друга. В стадах со средними удоями коров на уровне 4000-4500 кг молока целесообразен гетерогенный подбор по удою, при этом удой матерей отцов при составлении родительских пар должен превышать удой матерей на 3000-5000 кг молока, однако при слишком большой разнице при подборе желаемого результата не будет. Также не следует осеменять коров-рекордисток семенем быков с потенциалом их матерей на уровне 4000-4500 кг молока, поскольку это неизбежно приведёт к падению средних удоев в будущем поколении. Особое внимание следует уделять закреплению к стадам быков-улучшателей, т. к. они могут способствовать положительной генетической корреляции между величиной удоев, жирности молока и др. признаков. Существуют предпосылки наследования гинекологических заболеваний и яловости у чёрно-пёстрых коров, а весенний сезон оплодотворения может способствовать большей вероятности рождения телят-двоен. Не исключено, что в стадах чёрно-пёстрых коров следует интенсивно выращивать и осеменять тёлок при живой массе более 420 кг, в результате чего у них уменьшится % мёртворожденных телят, а также будет рождаться равное количество тёлочек и бычков. В стадах не следует применять нецеленаправленного инбридинга, т. к. инбредные тёлки практически всегда уступают аутбредным по интенсивности роста. Обязательным для современных племзаводов и репродукторов должно быть тестирование дойного стада по группам крови и локусу каппа-казеина. Правильно организованный подбор на закрепление генотипа ВВ, ассоциирующегося с высокой сыропригодностью молока, может способствовать повышению удоев. Подробный генетикостатистический анализ в стадах позволит вести селекцию молочного скота более эффективно и добиваться удоев 6000-7000 кг молока.
Кроме того, наши предыдущие исследования [13] позволяют сделать общение биологических параметров и сформировать их по степени подвержен- хозяйственных животных. Вестник Орёл ГАУ. 2012; 6:53-63.
Список литературы Совершенствование системы оценки генетических и средовых факторов при составлении родительских пар в молочном скотоводстве
- Айсанов З. М. Определение эффекта инбридинга у крупного рогатого скота молочных и комбинированных пород Вестник РАСХН. 2004; 5:19-21.
- Бакай А.В. Кочиш И. И., Скрипниченко Г. Г. Генетика М.: КолосС, 2007; 448 с.
- Кузнецов В. М. Разработка оптимальных программ селекции в молочном скотоводстве. Зоотехния. 1996; 1:5-13.
- Винничук Д. Парадоксы инбридинга Молочное и мясное скотоводство. 2003; 5:18-22.
- Воронина Е., Стрекозов Н., Абрампальский Ф., Абылкасымов Д. Влияние вариантов подбора коров на их молочную продуктивность Молочное и мясное скотоводство. 2007; 4:8-9.
- Мымрин В. К вопросу о гетерогенности подбора Молочная и мясное скотоводство. 2006; 4:22-24.
- Сельцов В. И., Сермягин А. А. Продуктивные качества инбредных и аутбредных коров симментальской породы. Зоотехния. 2011; 10:2-4.
- Чернушенко В., Комошенков А., Бабичева В. Тип подбора родителей по ЕАВ-локусу групп крови и хозяйственно-биологические свойства дочерей. Молочное и мясное скотоводство. 2009; 2:9-10.
- Шендаков А. И. Оценка эффективности отбора скота чёрно-пёстрой породы по молочной продуктивности. Вестник ОрёлГАУ. 2010; 6: 93-100.
- Шендаков А. И. Модернизация селекции в молочном скотоводстве Орловской области. Молочное и мясное скотоводство. 2008; 6:15-19.
- Шендаков А. И., Шендакова Т. А. Генетические аспекты модернизации молочного скотоводства. Вестник Орёл ГАУ. 2009; 2:30-35.
- Шендаков А. И. Влияние быков-производителей и типов их подбора на воспроизводительные качества чёрно-пёстрого и симментальского скота. Проблемы биологии продуктивных животных. 2011; 4 (спецвыпуск):159-162.
- Шендаков А. И. Результаты комплексной оценки биологических параметров в селекции сельскохозяйственных животных. Вестник Орёл ГАУ. 2012; 6:53-63.
- Daetwyler H. D., Daetwyler H., Villanueva В., Bijma Р., Woolliams J. Inbreeding in genome-wide selection. J. Anim. Breed. Genet. 2007; 124:369-376.
- Dal Zotto R., De Marchi M., Dalvit C., Cassandro M., Gallo L. Heritabilities and genetic correlations of body condition score and calving interval with yield, somatic cell score, and linear type traits in Brown Swiss cattle. J. Dairy Sci. 2007; 90:5737-5743.
- Parland, S. Mc., Parland S. Mc., Kearney J. F., Rath M., Berry D. P. Inbreeding effect on milk production, calving performance, fertility, and conformation in Irish Holstein-Friesians. J. Dairy Science. 2007; 90:4411-4419.
- Pryce J.E., Coffey M.P., Brotherstone S. The genetic relationship between calving interval, body condition score and linear type and management traits in registered Holsteins. J. Dairy Sci. 2000; 83:2664-2671.
- Thompson J.R., Everett R.W., Wolfe C.W. Effects of inbreeding on production and survival in Jerseys. J. Dairy Sci. 2000; 83:2131-2138.
- VanRaden P.M., Smith L.A. Selection and mating considering expected inbreeding of future progeny. J. Dairy Sci. 1999; 82:2771-2778.
- Veerkamp R.F. Brotherstone S. Genetic correlations between linear type traits, food intake, live weight and condition score in Holstein Friesian dairy cattle. Anim. Sci. 1997; 64:385-392.
- Weigel K.A., Lin S. W. Controlling inbreeding by constraining the average relationship between parents of young bulls entering AI progeny test programs. J. Dairy Sci. 2002; 85:2376-2383.
- Wiggans G.R., Van Raden P.M., Zuurbier J. Calculation and use of inbreeding coefficients for genetic evaluation of United States dairy cattle. J. Dairy Sci. 1995; 78:1584-1590.
