Создание пентагенной полимутантной по жирнокислотному составу линии подсолнечника ЛГ 50

Автор: Чебанова Ю.В., Демурин Я.Н.
Рубрика: Селекция, семеноводство и биотехнология сельскохозяйственных растений
Статья в выпуске: 2 (206), 2026 года.
Бесплатный доступ
В мире созданы генотипы подсолнечника, несущие мутации, влияющие на повышенное содержание пальмитиновой, стеариновой и олеиновой жирных кислот в различных комбинациях. Однако линии, в генотипе которой содержались бы фенотипически проявляемые мутантные аллели всех пяти генов (p1, p2, es1, es2 и Ol), в настоящее время нет. В результате проведенного исследования получены эмпирические доказательства положения биохимической генетики о мутационных блоках в последовательных этапах биосинтеза жирных кислот с накоплением вещества-предшественника, а также верифицирована гипотеза о возможности получения полимутанта с фенотипической экспрессивностью наследственных изменений в семенах. Создана пентагенная полимутантная линия ЛГ50, в масле семян которой содержится около 11,53 % пальмитиновой, 17,19 % стеариновой, 64,21 % олеиновой и 2,16 % линолевой кислот, а также около 33 % суммы насыщенных жирных кислот.
Подсолнечник, селекция, мутация, жирные кислоты, качество масла
Короткий адрес: https://sciup.org/142248081
IDR: 142248081 | УДК: 633.854.78:575 | DOI: 10.25230/2412-608X-2026-2-206-29-34
Development of the sunflower line LG 50: the pentagenic and polymutant line in terms of fatty acid composition
Sunflower genotypes that carry mutations leading to increased levels of palmitic, stearic and oleic fatty acids in various combinations have been developed worldwide. However, there are currently no lines whose genotype contains phenotypically expressed mutant alleles of all five genes (p1, p2, es1, es2, and Ol). The research provided empirical evidence for the biochemical genetics’ theory of mutational blocks in successive stages of fatty acid biosynthesis involving the accumulation of a precursor substance. It also verified the hypothesis that it is possible to obtain a polymutant with phenotypic expression of hereditary changes in seeds. The pentagenic polymutant line LG50 has been developed; its seed oil contains approximately 11.53% palmitic acid, 17.19% stearic acid, 64.21% oleic acid and 2.16% linoleic acid, as well as around 33% total saturated fatty acids.
Текст научной статьи Создание пентагенной полимутантной по жирнокислотному составу линии подсолнечника ЛГ 50
Научная статья УДК 633.854.78
Введение. В семенах традиционного подсолнечника содержится около 85–90 % ненасыщенных жирных кислот, основными из которых являются олеиновая и линолевая, и 10–15 % насыщенных жирных кислот, из которых большую долю занимают пальмитиновая и стеариновая. С помощью химического мутагенеза и обработки семян подсолнечника гамма-излучением в разных странах были индуцированы мутации высокопальмитиновости и высо-костеариновости и созданы линии подсолнечника с высоким содержанием пальмитиновой (HP275, CAS-5, Р-20, CAS-12, CAS-37, ЛГ30 [1–6]) и стеариновой (CAS-8, CAS-4, S-30 и CAS-3 [2; 3], CAS-14 [7; 8], CAS-19 и CAS-20 [9], CAS-29, CAS-30, CAS-15 и CAS-33 [10]) кислот.
Испанские ученые изучали также возможность объединения мутаций высоко-пальмитиновости у CAS-5 и высокостеа-риновости у CAS-3 и установили, что локусы, контролирующие признак высокого содержания C16:0, оказывают эпистатиче-ское влияние на локусы, ответственные за признак высокого содержания C18:0. В результате фенотипическая комбинация, содержащая одновременно высокий уровень пальмитиновой (C16:0) и стеариновой (C18:0) кислот была невозможна. Однако в поколении F3 были выявлены фенотипы с содержанием насыщенных жирных кислот до 44 % (34,5 % C16:0 + 9,5 % C18:0) [11; 12]. В результате скрещивания мутантных линий CAS-5 и CAS-3 были созданы линии CAS-18 (33,2 % C16:0 + 11,1 % C18:0 на линолевом фоне) и CAS-31 (8,8 % C16:0 + 31,9 % C18:0 на линолевом фоне). Линия CAS-25, отобранная из потомства от скрещивания линий CAS-3 и CAS-12, содержала до 30,1 % С16:0 и 6,8 % С18:0, тогда как олеиновой кислоты было 45,8 % [13].
Таким образом, ранее не удавалось создать линии, в масле семян которых было бы повышенное содержание пальмитиновой, стеариновой и олеиновой жирных кислот.
Целью данной работы являлась экспериментальная проверка возможности рекомбинационного объединения в одном генотипе фенотипически проявляемых мутантных аллелей пяти генов ( p1 , p2 , es1 , es2 и Ol ), контролирующих жирнокислотный состав масла подсолнечника.
Материалы и методы. Исследования проведены в лаборатории генетики отдела селекции и первичного семеноводства подсолнечника ФГБНУ ФНЦ ВНИИМК в период 2023–2025 гг. Опыты закладывали в полевых условиях на 2-м отделении в х. Октябрьском г. Краснодара и в камере гидропоники. Посев семян подсолнечника на опытных делянках проводили с помощью ручных сажалок при расстановке 70 × 35 см. В условиях камеры гидропоники выращивали растения подсолнечника при искусственном освещении при фотопериоде 16 часов – день, 8 часов – ночь. В качестве субстрата использовали маты из минеральной ваты; полив осуществляли сбалансированным питательным раствором с помощью специализированной установки полива. Дневная температура в среднем составляла 24– 28 °С, ночная – 18–20 °С. Влажность воздуха поддерживалась на уровне 50–60 %.
В данном исследовании использовали две исходные линии генетической коллекции ВНИИМК: высокостеариновую среднеолеиновую ЛГ36 (LPHSMO) и высокопальмитиновую высокостеариновую ЛГ51 (HPHSLO).
Гибридизацию линий выполняли с использованием ручной кастрации под индивидуальными изоляторами в полевых условиях 2023 г. Растения F 1 принудительно самоопылили в условиях камеры гидропоники (ГП3). Семена F 2 анализировали по составу жирных кислот с использованием метода «прижизненного» анализа. Всего в поколении F 2 проанализировали 145 отдельных семянок. Половинки семян после анализа высевали в камере гидропоники (ГП4). Семена каждого последующего поколения получали самоопылением растений предыдущей генерации.
Лабораторные анализы выполняли в лаборатории генетики на базе экспериментального оборудования лаборатории биохимии. Состав десяти жирных кислот (насыщенных: пальмитиновой, стеариновой, арахиновой, бегеновой и лигноцери-новой; ненасыщенных: пальмитолеиновой, олеиновой, линолевой, линоленовой и эйкозеновой) анализировали с использованием газожидкостной хроматографии метиловых эфиров [14; 15].
Результаты и обсуждение. Имея теоретические и экспериментальные доказательства сложного характера наследования признаков высокого содержания пальмитиновой, стеариновой и олеиновой жирных кислот, данная работа потребовала бы анализа огромной выборки в F 2 отдельных семян при пентагибридном скрещивании. В этом случае было целесообразно применить вариант ступенчатых скрещиваний. В связи с чем приняли решение сократить количество участвующих в расщеплении генов, в первую очередь рецессивных. Поэтому в гибридизационную пару были подобраны родительские линии ЛГ36 и ЛГ51, уже гомозиготные по признаку высокостеариновости.
В поколении F2 искомым генотипом был HPHSMOLL, т.е. отбирали семянки с одновременно повышенным содержанием пальмитиновой (>10 %), стеариновой (>10 %), а также пониженным содержанием линолевой (<10 %) и повышенным или средним – олеиновой кислоты. В результате в опыте в условиях гидропоники из 145 семянок только три относились к данному типу. Для получения семян поколения F3 высеяли все проанализированные «половинки» семянок в пробки из минеральной ваты в камере гидропоники. Часть из них оказалась нежизнеспособной, проросли только 117 семянок. Были получены потомства 84 растений F2.
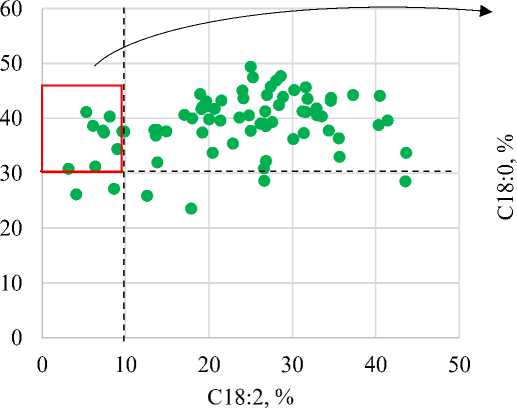
Первыми критериями для отбора в поколении F3 были сумма насыщенных жирных кислот более 30 % и низкое содержание линолевой кислоты (менее 10 %), что косвенно указывает на наличие мутации высокоолеиновости (рис. а). Такими признаками обладали семена 10 семей, для которых был проведен отбор по содержанию пальмитиновой и стеариновой жирных кислот. Из них только пять семей под номерами 25, 64, 73, 105 и 111 обладали одновременно высоким содержанием пальмитиновой и стеариновой жирных кислот (рис. б). Данные семьи являются уникальными, т.к. в их семенах впервые установлено фенотипическое проявление всех изучаемых мутаций.
Данные семьи показали средний уровень содержания пальмитиновой кислоты (от 10,98 до 14,68 %), высокое – стеариновой (от 18,69 до 22,19 %), повышенное – олеиновой (от 51,06 до 54,87 %), низкое – линолевой (менее 10 %) (табл. 1). Наличие растений с таким составом масла семян доказывает возможность рекомбинации мутаций высокопальмитиновости, высо-костеариновости и высокоолеиновости в одном генотипе и их фенотипического проявления.
I м
а (a)
№105
№111
л* • №25
№73 • №64
0 5 10 15 20
С16:0, %
б (b)
Рисунок – Диаграмма рассеивания значений содержания основных жирных кислот в масле семян F 3 :
а) связь содержания линолевой кислоты (С18:2) и суммы НЖК; б) связь содержания пальмитиновой (С16:0) и стеариновой (С18:0) кислот
Fig. – Graph showing the distribution of the content of essential fatty acids in F 3 seed oil: a) the relationship between linoleic acid (C18:2) content and total essential fatty acid content; b) the relationship between palmitic acid (C16:0) and stearic acid (C18:0) content
Таблица 1
Жирнокислотный состав масла в семенах семей F 3 (камера гидропоники ГП4, 2024 г.)
Table 1
Fatty acid composition of the oil in seeds from the F 3 family (GP4 hydroponics chamber, 2024)
|
№ семьи |
С16:0 |
С16:1 |
С18:0 |
С18:1 |
С18:2 |
С18:3 |
С20:0 |
С20:1 |
С22:0 |
С24:0 |
Сумма НЖК |
|
25 |
14,68 |
0,28 |
21,66 |
51,06 |
8,12 |
0,15 |
1,58 |
0,05 |
2,12 |
0,30 |
40,34 |
|
64 |
14,00 |
0,22 |
18,69 |
54,39 |
7,32 |
0,20 |
1,57 |
0,08 |
3,20 |
0,33 |
37,79 |
|
73 |
11,46 |
0,13 |
21,41 |
52,25 |
9,86 |
0,11 |
1,55 |
0,03 |
2,92 |
0,27 |
37,61 |
|
105 |
10,98 |
0,14 |
22,17 |
54,87 |
7,43 |
0,12 |
1,60 |
0,05 |
2,23 |
0,42 |
37,40 |
|
111 |
12,46 |
0,26 |
21,92 |
54,83 |
6,12 |
0,11 |
1,78 |
0,06 |
2,18 |
0,28 |
38,62 |
Потомства семей № 25, 64 и 73 с искомым сочетанием генов (HPHSMOLL) после самоопыления также показали фенотипическое проявление наличия всех мутаций в одном генотипе. Потомство семьи № 25 (образец № 395/2р1) содержало в среднем 11,53 % С16:0, 17,19 % С18:0, 64,21 % С18:1 и 2,16 % С18:2 (табл. 2). Данный образец выделен в отдельную линию генетической коллекции ЛГ50. Состав жирных кислот этой линии значительно отличается от исходных родительских ЛГ36 и ЛГ51, несущих только одну из мутаций, влияющих на содержание насыщенных жирных кислот. Кроме того, в отличие от стандартных линий линолевого (ВК101) и высокоолеинового (ВК1-клп) типа, ЛГ50 содержит 33,11 % суммы насыщенных жирных кислот, что в 2,5–
-
3,5 раза выше. Линия с подобным жирнокислотным составом масла получена впервые в мире.
Заключение. Таким образом, в рамках разработки концепции биохимической генетики липидов о накоплении прекурсоров при генетических блоках в последовательных этапах биосинтеза жирных кислот впервые в мире экспериментально получена пентагенная полимутантная линия ЛГ50, обладающая фенотипическим проявлением всех известных мутантных аллелей состава жирных кислот ( p1 , p2 , es1 , es2 и Ol ). Факт рекомбинации мутаций высо-копальмитиновости, высокоолеиновости и высокостеариновости удалось не только установить в расщепляющейся популяции, но и закрепить в последующих ин-бредных поколениях.
Таблица 2
Жирнокислотный состав масла в семенах линий генетической коллекции ВНИИМК (полевые условия, 2025 г.)
Table 2
Fatty acid composition of the oil in seeds from the lines of the genetic collection of V.S. Pusto-voit All-Russian Research Institute of Oil Crops (field conditions, 2025)
|
Линия |
С16:0 |
С16:1 |
С18:0 |
С18:1 |
С18:2 |
С18:3 |
С20:0 |
С20:1 |
С22:0 |
С24:0 |
Сумма НЖК |
|
ЛГ50 |
11,53 |
0,27 |
17,19 |
64,21 |
2,16 |
0,11 |
1,62 |
0,14 |
2,36 |
0,41 |
33,11 |
|
ЛГ36 |
4,59 |
0,02 |
22,04 |
65,71 |
2,45 |
0,14 |
1,85 |
0,16 |
2,65 |
0,38 |
31,51 |
|
ЛГ51 |
21,03 |
0,46 |
14,25 |
15,17 |
45,48 |
0,06 |
1,17 |
0,05 |
1,95 |
0,38 |
38,79 |
|
ВК1-клп (st) |
4,11 |
0,22 |
2,50 |
87,83 |
3,23 |
0,11 |
0,31 |
0,36 |
0,84 |
0,49 |
8,25 |
|
ВК101 (st) |
6,38 |
0,10 |
5,30 |
33,23 |
53,16 |
0,06 |
0,39 |
0,12 |
0,94 |
0,32 |
13,33 |
