Сравнительное изучение устойчивости к септориозу и физиологических показателей у разных сортов пшеницы

Автор: Яруллина Любовь Георгиевна, Касимова Рита Ильшатовна, Шпирная Ирина Андреевна, Ахатова Альбина Рашитовна, Ибрагимов Ринат Исмагилович
Журнал: Известия Самарского научного центра Российской академии наук @izvestiya-ssc
Рубрика: Экология
Статья в выпуске: 3-5 т.15, 2013 года.
Бесплатный доступ
Проведено сравнительное изучение ряда показателей (экспрессионная активность генов анионной пероксидазы и оксалатоксидазы, а также активность пектиназ и их ингибиторов). у растений пяти сортов пшеницы, которые по предварительным данным различались по устойчивости к заражению патогенными грибами. По всем функциональным признакам устойчивые сорта превосходили чувствительные. Полученные данные указывают на перспективность использования таких признаков для отбора устойчивых форм растений.
Устойчивость к патогенам, экспрессия генов, оксалатоксидаза, пероксидаза, ингибиторы пектиназ, пектиназы
Короткий адрес: https://sciup.org/148202038
IDR: 148202038 | УДК: 581.1
Comporative research of resistance to septoria and physiological features in different wheat variety
Comparative research of number of indicators (gene expression of anion peroxidase and oxalate oxidase and activity of pectinase and pectinase inhibitors) was carried out on five wheat variety, which had different resistance to infection by pathogenic fungi. Resistant varieties were superior to sensitive for all functional attributes. The findings show perspective of using these features for selection of resistant plant forms.
Текст научной статьи Сравнительное изучение устойчивости к септориозу и физиологических показателей у разных сортов пшеницы
В естественных условиях растения испытывают на себе действие разнообразных неблагоприятных факторов как биотической, так и абиотической природы, и их выживание зависит от способности защитить себя от всех этих воздействий. С точки зрения интересов человека, важным аспектом устойчивости растений является сохранение их урожайности на фоне взаимодействия с различными патогенами. Важным подходом к изучению взаимодействия механизмов, обеспечивающих устойчивость к биотическим стрессам, может быть сравнение связанных с ними физиологических показателей у разных сортов пшеницы.
В настоящее время доказано, что важную роль во взаимоотношениях растений и патогенов играют активные формы кислорода (АФК), в том числе перекись водорода (Н 2 О 2 ) [1-3]. АФК не только непосредственно ингибируют рост патогена, накапливаясь в высоких концентрацией в зоне поражения, но участвуют в ускорении лигнификации и других модификациях клеточной стенки, обеспечивающих усиление их барьерной функции, а также индуцируют экспрессию защитных генов. Ок-салатоксидаза и пероксидазы играют важную роль в генерации АФК при заражении [4, 5]. Их активация и сопряженная генерация Н 2 О 2 под воздействием салициловой кислоты и хитоолигосахаридов повышали устойчивость пшеницы к возбудителям твердой головни и корневой гнили [6, 7]. Роль ок-салатоксидазы в защите от грибных патогенов определяется также тем, что при ее участии разрушаются продуцируемые грибами оксалаты, являющиеся агентами, направленными против защитных реакций растений.
Мы провели сравнительное изучение ряда по-
казателей у растений пяти сортов пшеницы, которые по предварительным данным различались по устойчивости к заражению патогенными грибами. В лабораторных условиях устойчивость растений к фитопатогенам оценивали по степени проявления поражения листьев при их инфицировании возбудителем септориоза. Одновременно оценивали ряд признаков, связанных с формированием устойчивости растений к инфицированию. В качестве показателей, имеющих отношение к защите растений от заражения, мы оценивали экспрессионную активность генов анионной пероксидазы и оксала-токсидазы, а также активность пектиназ и их ингибиторов. Как известно, эти белки относят к связанным с патогенезом (PR proteins). Изучение этих показателей не только могло прояснить их роль в формировании устойчивости растений, но и проверить возможность их использования в процессе отбора устойчивых форм растений.
Исследования проводили в лабораторных условиях на растениях мягкой пшеницы ( Triticum aesti-vum, сорта Башкирская 26, Казахстанская 10, Омская 35, Омская 36, Симбирка). Семена проращивали в темноте в течение 2-3 сут на дистиллированной воде. 3-суточные проростки пересаживали на разбавленный раствор Кнопа (1:2) и выращивали на светоплощадке с фотопериодом 16 ч/сут.
Для заражения листьев и оценки их устойчивости к фитопатогену полностью развернутые листья 7-суточных проростков срезали, помещали во влажную камеру, срезы прикрывали ватой, смоченной в растворе бензимидазола (40 мг/л) [8]. Листья выдерживали при комнатной температуре в темноте в течение 24 ч, переносили на искусственное освещение с фотопериодом 16 ч/сут. Отрезки листьев инокулировали суспензией пикноспор Sep-toria nodorum Berk. (106 спор/мл), которые были выделены авторами из местной популяции гриба. Инокулированные листья выдерживали при комнатной температуре в темноте в течение 24 ч, переносили на искусственное освещение с фотопе- риодом 16 ч/сут.
Интенсивность развития гриба на эпидермисе листьев оценивали в течение 120 ч, экспрессию генов оксалатоксидазы и пероксидазы через 24 и 48 ч после инокуляции. В качестве контроля использовали неинфицированные листья растений.
Тотальную РНК выделяли с помощью тризола (Molecular Research Center, Inc, США). Для получения кДНК на основе мРНК проводили реакцию обратной транскрипции с использованием M-MuLV обратной транскриптазы (Fermentas, Литва), согласно протоколам фирм-поставщиков. Концентрацию ДНК и РНК измеряли при А260/A280 на спектрофотометре Smart SpecTM Plus (Bio-Rad), предварительно растворив образцы в Тris-EDTA буфере. Перед амплификацией концентрации всех образцов выравнивались.
Для анализа экспрессии генов, кодирующих пероксидазу и оксалатоксидазу, с помощью программы «Primer Select» (DNAStar) были подобраны высокоспецифичные праймеры.
Праймеры к анионной пероксидазе T.aestivum :
Anper (F) 5’-ttc-gac-aag-cag-tac-tac-cac-aa- 3’
Anper (R) 5’-ccg-aag-tcc-gag-aag-aac-tg- 3’ которые фланкируют консервативный участок гена пероксидазы размером 220 п.н.
Праймеры к оксалатоксидазе T.aestivum:
OxO (F) 5’-atg-act-tcc-tct-tct-cgt-cca-ag- 3’
OxO (R) 5’-gga-gct-gaa-gag-tgt-caa-tgg- 3’, которые фланкируют консервативный участок гена ок-салатоксидазы размером 410 п.н.
В качестве положительного контроля использовали реакцию амплификации гена, кодирующего конститутивно экспрессирующийся белок «домашнего хозяйства» ADP-RF (house-keeping gene) пшеницы:
F 5’-gac- cac-cat-cct-cta-caa-g-3’
R 5’-agc-agc-aca-gca-tca-c-3’
Отрицательным контролем служила реакция с добавлением всех реагентов за исключением кДНК. Уровень экспрессии гена оценивали по интенсивности свечения полос, полученных при электрофоретическом разделении ПЦР-продуктов в 1-2% агарозном геле или 7% ПААГ, на анализаторе гелей («Bio-Rad», США) с помощью компьютерной программы TotalLab. В качестве маркеров использовали фрагменты ДНК фага l после расщепления рестрикта-зами Bgl I и MassRuler DNA Ladder, Low Range (Fermentas, Литва).
Активность пектолитических ферментов определяли использую гелевые пластины, приготовленные по [9], с яблочным пектином в 0.2M ацетатном буфере (pH 5.6) в качестве субстрата. Раствор пектина смешивали с раствором агарозы в ацетатном буфере и затем добавляли ацетат кальция. В конечном итоге, концентрация ингредиентов в смеси была следующей: агароза 1.5%, пектин 1%, ацетат кальция 0.5%. Для определения активности пектиназ и их ингибиторов листья растирали с кварцевым песком и экстрагировали дистиллированной водой в соотношении 1:5 в течение 2 ч при 40С. Экстракт фильтровали через фильтровальную бумагу и центрифугировали дважды при 3500 g в течение 10 мин. 20 μl супернатанта добавляли в лунки гелевых пластин. Для осуществления гидролиза иммобилизованного пектина кюветы с гелевыми пластинами прикрывали крышкой и инкубировали 9 ч при 370C. Действие пектиназ оценивали, обрабатывая пластины 10% ацетатом меди. Области геля, содержащие гидролизованный пектин, проявлялись как светлое пространство на голубом фоне. Активность фермента рассчитывали, измеряя размер области геля с гидролизованным субстратом вокруг лунок. Активность фермента, катализирующего гидролиз субстрата в области геля размером 1 мм2 принимали за условную миллиединицу (mU). Для определения активности ингибиторов пектиназ растительный экстракт смешивали с равным объемом 100 μg ml-1 пектиназы из Aspergillus niger (Sigma, USA) и добавляли в лунки. Активность ингибитора рассчитывали как разницу между значениями для 50 μg ml-1 пектиназы Aspergillus niger и смеси пектиназы с растительным экстрактом. Количество ингибитора, уменьшающего активность фермента на 1 mU, принимали за 1 миллиединицу ингибиторной активности (mIU).
Опыты проводили в 3 биологических повторах. В каждом варианте опыта фиксировали по 5 листьев. Компьютерный анализ аминокислотных и нуклеотидных последовательностей проводили с помощью пакета компьютерных программ Lasergene фирмы “DNASTAR, Inc.” (США). Статистическую обработку проводили по стандартным программам. Данные на графиках представляют собой средние величины и их стандартные ошибки.
Наблюдение за ростом возбудителя септориоза на эпидермисе листьев разных сортов пшеницы выявило различия в степени развития гриба. Так, на листьях пшеницы сорта Казахстанская 10 уже через 24 ч после инфицирования в местах нанесения спор появлялись слабые хлорозы и некрозы. Признаки развития септориоза на листьях растений сорта Омская 36, Симбирка, Башкирская 26, Казахстанская 10 наблюдались через 48 ч после инфицирования в виде слабого обесцвечивания листьев с последующим их побурением, а через 72 ч после инокуляции симптомы септориоза проявлялись в виде бурых пятен с пушистым налетом мицелия. В листьях растений сорта Омская 35 в указанные сроки симптомы болезни были слабо выражены.
В таблице 1 изученные сорта ранжированы по интенсивности развития возбудителя септориоза S. nodorum , которую оценивали по площади инфекционного пятна спустя 120 ч после инокуляции. Как видно из таблицы 1, листья пшеницы сорта Казахстанская проявляли наиболее выраженные симптомы заболевания, в то время как у сорта Омская 35 они были выражены в наименьшей степени.
Таблица 1 . Площадь инфекционного пятна в листьях пшеницы различных сортов пшеницы
|
Cорта пшеницы с различной полевой устойчивостью |
Площадь инфекционного пятна, мм2 |
|
Омская 35 |
7,3 ± 0,3 |
|
Омская 36 |
9,1± 0,4 |
|
Симбирка |
12,6±0,9 |
|
Башкирская 24 |
15,9 ±1,1 |
|
Казахстанская 10 |
19,6 ±1,3 |
Таким образом, наибольшую устойчивость к инфицированию возбудителем септориоза проявляли растения пшеницы сорта Омская 35, наименьшую – сорта Казахстанская 10. Сорта пшени- цы Омская 36, Симбирка и Башкирская 26 занимали по степени инфицируемости растительных тканей патогенным грибом S. nodorum промежуточное положение.
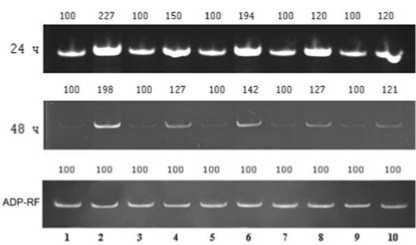
Рис. 1 . Экспрессия гена анионной пероксидазы (TC 151917) в листьях пшеницы разных сортов при инфицировании S. nodorum ( S. n .) через 24 и 48 ч.
1 – Омская 35, 2 – Омская 35 + S. n. , 3 – Омская 36, 4 – Омская 36 + S. n., 5 – Симбирка, 6 – Симбирка + S. n., 7 – Башкирская 26, 8 – Башкирская 26 + S. n., 9 – Казахстанская 10, 10 – Казахстанская 10 + S. n.
Заражение индуцировало повышение уровня экспрессии гена анионной пероксидазы в листьях растений сорта Омская 35, Омская 36 и Симбирка (рис. 1). В наибольшей степени это проявлялось у сорта Омская 35. Из сравнения данных, полученных через 24 и 48 ч после инокуляции, видно, что со временем уровень экспрессии продолжал возрастать у растений пшеницы сортов Омская 35 и 36
и снижался у сорта Симбирка. В листьях растений сортов Башкирская 26 и Казахстанская 10 достоверных изменений экспрессии гена анионной пероксидазы под влиянием заражения не было выявлено.
Заражение повышало экспрессию гена оксала-токсидазы у всех изученных сортов (рис. 2).

Рис. 2 . Экспрессионная активность гена оксалатоксидазы (AJ556991.1) в инфицированных и контрольных листьях растений пшеницы сортов различной устойчивости. 24 и 48 ч после инфицирования. Обозначения как на рис. 1
Хотя со временем уровень экспрессии этого гена в основном снижался, он оставался выше, чем в контроле. Экспрессия была максимальной в листьях растений сорта Омская 35 и минимальной у сортов Башкирская 26 и Казахстанская 10.
В отсутствие заражения различия между сортами по пектолитической активности были небольшими (табл. 2). Максимально высокое значение у сорта Омская 35 лишь в 1.2 раза превышало минимальный уровень у сорта Симбирка.
Таблица 2 . Активность пектиназ в инфицированных и контрольных листьях растений пшеницы различных сортов
|
Сорт |
Активность пектиназ в контрольных растениях, Е/ г |
Активность пектиназ в инфициров. растениях, Е/ г |
|
Омская 35 |
109 ± 5,45 |
87 ± 4,35 |
|
Омская 36 |
99 ± 4,95 |
68 ± 3,4 |
|
Симбирка |
89 ± 4,45 |
104 ± 5,2 |
|
Башкирская 26 |
104 ± 5,2 |
117 ± 5,85 |
|
Казахстанская 10 |
94 ± 4,7 |
103 ± 5,15 |
При заражении различия между сортами увеличились, и соотношение между максимальным уровнем у растений сорта Башкирская 26 и минимальным у сорта Омская 36 было уже 1.6. Это возрастание различий было следствием того, что у одних сортов (Симбирка, Башкирская 26 и Казахстанская 10) пектолитическая увеличилась, что могло быть следствием ферментативной активности патогена, а у других (Омская 35 и 36) – снизилась. Возможно, в результате повышения активности ингибитора пектиназ (рис. 3).
В отсутствие заражения у растений сорта Казахстанская 10 активность ингибитора пектиназы была достоверно ниже, чем у всех остальных сортов, и хотя она возрастала при заражении до уровня, зафиксированного в листьях растений сорта Башкирская 26, была ниже, чем у всех изученных сортов. В результате заражения активность ингибитора возрастала в листьях пшеницы сорта Омская 35 и снижалась у сорта Симбирка. Эти вызванные заражением изменения в уровне активности ингибитора привели к появлению сильной отрицательной корреляция между этим показателем и тяжестью заболевания растений (r=-0.95).
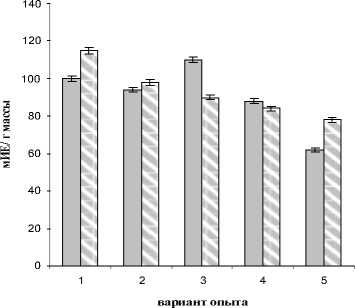
□ Контроль
□ Инфицирование
Рис. 3. Активность ингибитора пектиназы в растениях пшеницы различных сортов при инфицировании S. nodo- rum.
1 – Омская 35, 2 – Омская 36, 3 – Симбирка, 4 – Башкирская 26, 5 – Казахстанская 10
Таким образом, в нашей работе уровень повышения экспрессии гена оксалатоксидазы коррелировал с устойчивостью растений к грибным патогенам. Уровень экспрессии гена пероксидазы резко возрастал при заражении устойчивых растений (Омская 35 и 36) и не менялся у чувствительных к заражению растений сортов Башкирская 26 и Казахстанская 10. Представляет интерес то, что индуцируемый заражением уровень экспрессии гена ок-салатоксидазы достигал максимума раньше, чем пероксидазы (через 24 и 48 ч после заражения соответственно). Эти результаты указывают на более раннее проявление роли оксалатоксидазы.
Таким образом, по всем перечисленным функциональным признакам (уровень индукции экспрессии генов пероксидазы и оксталатоксидазы, активность пектиназ и их ингибиторов на фоне заражения) устойчивые сорта превосходили чувстви- тельные, что указывает на перспективность использование таких признаков для отбора устойчивых форм растений.
Работа проведена при финансовой поддержке гранта РФФИ_поволжье_а № 11-04-97037 и ФЦП ГК № 01201353578.
-
1. Shirasu K., Schulze-Lefert P. Regulators of cell death in disease resistance // Plant Mol. Biol. 2000. V. 44. N 3. P. 371385.
-
2. Тарчевский И.А., Чернов В.М. Молекулярные аспекты фитоиммунитета // Микология и фитопатология. 2000. Т. 34. Вып. 3. С. 3-10.
-
3. Ramputh A.I., Arnason J.T., Cass L., Simmonds J.A. Reduced herbivory of the European corn borer ( Ostrinia nubi-lalis ) on corn transformed with germin, a wheat oxalate oxidase gene // Plant Sci. 2002. V. 162. P. 431-440.
-
4. Яруллина Л.Г., Трошина Н.Б., Максимов И.В., Хайруллин Р.М. Участие оксалатоксидазы в неспецифичной защит-
ной активации окисления ортофенилендиамина в проростках пшеницы при стрессе // Агрохимия. 2003. № 12. С. 55-59.
-
5. Максимов И.В., Валеев А.Ш., Черепанова Е.А., Яруллина Л.Г. Продукция активных форм кислорода в листьях пшеницы, инфицированных разновирулентными штаммами S. nodorum Berk. // Прикладная биохимия и микробиология. 2009. Т. 46. № 4. С. 481-486.
-
6. Трошина Н.Б., Яруллина Л.Г., Валеев А.Ш., Максимов И.В. Индукция салициловой кислотой устойчивости пшеницы к Septoria nodorum Berk. // Известия РАН. 2007. № 5.
-
7. Бурханова Г.Ф., Яруллина Л.Г., Максимов И.В. Пути регуляции хитоолигосахаридами защитных реакций в рас-
тениях пшеницы при инфицировании Bipolaris sorokiniana // Физиология растений. 2007. Т. 54. С. 119126.
-
8. Пыжикова Г.В., Карасева Е.В. Методика изучения возбудителей септориоза на изолированных листьях пшеницы // С.-х. биология. 1986. №.12. С.112-114.
-
9. Шпирная И.А., Умаров И.А., Шевченко Н.Д., Ибрагимов Р.И. Определение активности гидролаз и их ингибиторов с использованием субстратов, иммобилизованных в геле агарозы // Прикладная биохимия и микробиология. 2009. Т. 45. № 4. С. 497-501.
COMPORATIVE RESEARCH OF RESISTANCE TO SEPTORIA AND PHYSIOLOGICAL FEATURES IN DIFFERENT WHEAT VARIETY
Список литературы Сравнительное изучение устойчивости к септориозу и физиологических показателей у разных сортов пшеницы
- Shirasu K., Schulze-Lefert P. Regulators of cell death in disease resistance//Plant Mol. Biol. 2000. V. 44. N 3. P. 371-385.
- Тарчевский И.А., Чернов В.М. Молекулярные аспекты фитоиммунитета//Микология и фитопатология. 2000. Т. 34. Вып. 3. С. 3-10.
- Ramputh A.I., Arnason J.T., Cass L., Simmonds J.A. Reduced herbivory of the European corn borer (Ostrinia nubilalis) on corn transformed with germin, a wheat oxalate oxidase gene//Plant Sci. 2002. V. 162. P. 431-440.
- Яруллина Л.Г., Трошина Н.Б., Максимов И.В., Хайруллин Р.М. Участие оксалатоксидазы в неспецифичной защитной активации окисления ортофенилендиамина в проростках пшеницы при стрессе//Агрохимия. 2003. № 12. С. 55-59.
- Максимов И.В., Валеев А.Ш., Черепанова Е.А., Яруллина Л.Г. Продукция активных форм кислорода в листьях пшеницы, инфицированных разновирулентными штаммами S. nodorum Berk.//Прикладная биохимия и микробиология. 2009. Т. 46. № 4. С. 481-486.
- Трошина Н.Б., Яруллина Л.Г., Валеев А.Ш., Максимов И.В. Индукция салициловой кислотой устойчивости пшеницы к Septoria nodorum Berk.//Известия РАН. 2007. № 5.
- Бурханова Г.Ф., Яруллина Л.Г., Максимов И.В. Пути регуляции хитоолигосахаридами защитных реакций в растениях пшеницы при инфицировании Bipolaris sorokiniana//Физиология растений. 2007. Т. 54. С. 119-126.
- Пыжикова Г.В., Карасева Е.В. Методика изучения возбудителей септориоза на изолированных листьях пшеницы//С.-х. биология. 1986. №.12. С.112-114.
- Шпирная И.А., Умаров И.А., Шевченко Н.Д., Ибрагимов Р.И. Определение активности гидролаз и их ингибиторов с использованием субстратов, иммобилизованных в геле агарозы//Прикладная биохимия и микробиология. 2009. Т. 45. № 4. С. 497-501.
