Сравнительные исследования выхода однонитевых разрывов и щелочнолабильных сайтов ДНК при воздействии 365 нм ультрафиолетового и рентгеновского излучений на лимфоциты периферической крови человека

Автор: Сметанина Н.М., Пустовалова М.В., Бушманов А.Ю., Осипов А.Н.
Журнал: Саратовский научно-медицинский журнал @ssmj
Рубрика: Генетика
Статья в выпуске: 4 т.9, 2013 года.
Бесплатный доступ
Цель; сравнить количественный выход однонитевых разрывов (ОР) и щелочнолабильных сайтов (ЩС) ДНК, индуцированных 365 нм ультрафиолетовым (УФ) и рентгеновским излучениями в лимфоцитах периферической крови человека in vitro. Материал и методы. Выделение лимфоцитов крови человека проводили путем центрифугирования в градиенте плотности фикол-верографин. Облучение клеток проводили на УФ-установке BLX-365 (длина волны 365±10 нм) и рентгеновской биологической установке РУБ РУСТ-М1. Анализ ОР и ЩС ДНК проводили с использованием метода ДНК-комет в щелочных условиях. Результаты. Впервые показано, что выход ОР и ЩС ДНК в лимфоцитах крови человека при воздействии 365 нм УФ излучения в дозе 10 кДж/м2 соответствует эффекту рентгеновского излучения ~ в дозе 1 Гр. Заключение. Воздействие 365 нм УФ излучения в дозе 1 кДж/м2 на лимфоциты периферической крови человека индуцирует -100 ОР ДНК на клетку.
365 нм уф излучение, лимфоциты, метод днк-комет, однонитевые разрывы днк, рентгеновское излучение
Короткий адрес: https://sciup.org/14917842
IDR: 14917842 | УДК: 577.34:
Comparative study of the amount of DNA single-strand breaks and alkali-labile sites in human peripheral blood lymphocytes exposed to 365 nm UV versus X-ray radiation
Aim: to compare the amount of DNA single-strand breaks (SSB) and alkali-labile sites (ALS) induced by DNA 365 nm ultraviolet (UV) versus X-rays radiation in human peripheral blood lymphocytes at in vitro conditions. Material and methods. Blood lymphocytes were isolated in a Ficoll-verografin density gradient. Irradiation of cells was performed using BLX-365 UV-unit (wavelength 365 ± 10nm)and RUSTM1 biological X-ray unit. Analysis of DNA SSB and ALS was carried out using comet assay at alkaline conditions. Results. For the first time it was demonstrated that the amount of SSB and ALS induced by 365 nm UV radiation at a dose of 10 kJ/m2 in human lymphocytes corresponds to the effect of X-rays at a dose of- 1 Gy. Conclusion. Exposure of human peripheral blood lymphocytes to 365 nm UV radiation at a dose of 1 kJ/m2 induces - 100 DNA SSB per cell.
Текст научной статьи Сравнительные исследования выхода однонитевых разрывов и щелочнолабильных сайтов ДНК при воздействии 365 нм ультрафиолетового и рентгеновского излучений на лимфоциты периферической крови человека
1Введение. Основное внимание исследователей биологических эффектов ультрафиолетового (УФ) излучения долгие годы было сосредоточено на коротковолновом (менее 280 нм) излучении, вызывающем серьезные повреждения ДНК и других макромолекул, но почти полностью поглощаемом озоновым слоем Земли. УФ излучение с длинами волн 315–400 нм (согласно международной классификации — УФ-A), более 90% которого достигает земной поверхности, считалось относительно безвредным. В последние десятилетия было показано, УФ-А излучение, обладающее большой проникающей способностью и воздействующее на глубокие слои кожи, может индуцировать развитие злокачественных новообразований [1–2]. В отличие от коротковолнового и средневолнового (280–315 нм), УФ-A излучение слабо поглощается ДНК и индуцирует повреждения ее структуры преимущественного через генерацию активных форм кислорода и азота [3–4].
Большинство работ по изучению биологического действия УФ-А излучения сосредоточенно на эффектах в клетках базального слоя кожи. Однако, в отличие от более коротковолнового УФ излучения, УФ-А излу-
чение проникает в глубокие слои кожи и оказывает воздействие на дерму, которая богата капиллярами. При этом происходит облучение клеток периферической крови, в том числе и лимфоцитов. Тем не менее, до настоящего времени, работы, посвященные изучению ДНК повреждающего действия УФ-А излучения на лимфоциты крови, были единичны [5–7].
Цель нашей работы : исследования количественного выхода однонитевых разрывов (ОР) и щелочнолабильных сайтов (ЩС) ДНК, индуцированных 365 нм УФ излучением в лимфоцитах периферической крови человека в сравнении с выходом этих повреждений при воздействии рентгеновского излучения.
Выбор рентгеновского излучения в качестве агента сравнения был обусловлен хорошей изученностью механизмов его ДНК повреждающего действия.
Материал и методы. Для исследований использовалась кровь физически здоровых мужчин-доноров в возрасте 21–28 лет. Забор периферической крови проводили в K2ЭДТА-вакутейнеры (Vacuette). У всех доноров было получено согласие на проведение данного исследования.
Выделение лимфоцитов крови человека проводили путем центрифугирования в градиенте плотности фикол-верографин (Histopaque, Sigma-Aldrich) в соответствии с прилагаемой инструкцией. После выделения лимфоциты отмывали и ресуспензировали в фосфатно-солевом буфере до конечной концентрации 1x106 клеток/мл.
200 мкл суспензии клеток смешивали с 600 мкл 1 % раствора легкоплавкой агарозы (Thermo Scientific, TopVision Low Melting Point Agarose) в фосфатно-солевом буфере (рН 7,4) при температуре 37,5ºС. 75 мкл полученной смеси наносили на предварительно покрытые слоем 1 % нормоплавкой агарозы предметные стекла, накрывали покровными стеклами и оставляли на 10 мин при 4ºC до образования плотного геля.
Слайды с иммобилизованными в агарозу клетками помещали в холодный (4ºС) фосфатно-солевой буфер (рН 7,4). Жизнеспособность клеток контролировали с помощью двойной флуоресцентной окраски акридиновым оранжевым и йодистым пропидием согласно общепринятой методике [8]. В этих условиях клетки сохраняли жизнеспособность (не более 5% погибших клеток) в течение нескольких часов. Время самих экспериментов не превышало 15 мин.
Облучение клеток рентгеновскими лучами в дозах 1–5 Гр проводили на рентгеновской биологической установке РУБ РУСТ-М1 (Россия) при мощности дозы 0,85 Гр/мин и температуре 4˚C (для охлаждения использовались термогранулы LAB ARMOR BEADS).
Для ультрафиолетового облучения использовали установку BLX-365 (Bio-Link), длина волны 365±10 нм. Облучение проводилось при 4°С в дозах 5–20 кДж/м2. Интенсивность излучения — 2,92 кДж/м2 за 1 мин.
Анализ ОР и ЩС ДНК проводили с использованием метода ДНК-комет в щелочных условиях [9, 10]. Коротко: сразу после облучения (10–15 сек) агарозные слайды переносили в холодный (4˚C) лизирующий буфер (2.5 M NaCl, 100 мM Na2EDTA, 20 мM Tris-HCl, pH 10.0, 1% Triton X-100 и 10% DMSO) и выдерживали в течение 1 ч в темноте при 4˚C. После лизиса клеток слайды помещали в холодный (4˚C) щелочной раствор (300 мM NaOH, 1 мM EDTA, рН>13) и выдерживали 20 мин для расплетания (щелочной денатурации) нитей ДНК. Электрофорез проводили в щелочном буфере при напряжении 0,75 В/см при комнатной температуре в течение 20 мин. После электрофореза проводили нейтрализацию для ренатурации (восстановления нативности) ДНК (3-кратная промывка в 0,4 М Трис-НСl буфере, рН 7.4). Затем слайды слегка подсушивали и фиксировали в 70% этаноле в течение 10 мин. Для окраски ДНК использовали SybrGreen I (Invitrogen). Визуализацию и документирование микроизображений ДНК-комет проводили с помощью микроскопа люминесцентного микроскопа Axioscop-40 FL (Carl Zeiss) и видеосистемы на основе цифровой камеры MRс 5 (Carl Zeiss) с программой AxioVision 4.8 (Carl Zeiss). На каждом слайде регистрировали по 100 комет. Обрабатывали по 3 слайда от каждого донора на экспериментальную точку. Для анализа и обработки ДНК-комет использовали программу СASP 1.2.2 (СASPlab). В качестве критерия поврежденности ДНК использовали % ДНК в хвосте.
Статическая обработка результатов проводилась с использованием пакета статистических программ Statistica 8.0 (StatSoft). Результаты исследований представлены как среднее арифметическое результатов трех независимых экспериментов ± стандартная ошибка.
Результаты. Для того чтобы минимизировать вклад клеточных метаболических процессов (продукция свободных радикалов в митохондриях, эксцизионная репарация ДНК) в наблюдаемые эффекты, облучение клеток проводилось при низкой температуре (4°С), а сам анализ повреждений ДНК выполнялся сразу (10–15 сек) после облучения. Для количественной оценки ОР и ЩС ДНК была использована щелочная версия метода ДНК-комет (метод гель-электрофореза ДНК единичных клеток). Этот метод отличается высокой чувствительностью и позволяет анализировать изменения количества ОР и ЩС ДНК при дозах редкоионизирующего излучения всего 5–10 сГр [11]. После щелочной денатурации (рН>13) ДНК, иммобилизованных в агарозу клеток проводится электрофорез, во время которого петли и фрагменты ДНК мигрируют в геле агарозы к аноду, создавая своеобразный «хвост». Таким образом, % ДНК в хвосте ДНК-комет отражает степень повреж-денности ДНК.
На рис. 1 представлены результаты исследований выхода ОР и ЩС ДНК в лимфоцитах периферической крови человека при их облучении in vitro рентгеновским излучением в дозе 1–5 Гр. Зависимость «доза — эффект» хорошо описывается линейной функцией у =5,94+6,60 x (R2=0,99), где у — % ДНК в хвосте комет, а x — доза облучения в Гр.
При воздействии 365 нм УФ излучения в дозах 5–20 кДж/м2 также наблюдается линейное увеличение % ДНК в хвосте комет во всем изученном диапазоне доз (рис. 2). Зависимость «доза — эффект» для УФ излучения аппроксимируется линейной функцией
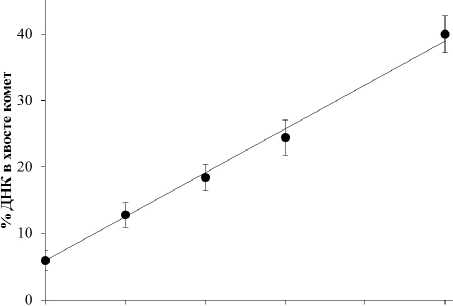
Доза, Гр
Рис. 1. Зависимость изменений относительного количества ОР и ЩС ДНК (% ДНК в хвосте) в лимфоцитах периферической крови человека от дозы рентгеновского излучения
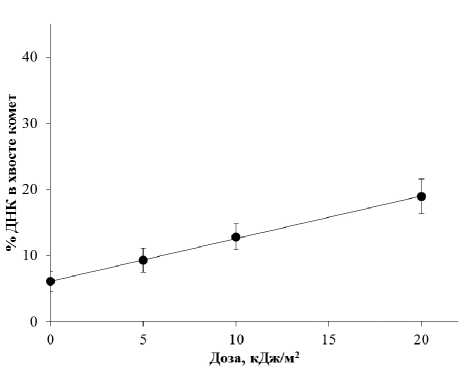
Рис. 2. Зависимость изменений относительного количества ОР и ЩС ДНК (% ДНК в хвосте) в лимфоцитах периферической крови человека от дозы ультрафиолетового излучения с длиной волны 365 нм у=6,08+0,65x (R2=0,99), где у — % ДНК в хвосте комет, а x — доза облучения кДж/м2.
Обсуждение. Ионизирующее излучение повреждает ДНК как при прямом попадании кванта или частицы в молекулу ДНК (непосредственная передача энергии молекуле ДНК и молекулам воды, вплотную прилегающим к ДНК), так и косвенно, индуцируя свободные радикалы (преимущественно HO•), вызывающие главным образом модификацию оснований и однонитевые разрывы ДНК. Известно, что около 70% повреждений ДНК можно предотвратить добавлением низкомолекулярных антиоксидантов, нейтрализующих HO• [12].
В живых клетках энергия УФ-А излучения поглощается различными фотосенсибилизаторами, например эндогенным меланином [13], белками, содержащими порфирины, гемы, хиноны или флавины [14]. В процессе фотоокислительной реакции I типа возбужденный сенсибилизатор принимает электрон (или атом водорода) непосредственно от находящейся вблизи молекулы. По реакции I типа в ДНК происходит в основном окисление наиболее реакционного основания — гуанина с образованием 8-оксо-7,8-дигидрогуанина (8-оксо-гуанина) [15]. В ходе фотоокислительной реакции II типа происходит образование синглетного кислорода (основной путь) и супероксид-анион радикала (минорный путь) [16]. Синглетный кислород, взаимодействуя с ДНК, также в конечном счете индуцирует образование 8-ок-со-гуанина [17], в то время как супероксид-анион в реакциях спонтанной и ферментативной дисмутации образует перекись водорода, которая затем в реакциях Фентона и Габера — Вейса генерирует ОН• радикал, индуцирующий ОР и ЩС ДНК [18]. Таким образом, механизм образования ОР и ЩС ДНК при воздействии УФ-А излучения сходен с механизмом действия редкоионизирующего излучения.
При сравнительном анализе кривых «доза — эффект», полученных при воздействии рентгеновского и 365 нм УФ излучений (см. рис. 1 и 2 соответственно), нами было показано, что эффект воздействия 365 нм УФ излучения в дозе 10 кДж/м2 соответствует эффекту рентгеновского излучения в дозе 1 Гр. Известно, что при воздействии редкоионизирующего излучения в дозе 1 Гр в клетке образуется ~1000 ОР ДНК [19], то есть мы можем предварительно заключить, что воздействие 365 нм УФ излучения в дозе 1 кДж/м2 в расчете на одну клетку индуцирует ~100 ОР ДНК.
Заключение. В результате сравнительных исследований количественного выхода ОР и ЩС ДНК, индуцированных 365 нм УФ излучением и рентгеновским излучением в лимфоцитах периферической крови человека, впервые было показано, что выход ОР и ЩС ДНК при воздействии 365 нм УФ излучения в дозе 10 кДж/м2 соответствует эффекту рентгеновского излучения ~ в дозе 1 Гр. На основании результатов проведенных исследований и литературных данных о выходе ОР ДНК при воздействии редкоио-низирующего излучения сделано заключение о том, что воздействие 365 нм УФ излучения в дозе 1 кДж/м2 индуцирует ~100 ОР ДНК на клетку.
Список литературы Сравнительные исследования выхода однонитевых разрывов и щелочнолабильных сайтов ДНК при воздействии 365 нм ультрафиолетового и рентгеновского излучений на лимфоциты периферической крови человека
- Chronic UVA irradiation of human HaCaT keratinocytes induces malignant transformation associated with acquired apoptotic resistance/Y.Y. He, J. Pi, J.L. Huang [et al.]//Oncogene. 2006. Vol. 25. P. 3680-3688
- Ridley A.J., Whiteside J.R., McMillan T. J., Allinson S. L. Cellular and sub-cellular responses to UVA in relation to carcinogenesis//Int. J. Radiat. Biol. 2009. Vol. 85 (3). P. 177-195
- UV irradiation increases ROS production via PKCdelta signaling in primary murine fibroblasts/O. Bossi, M. Gartsbein, M. Leitges [etal.]//Cell Biochem. 2008. Vol. 105(1). P. 194-207
- Cooper K. L, Liu K.J., Hudson L.G. Enhanced ROS production and redox signaling with combined arsenite and UVA exposure: contribution of NADPH oxidase//Free Radical Biol. Med. 2009. Vol. 47 (4). P. 381-388
- Najafzadeh M., Baumgartner A., Gopalan R. [et al.] In vitro sensitivities to UVA of lymphocytes from patients with colon and melanoma cancers and precancerous states in the micronucleus and the Comet assays//Mutagenesis. 2012. Vol. 27 (3). P. 351-357
- DNA damage after acute exposure of mice skin to physiological doses of UVB and UVA light/A.R. Svobodova, A. Galandakova, J. Sianska [et al.]//Arch. Dermatol. Res. 2012. Vol. 304(5). P. 407-412
- Сметанина H.M., Пустовалова M.B., Осипов A. H. Модифицированный метод ДНК-гало для оценки повреждений ДНК, индуцированных различными генотоксическими агентами//Радиационная биология: Радиоэкология. 2013. Т. 53, № 4. С. 389-393
- Mascotti К., McCullough J., Burger S. R. HPC viability measurement: trypan blue versus acridine orange and propidium iodide//Transfusion. 2000. Vol. 40 (6). P. 693-696
- Оценка молекулярных и цитогенетических эффектов хронического воздействия низкоинтенсивного у-излучения у мышей/А. Н. Осипов, А. Л. Елаков, П. В. Пучков [и др.]//Генетика. 2002. Т. 38, № 10. С. 1345-1350
- Мязин А.Е., Осипов А. Н., Елаков А. Л., Сыпин В. Д., Шевченко В. А. Оценка методом ДНК-комет однонитевых разрывов ДНК лимфоцитов селезенки в отдаленные сроки после острого облучения мышей в сублетальных и среднелетальных дозах//Радиационная биология: Радиоэкология. 2002. Т. 42, вып. 6. С. 731-734
- Comparison in vivo study of genotoxic action of high versus very low dose-rate y-irradiation/A. N. Osipov, D.Y. Klokov, A. L. Elakov [et al.]//Nonlinearity in Biology, Toxicology and Medicine. 2004. Vol. 2, № 3. P. 223-232
- Roots R., Okada S. Protection of DNA molecules of cultured mammalian cells from radiation-induced single strand scissions by various alcohols and SH compounds//Int. J. of Radiat. Biol. 1972. Vol. 21. P. 329-342
- UVcausationofmelanomainXiphophorusisdominated by melanin photosensitized oxidant production/S. R. Wood, M. Berwick, R.D. Ley [et al.]//Proc. Natl. Acad.Sci. USA. 2006. Vol. 103. P. 4111-4115
- Kawanishi S., Hiraku Y. Sequence-specific DNA damage induced by UVA radiation in the presence of endogenous and exogenous photosensitizers//Curr. Probl. Dermatol. 2001. Vol. 29. P. 74-82
- Cadet J., Douki T. Oxidatively generated damage to DNA by UVA radiation in cells and human skin//J. Invest. Dermatol. 2011. Vol. 131 (5). P. 1005-1007
- Ridley A.J., Whiteside J.R., McMillan T. J., Allinson S. L. Cellular and sub-cellular responses to UVA in relation to carcinogenesis//Int. J. Radiat. Biol. 2009. Vol. 85 (3). P. 177-195
- Singlet oxygen induces oxidation of cellular DNA/J. L. Ravanat, P. Di Mascio, G. R. Martinez [et al.]//J. Biol. Chem. 2001. Vol. 276. P. 40601-40604
- Cadet J., Douki T, Ravanat J. L., Di Mascio P. Sensitized formation of oxidatively generated damage to cellular DNA by UVA radiation//Photochem. Photobiol. Sci. 2009. Vol. 8 (7). P. 903-911
- Ward J. F. DNA damage produced by ionizing radiation in mammalian cells: identities, mechanisms of formation, and reparability//Prog. Nucleic Acid Res.Mol. Biol. 1988. Vol. 35. P. 95-125.
