Стимуляция костным мозгом остеогенеза в дистракционном регенерате (экспериментальное исследование)
")
Автор: Шевцов В.И., Ерофеев С.А., Мигалкин Н.С., Осипова Е.В.
Журнал: Гений ортопедии @geniy-ortopedii
Рубрика: Оригинальные статьи
Статья в выпуске: 3, 2003 года.
Бесплатный доступ
Проведены эксперименты на 9 взрослых собаках по стимуляции аутологичным костным мозгом остеогенеза в дистракционном регенерате. Установлено, что после стимуляции, через 7 дней фиксации, в половине опытов регенерат утрачивал зональное строение. Дальнейшая динамика исследования остеогенеза на этапах фиксации показала прогрессивное замещение соединительнотканной прослойки регенерата костными структурами. Выявлено, что формирование и дифференцировка костноостеоидных трабекул регенерата происходит на фоне повышенной плотности остеогенных клеточных элементов и остеобластов. Таким образом, комплекс проведенных исследований свидетельствует о стимулирующем влиянии аутологичного костного мозга, введенного в соединительнотканную прослойку дистракционного регенерата, на процессы костеобразования.
Эксперимент, собака, удлинение конечности, стимуляция остеогенеза, костный мозг
Короткий адрес: https://sciup.org/142120615
IDR: 142120615
Stimulation of osteogenesis in the distraction regenerate bone with bone marrow (experimental study)
The experiments in osteogenesis stimulation in the distraction regenerate bone with autologous bone marrow were made using 9 adult dogs. It was revealed that the regenerate bone lost its zonal structure after stimulation, after 7 days of fixation, in half of the experiments. The further dynamics of osteogenesis study at fixation stages demonstrated progressive replacement of the regenerate bone connective-tissue layer with bone structures. Formation and differentiation of the regenerate bone osteoosteoid trabeculae were revealed to occur through increased density of osteogenic cellular elements and osteoblasts. Thus, the complex of the studies made shows stimulating effect of autologous bone marrow, put into the regenerate bone connective-tissue layer, on osteogenesis processes.
Текст научной статьи Стимуляция костным мозгом остеогенеза в дистракционном регенерате (экспериментальное исследование)
Способы уравнивания длины конечностей по методу Илизарова обеспечивают восстановление анатомии, строения и функции костного органа, но требуют много времени для трансформации сформировавшегося дистракционного регенерата в новую кость. Общий срок лечения больных, в зависимости от величины удлинения, колеблется от 4 до 18 месяцев [1, 2]. В последние годы актуализируется потребность в дополнительных воздействиях на процесс дистракционного костеобразования с целью ускорения консолидации, дифференцировки и созревания регенератов с последующим сокращением сроков лечения больных, в том числе при вялотекущем остеогенезе.
Для ускорения перестройки новообразованной костной ткани в РНЦ "ВТО" имени академика Г.А.Илизарова был предложен и экспериментально апробирован ряд механических способов "воспитания" регенерата [3-5]. Этот метод используют украинские ортопеды, которые для ускорения перестройки регенерата при помощи специально разработанных устройств применили управляемые механодинамические воздействия [6].
Для стимуляции органотипической перестройки новообразующейся костной ткани используются различного рода медикаментозные препараты [ 7, 8]. Однако в ряде случаев их применение не дает желаемого эффекта, что затягивает сроки лечения больных.
Определенный эффект при стимуляции дистракционного регенерата был достигнут при использовании белковых фракций, выделенных из костной ткани или сыворотки крови животных в период активного остеогенеза, и отсутствовал при введении в прослойку регенерата физиологического раствора [9-11].
Одним из аспектов проблемы стимуляции остеогенеза является применение костного моз- га. Известны исследования, в которых с целью стимуляции костеобразования использовался непосредственно костный мозг, в том числе и облученный ультрафиолетовыми лучами или монохромным красным светом [12]. Для лечения больных с ложными суставами и дефектами применяли аутотрансплантацию костномозговых фибробластов человека, выращенных in vitro [13]. При изучении заживления повреждений костей [14] было установлено, что использование костномозговых аутоэксплантатов определяло более раннее образование остеобла- стической ткани в регенерате. Однако в литературе нет данных по применению костного мозга для стимуляции дистракционного остеогенеза, за исключением упоминания об использовании инъекции костного мозга для заживления кости при слабом костеобразовании после удлинения конечности [15].
Цель настоящего исследования – разработка и апробация предложенной В.И. Шевцовым методики стимуляции дистракционного остеогенеза аутологичным костным мозгом [18], изучение динамики процессов костеобразования.
МАТЕРИАЛ И МЕТОДЫ ИССЛЕДОВАНИЯ
Эксперименты проведены на 9 взрослых беспородных собаках (возраст 1 - 3 года), которым в условиях операционной на голень накладывали аппарат Илизарова из четырех опор и производили пилкой Джигли открытую поперечную остеотомию берцовых костей в средней трети. Удлинение голени начинали через 3 дня после операции. Режим дистракции составил 1 мм в сутки за 4 приема, по 0,25 мм в течение рабочего времени. Удлинение осуществляли 28 дней. Период последующей фиксации отломков продолжался 7, 14, 21 и 28 дней.
По окончании дистракции, после рентгенографии удлиненной голени, под наркозом, с помощью иглы Кассирского из проксимального метафиза плечевой кости получали пунктат костного мозга (КМ) в объеме 3 мл. В пробирку с пунктатом добавляли в том же объеме питательный солевой раствор № 199 и центрифугировали в течение 5 минут. Затем из пробирки забирали осадочную взвесь костного мозга (2-3 мл) и вводили ее в прослойку дистракционного регенерата.
Таким образом, условия дистракционного остеосинтеза были сознательно "ужесточены" для снижения активности остеогенеза, на фоне которого стимулирующий эффект костного мозга проявился бы наиболее отчетливо.
Содержание, оперативные вмешательства и выведение животных из опыта осуществляли согласно приказу МЗ СССР № 755, 1977 г.
Компьютерный анализ изображений рентгенограмм выполнен на аппаратно-программном комплексе "ДиаМорф" (Пакет прикладных программ анализа изображений ДиаМорф-Cito. Версия 1.1. 1996. Москва). Для выделения в тенях регенератов структур, имеющих разную рентгеновскую плотность и, соответственно, степень минерализации, использовали операцию интерактивной 3-х уровневой сегментации. Измеряли площади, занимаемые высоко-, средне, слабо- и неминерализованными структурами [16]. Полученные цифровые данные обрабатывали в программе Microsoft Excel - 97.
Для гистологического исследования регенерата большеберцовую кость фиксировали в 10% растворе формалина, затем выпиливали блок, включающий регенерат и концы отломков. Гис-тотопографические срезы окрашивали гематоксилином и эозином, пикрофуксином по Ван-Гизону.
Для оценки клеточного состава трансплантата костного мозга из пунктата и его центрифуга-та готовили мазки, которые окрашивали по способу Паппенгейма. Цитологическое исследование трансплантата костного мозга (КМ) показало, что центрифугат представлен преимущественно зрелыми эритроцитами, в числе немногочисленных малодифференцированных клеток преобладали молодые формы эритропоэтического ряда. Определялись также метамиелоциты и палочкоядерные нейтрофилы (рис. 1). После разведения питательной средой и центрифугирования отмечено изменение тона окраски ядер и цитоплазмы, а также снижение четкости ядер-ных контуров в части клеток (рис. 2).
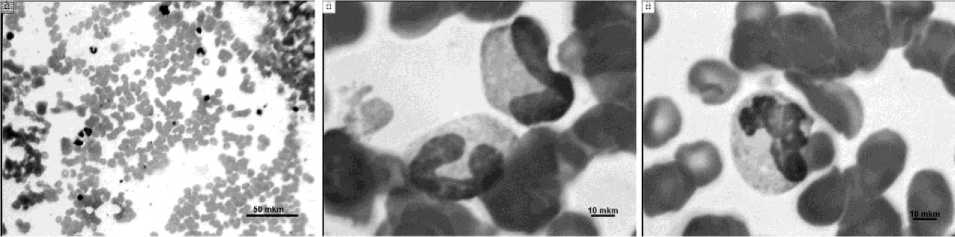
а б в
Рис. 1. Мазок костного мозга: а – обзор; б – палочкоядерные и в – миелоцитарные формы миелогенеза. Окраска по Паппенгейму
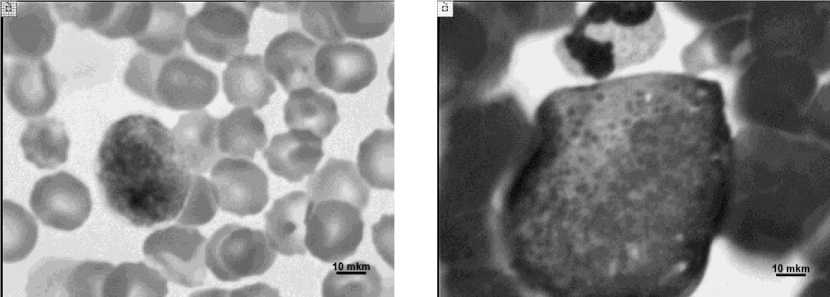
Рис. 2. Снижение четкости ядерных контуров после подготовки аутотрансплантата костного мозга. Окраска по Паппенгейму
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
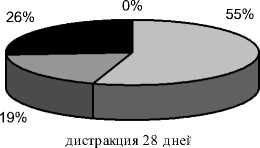
Через 28 дней дистракции (9 собак, рис. 3а) диастаз был равен 27,0±0,22 мм. Регенерат, заполнявший диастаз, имел типичное зональное строение и продольно исчерченную структуру. Его поперечник в 2-х опытах был равен, в 4-х -меньше на 1,0-2,0 мм, в 3-х случаях - больше на 1,0-2,0 мм поперечника прилежащих концов отломков. Протяженность костных отделов составляла: проксимального - 12,8±0,88 мм, дистального - 8,3±0,3 мм. Высота разделяющей их прослойки колебалась от 3,1±0,4 мм до 8,3±1,3 мм. В большинстве опытов ее пересекали единичные нитевидные трабекулярные тени. По периферии костных отделов регенерата только в 5 опытах определялись признаки формирующейся корковой пластинки. Также в 5 случаях произошло слияние эндо- и периостальных теней регенерата, причем в 2 -х опытах регенерат имел эндостальное происхождение и определялся только в проекции костномозговой полости. Доля неминерализованных структур в регенерате составляла 26,11±2,56%, слабоминерализованных - 19,25±1,56%. Суммарно слабо- и неминерализованные структуры занимали от 33% до 59% площади регенерата, соотношения этих структур в большинстве случаев были примерно 1:1. На долю среднеминерализованных структур приходилось 54,24±2,92% (рис. 4а).
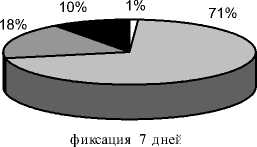
Через 7 дней фиксации (после окончания дистракции и стимуляции остеогенеза; рис. 3б.) в большинстве опытов поперечник регенерата был больше поперечника отломков на 1,0-4,0 мм. В 4-х опытах регенерат утрачивал зональное строение, на месте прослойки определялась либо полоса уплотнения, либо чередовались участки уплотнения и просветления. В других 5 случаях протяженность проксимального костного отдела практически не изменялась, а дистального - увеличивалась до 12,0±1,2 мм. Высота прослойки в этих опытах в основном была 1,0-3,0 мм. Ее сплошь пересекали трабекулярные тени, слившиеся в костные «мостики». Структура регенерата в основном была продольно исчерченной, а в 2-х случаях приобрета- ла гомогенность. Также в 2-х опытах формирующаяся корковая пластинка с 3-х сторон полностью перекрыла весь диастаз. В остальных наблюдениях она либо прерывалась на уровне прослойки, либо вообще определялась на одном из костных отделов регенерата. К этому сроку происходило уменьшение долей неминерализованных (9,68±2,84%, p<0,01) и слабоминерализованных структур (18,21±2,93%, p>0,001) и одновременное увеличение долей средне-(71,54±5,16%, p<0,01) и высокоминерализованных структур (0,57±0,13%, p>0,001). В сумме слабо- и неминерализованные структуры составляли 3-51%, высоко- и среднеминерализованные - 49-97% (рис. 4б). При этом высокоминерализованные структуры были выявлены у 4 животных.
Гистологически на 7 сутки (2 случая) после введения аутотрансплантата КМ в исследованных препаратах отмечено замещение срединной прослойки костноостеоидными структурами. В одном опыте определялась более выраженная консолидация с почти полным замещением прослойки костноостеоидными трабекулами как в эндостальной, так и в интермедиарной проекциях объема регенерата (рис. 5а). В другом - отмечено некоторое смещение прослойки в дистальном направлении. В участках замещения прослойки расположены продольно ориентированные костноостеоидные трабекулы. Особенность срединной прослойки регенератов на этом сроке состояла в высокой плотности расположения ядер фибробластов. При этом, в некоторых участках определялось ритмичное их расположение с формированием муарового рисунка, что обусловлено разным характером накопления масс межклеточного матрикса. Следует отметить, что глыбки матрикса в вышеописанных участках по характеру окраски и аморфности сходны с остеоидом. Формирующиеся костноостеоидные трабекулы вблизи прослойки также характеризовались высокой плотностью расположения клеток как входящих в состав трабекул, так и остеогенных элементов, расположенных в меж- трабекулярных пространствах.
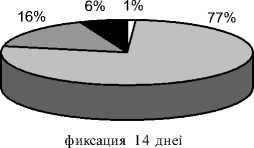
Через 14 дней фиксации (7 собак, рис. 3в) в 3-х опытах не определялось зонального строения регенерата, который приобретал гомогенную структуру. На месте прослойки определялась полоса более плотной тени. Новообразованная корковая пластинка с 2-4-х сторон полностью перекрывала диастаз. В остальных 4-х опытах величина костных отделов увеличилась незначительно. Прослойка выявлялась в виде отдельных очагов просветления либо была замещена на 1/3 и более трабекулярными тенями. Регенерат в подавляющем большинстве имел бочкообразную форму. Корковая пластинка формирующегося по периферии регенерата только в 3-х опытах с 2-х сторон полностью перекрывала диастаз. Отмечали увеличение доли высоко- и среднеминерализованной костной ткани и уменьшение доли слабо- и неминерализованных структур (рис. 4в). Гистологически в исследованных регенератах явления консолидации с замещением прослойки продольноориентированными костноостеоидными трабекулами более выражены в сравнении с предыдущим сроком (рис. 5б). В структурах срединной прослойки регенерата продолжали выявляться участки многоклеточной компоновки остеогенных элементов и остеобластов с отложениями остеоида и формированием примитивных трабекул.
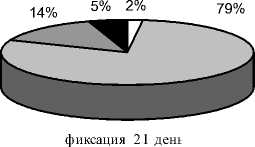
Через 3 недели фиксации (5 собак, рис. 3г) поперечник регенерата по-прежнему преимущественно был больше на 3 мм прилежащих концов отломков. В 2-х опытах к этому сроку диастаз был полностью замещен костной тканью. На месте бывшей прослойки определялся участок уплотнения. У 3-х собак еще прослеживалась зональность регенерата. При этом прослойка не выглядела сплошной зоной просветления. Ее интенсивно пересекали костные мостики, исходящие из костных отделов регенерата. В результате прослойка представляла собой чередование участков уплотнения и просветления. Последние были высотой не более 3,0 мм. В 4-х опытах структура регенерата была гомогенной. Новообразованная корковая пластинка в 3-х опытах с 2 - 3-х сторон полностью перекрывала диастаз. В регенерате преобладали высоко- и среднеминерализованные структуры, составляющие в среднем соответственно 2,32±0,94% (p<0,05) и 78,54±5,74% (p<0,01) и занимающие от 63% до 97% площади регенерата (рис. 4г). На долю неминерализованных структур приходилось не более 3% и только в одном случае -17%. На гистологических препаратах (рис. 5в) консолидация регенератов сопровождалась выраженным формированием субпериостальной костной манжетки. Остаточные структуры соединительнотканной прослойки содержали поля хондроидной трансформации.
В течение 28 дней фиксации наблюдали 3
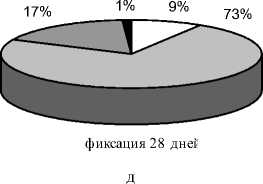
собаки. У всех животных регенерат сохранял бочкообразную форму (рис. 3д). Лишь только в одном опыте к этому сроку сохранялись признаки зональности регенерата, причем высота прослойки, которая определялась в виде отдельных участков просветления, не превышала 1,0 - 2,0 мм. У оснований регенерата увеличились зоны просветления - формировалась костномозговая полость, а в одном из опытов новообразованная костномозговая полость сливалась с костномозговой полостью отломков. Регенерат, как правило, с 2-3-х сторон был перекрыт тенями новообразованной корковой пластинки, толщина которой к этому сроку достигала 3,0 мм. Через 28 дней фиксации доля высокоминерализованных структур увеличивалась до 8,52±2,89% (p<0,05). По сравнению с 21 днем фиксации незначительно изменялось содержание средне- и слабоминерализованных структур. Неминерализованные структуры составляли в среднем 0,56±0,39% (p<0,01) от площади регенерата (рис. 4д). На 28 сутки после стимуляции гистологически в исследованных регенератах определялась выраженная консолидация густой сетью анастомозирующих трабекул без продольной ориентации, в межтрабекулярных пространствах - рыхлая волокнистая соединительная ткань с очагами миелоидной ткани. В одном случае сохранялась тонкая остаточная структура прослойки, почти полностью замещенная хрящевой тканью. В некоторых участках хрящевые поля имели строение гиалинового (рис. 5г).
Следует отметить, что к концу эксперимента было отмечено формирование костного футляра вокруг питательной артерии, поврежденной на операции. Вероятно, происходит ее реканализация в связи с хорошей адаптацией концов отломков на операции и отсутствием их смещений на этапах удлинения конечности в подавляющем числе опытов.
Таким образом, комплекс проведенных исследований свидетельствует о стимулирующем влиянии аутологичного костного мозга, введенного в соединительнотканную прослойку дистракционного регенерата, на процессы костеобразования. При этом следует отметить, что к концу дистракции высота прослойки достигала 8 мм, ее доля в регенерате составляла 26%. А уже через 7 дней фиксации (после стимуляции) в половине опытов регенерат утрачивал зональное строение. В других случаях высота соединительнотканной прослойки в основном была 1,0-3,0 мм и доля неминерализованных структур составляла 10%. Выявлено, что формирование и дифференцировка костноостеоидных трабекул регенерата происходит на фоне повышенной плотности остеогенных клеточных элементов и остеобластов. Активизированный фенотип остеогенных клеток и остеобластов, выражающийся в смещении ядер к одному из полюсов, более светлой окраске ядер за счет преобладания эухроматина, наличии 2-3 ядрышек и в повышении объема вакуолизированной цитоплазмы, может свидетельствовать об усилении клеточных биосинтетических процессов. Дальнейшая динамика исследования остеогенеза на этапах фиксации показала прогрессивное замещение соединительнотканной прослойки регенерата костными структурами. Так, через 3 недели фиксации доля неминерализованных структур регенерата уменьшилась до 5-6%, доля среднеминерализованных структур увеличивалась до 77-79% в сравнении с их показателями в 55% к концу дистракции. Через 28 дней фиксации у всех животных регенерат имел форму, близкую к бочкообразной. Только в одном опыте к этому сроку соединительнотакнная прослойка регенерата определялась в виде отдельных участков просветления и не превышала 1,0-2,0 мм. При этом к концу дистракции ее высота в этом случае достигала 10 мм, что соответствовало параметрам, выявленным ранее при удлинении с однократной дробностью. В этих опытах через месяц фиксации прослойка оставалась значительной по высоте, а период фиксации для ее полного замещения достигал 90-150 дней [17].
Таким образом, в арсенал ортопедов-травматологов предлагается новый эффективный способ стимуляции дистракционного остеогенеза, не требующий дорогостоящего оборудования и материалов.
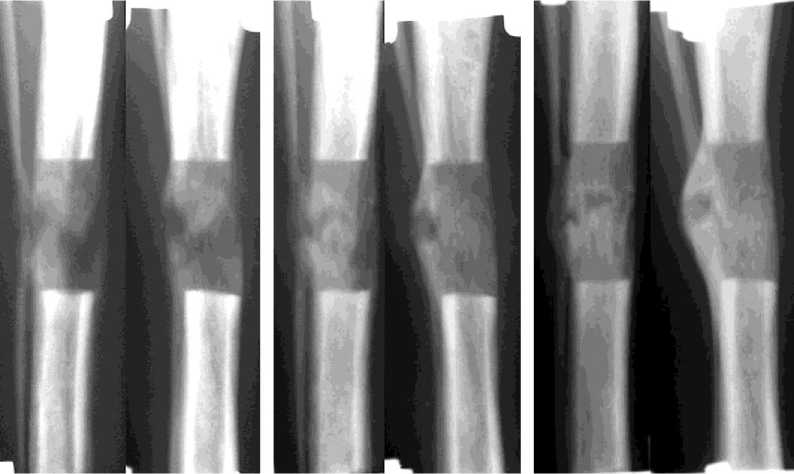
а
б
в
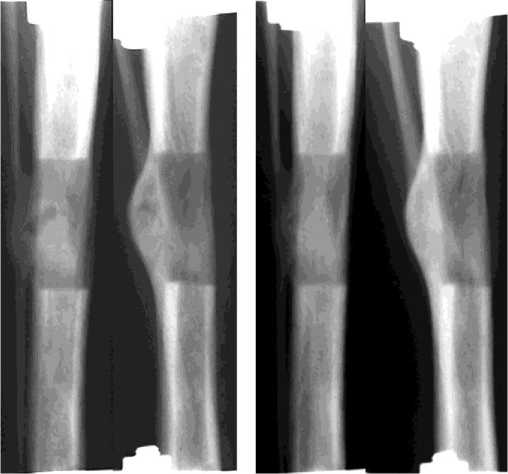
г д
Рис. 3. Рентгенограммы дистракционного регенерата до (а – дистракция 28 дней) и после стимуляции: б – фиксация 7 дней; в – фиксация 14 дней; г – фиксация 21 день; д – фиксация 28 дней
а
г
б
в
-
- высокоминерализованные структуры
-
- среднеминерализованные структуры
-
- слабоминерализованные структуры
-
- неминерализованные структуры
Рис. 4. Доли структур различной рентгеновской плотности в площади дистракционного регенерата на этапах эксперимента (а – дистракция 28 дней; б – фиксация 7 дней; в – фиксация 14 дней; г – фиксация 21 день; д – фиксация 28 дней)
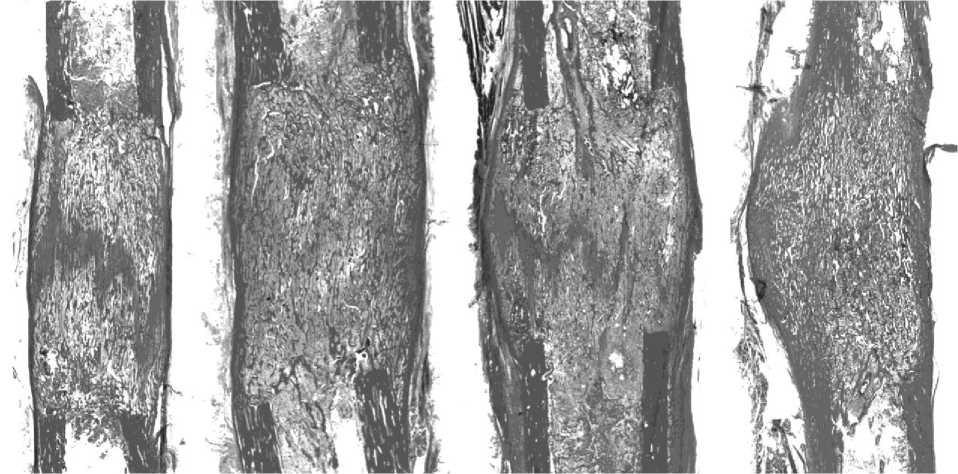
а б в г
Рис. 5. Гистологическая динамика костеобразования на обзорных гистотопограммах дистракционного регенерата после стимуляции остеогенеза костным мозгом (а – фиксация 7 дней, б – 14 дней, в – 21 день и г – 28 дней фиксации). Окраска пикрофуксином по Ван-Гизону. Ув. лупное
