STR-генотипирование осетровых рыб, выращиваемых в УЗВ: значение и перспектива применения

Автор: Гинаятов Н.С., Бейшова И.С., Ульянов В.А., Ульянова Т.В., Ковальчук А.М., Сидарова А.Ж.
Статья в выпуске: 3 т.255, 2023 года.
Бесплатный доступ
В статье приведены результаты исследований по оценке полиморфизма по 7 микросателлитным локусам ДНК осетровых рыб, в которых идентифицировано 69 аллелей. Все исследуемые особи были гетерозиготными, соответственно, уровень наблюдаемой гетерозиготности равнялся 1. На основании полученных результатов оценки полиморфизма STR-локусов у сибирских осетров, выращиваемых в УЗВ, установлено, что в исследуемой группе соотношение ожидаемой и наблюдаемой гетерозиготности, а также показателя индекса фиксации (Fis), говорит об избытке гетерозигот в них и высоком «запасе» генетического разнообразия по STR-локусам.
Рыбы, осетр, генотипирование, str, узв
Короткий адрес: https://sciup.org/142238904
IDR: 142238904 | УДК: 639.371/.374, | DOI: 10.31588/2413_4201_1883_2_255_116
STR-genotyping of sturgeon fish grown in the RAS: significance and prospects of application
The article presents the results of studies on the assessment of polymorphism in 7 microsatellite DNA loci of sturgeon fish, in which 69 alleles were identified. All the studied individuals were heterozygous, respectively, the level of observed heterozygosity was 1. Based on the results of the evaluation of STR-loci polymorphism in Siberian sturgeons grown in the RAS, it was found that in the study group, the ratio of expected and observed heterozygosity, as well as the index of fixation (Fis), indicates an excess of heterozygotes in them and a high "reserve" of genetic diversity in STR-loci.
Текст научной статьи STR-генотипирование осетровых рыб, выращиваемых в УЗВ: значение и перспектива применения
Генетическая идентификации осетровых рыб в условиях индустраильной аквакультуры обусловлена необходимостью при формировании пар для воспроизводства, т.к. при разведении осетровых на установках замкнутого водоснабжения (УЗВ) имеется риск близкородственого скрещивания и как следствие – инбридинг, приводящий к снижению продуктивности и плодовитости рыб [3, 5]. Для изучения генетической структуры осетровых рыб в качестве генетического маркера является микросателлиты, ввиду широкой распространенности их по всему геному, и служат инструментом для высокоэффективного генетического картирования и характеристики таксонов, установления филогенетических связей, уточнения конспецифических гибридизаций, идентификации посадочного материала [4, 8]. Следовательно, оценка полиморфизма STR-локусов у осетров, выращиваемых в условиях УЗВ, послужила целью исследований, для достижения которой определены задачи:
– Охарактеризовать аллелофонд популяций осетровых рыб, выращиваемых в УЗВ;
– Рассчитать популяционногенетические характеристики по микросателлитным маркерам;
– Определить степень генетического разнообразия в популяции осетровых рыб.
Материал и методы исследований. В качестве исходного материала для исследования послужила ДНК, выделенная из фрагментов грудных плавников, отобранных от ремонтно- маточного поголовья осетровых рыб, выращиваемых в УЗВ. Объектом исследований послужили 20 особей сибирского осетра (Acipenser baerii) выращиваемых на базе лаборатории ихтиологии и аквакультуры ЗападноКазахстанского аграрно-технического университета имени Жангир хана. Лабораторные исследования проведены в лаборатории биотехнологии и диагностики инфекционных болезней Испытательного центра Западно-Казахстанского аграрнотехнического университета имени Жангир хана.
ДНК было экстрагировано из тканей плавников с использованием коммерческого набора «ДНК-Экстран-2», который позволяет качественно выделить геномную ДНК с высоким выходом и чистотой. Для оценки генетического полиморфизма были использованы 7 микросателлитных локусов (Таблица 1) [11].
В системе капиллярного электрофореза «АBI 3500 Genetic аnаlyzer» проводилось электрофоретическое разделение продуктов амплификации и с применением программного обеспечения GeneMаpper определены длины аллелей.
При анализе учитывались только те фрагменты, которые располагались в ожидаемом аллельном диапазоне и соответствовали границам локуса. Все фрагменты, локализованные за пределами крайних аллелей данного локуса, не подлежали к дальнейшему анализу.
Популяционно-генетические характеристики были определены по формулам:
Наблюдаемая гетерозиготность вычислялась по М. Нею [9]:
Ho = 1 / n * Ʃ hi где, hi – количество гетерозигот на объем выборки, усредненное по локусам.
Ожидаемая гетерозиготность вычислялась по формуле
Не = 1 – Са, где, Са – ожидаемая гомозиготность, ее определяют через коэффициент гомозиготности по формуле Робертсона:
Ca = Ʃ pi2
где, pi2 – квадраты частот аллелей локусов.
Индекс фиксации (Fis) дает возможность установить связь между индивидуумами отдельной популяции и популяцией в целом. Положительное значение данного показателя означает нехватку гетерозигот в данной популяции, в то время как отрицательное значение индекса указывает на избыток гетерозигот, т.е. количественно отражает отклонение частот встречаемости гетерозиготных генотипов от теоретически ожидаемой по Харди-Вайнбергу:
Fis = 1 – (Ho / He)
где, Ho – наблюдаемая гетерозиготность;
He – ожидаемая гетерозиготность.
Уровень полиморфности это величина, обратная коэффициенту гомозиготности Робертсона и является важным показателем, характеризующий число активно действующих аллелей в популяции:
Ae = 1 / Ca где, Ca – ожидаемая гомозиготность.
Результаты оцениваются по принципу, чем выше степень ожидаемой гомозиготности, тем значительнее снижается генетическое разнообразие в популяции.
Таблица 1 – Локусы для анализа полиморфизма осетровых рыб
|
Локус |
Прямая и обратная последовательность (5´-3´) |
Диапазон фрагментов (bp) |
Структура повтора |
|
AoxD161 |
F:GTTTGАААTGАTTGАGААААTGC R:TGАGАCАGАCАCTCTАGTTАААCАGC |
98-153 |
ATCT |
|
Afug41 |
F:TGАCGCАCАGTАGTАTTАTTTАTG R:TGАTGTTTGCTGАGGCTTTTC |
182-274.5 |
ATCT |
|
LS19 |
F:CATCTTAGCCGTCTGGGTAC R:CAGGTCCCTAATACAATGGC |
124.5-164 |
GTT |
|
Afug135 |
F:GCCААTTCCTGАААTАTАCCАG R:CGАААCCGCTTCАGАCCTT |
184-276 |
ATCT |
|
Afug37 |
F:CAGGGAATCATGAGCACACG R:TGGCGCAGGATTTTGACAC |
144-212 |
ATCT |
|
Spl173 |
F:GGCTTTTGTCTGAAACGTCC R:TGGTGTGTGATTTTGAAGGC |
228-324 |
ATCT |
|
AoxD165 |
F:TTTGАCАGCTCCTААGTGАTАCC R:ААGCCCTАCААCАААTGTCАC |
157-226 |
ATCT |
Результат исследований.
Прогенотипировано 20 особей сибирского осетра, выращиваемых в условиях УЗВ, при котором аллелофонд исследуемой группы по 7 микросателлитным локусам ДНК были получены данные, характеризующие полиморфизм маркеров.
У группы сибирского осетра, выращиваемых в УЗВ, по локусу AoxD161 было выявлено 7 аллелей. Наибольшее часто встречались особи, имеющие аллель 9 – 0,262, а наименьшее количество особей имели аллель 10 (0,033).
В локусе Afug41 у группы сибирского осетра выявлено большее количество аллелей – 11, наиболее высокая частота отмечена для аллеля 17 (0,246), наименьшая частота наблюдалась для аллелей 12 (0,015), 13,3 (0,015) и 18 (0,015). Козлова Н.В. с соавт. проведен анализ генетического разнообразия производителей русского осетра в условиях аквакультуры по микросателлитным ДНК-маркерам, из которого было показано, что локус Afug41 был высокополиморным и имел 17 аллелей [2].
Локус LS19 является одним из наименее полиморфных для исследуемой группы, т.к. содержит 6 аллелей, а высокая частота отмечается для аллелей 10 (0,370) и 11 (0,296). Наименьшей частотой характеризуется аллель 8 (0,019). Georgescu S.E. et al. изучали генетическое разнообразие гибридных осетровых рыб ( A. baerii x A. gueldenstaedtii ), выращиваемых в Румынии. Ими было выявлено, что локус LS19 имел 8 аллелей [6].
Восемь аллелей выявлено в локусе Afug135. В группе сибирского осетра с наибольшей частотой встречаются аллели 10 (0,203) и 14 (0,203). Аллель 16 является одной из самых редких, и ее частота составляет 0,016.
Одним из высокополиморфных для группы сибирского осетра является локус Afug37, т.к. в этом локусе было выявлено 12 аллелей. Наиболее распространены аллели 12 (0,183) и 17 (0,169). Аллели 8, 19 и 20 являются редкими, т.к. их частота в исследуемой группе была одинаковой и составила 0,014. Напротив, B. Welsh et al. было обнаружено, что у озерного осетра Acipenser fulvescens в локусе Afug37 было выявлено всего 5 аллелей, а у зеленого осетра A. medirostris – 7 аллелей [10].
Локус Spl173, также как локусы Afug41 и Afug37, является одним из наиболее полиморфных для исследуемой группы сибирского осетра. В нем выявлено 11 аллелей. С наибольшей частотой встречается аллель 14.3 (0,189), с наименьшей – аллели 12,3, 13 и 15, у которых отмечена одинаковая частота – 0,027.
В локусе AoxD165 выявлено 10 аллелей. В группе наиболее распространенным оказался аллель 13 (0,263), а наименее распространенными – аллели 15.2 и 19.3, их частота была одинаковой и составляла 0,015. При исследовании полиморфизма микросателлитных локусов у сибирского осетра, обитающего в России, в локусе AoxD165 было выявлено 23 аллеля, что является показателем его высокой полиморфности.
Таким образом, анализ полиморфизма ДНК-микросателлитных локусов сибирских осетров, выращиваемых в УЗВ, следует отметить, что всего в 7 локусах идентифицировано 69 аллелей, причем их количество составляет от 6 аллелей в локусе LS19 до 12 – в локусе Afug37.
В ходе анализа генофонда исследуемой группы сибирских осетров, выращиваемых в УЗВ, по 7 STR-локусам были получены данные, характеризующие полиморфизм каждого из маркеров (Таблица 2).
Таблица 2 – Характеристика микросателлитных локусов сибирских осетров, выращиваемых в
УЗВ (n=20)
|
Показатель |
AoxD161 |
Afug41 |
LS19 |
Afug135 |
Afug37 |
Spl173 |
AoxD165 |
Среднее |
|
Диапазон (размер) повторностей |
7-15 |
10-20 |
5-16 |
10-19 |
8-20 |
8,1-16,1 |
11,2-19,3 |
– |
|
Среднее количество аллелей на особь |
3,05 |
3,25 |
2,7 |
3,2 |
3,55 |
1,85 |
3,25 |
2,98 |
|
Наблюдаемая гетерозиготность |
1 |
0,950 |
1 |
0,950 |
1 |
0,550 |
1 |
0,921 |
|
Ожидаемая гетерозиготность |
0,817 |
0,853 |
0,742 |
0,846 |
0,873 |
0,878 |
0,815 |
0,832 |
|
Ожидаемая гомозиготность |
0,183 |
0,147 |
0,258 |
0,154 |
0,127 |
0,122 |
0,185 |
0,168 |
|
Уровень полиморфности |
5,464 |
6,803 |
3,876 |
6,494 |
7,874 |
8,197 |
5,405 |
5,952 |
|
Индекс фиксации |
-0,224 |
-0,114 |
-0,348 |
-0,123 |
-0,145 |
0,374 |
-0,227 |
-0,107 |
—♦—Наблюдаемая гетерозиготность Я Ожидаемая гетерозиготность
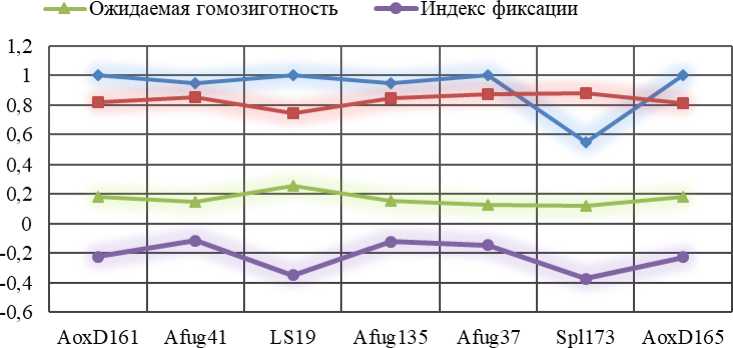
Рисунок 1 – Характеристики полиморфизма микросателлитных STR-локусов исследованной группы сибирского осетра (наблюдаемая и ожидаемая гетерозиготность, ожидаемая гомозиготность, индекс фиксации)
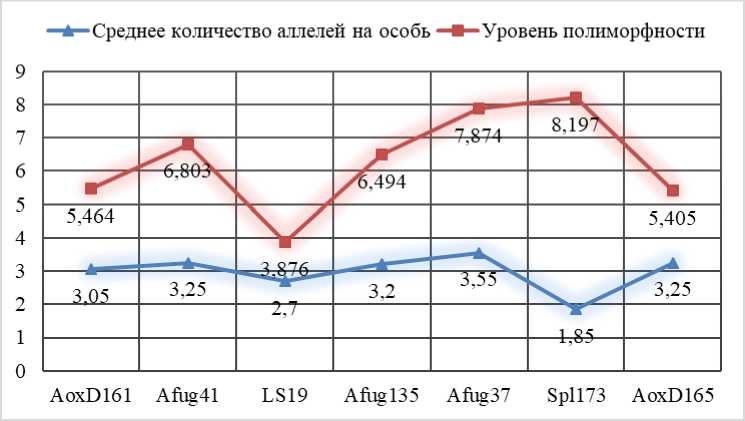
Рисунок 2 – Характеристики полиморфизма микросателлитных STR-локусов исследованной группы сибирских осетров
В вопросах динамики генетического состава популяций важным параметром является гетерозиготность, возникающая при слиянии разнокачественных гамет в гетерозиготу, т.к. она положительно сказывается при адаптации популяций к новым условиям окружающей среды, а также в микроэволюционном процессе.
Наблюдаемая гетерозиготность (Но) является мерой генетической изменчивости в популяции, а для точной оценки изменчивости популяции используют ожидаемую гетерозиготность (Не), рассматривающий уровень аллельного разнообразия, в связи, с чем были оценены как наблюдаемая, так и ожидаемая гетерозиготность по 7 STR-локусам.
По локусу Afug41 у сибирского осетра показатель ожидаемой гетерозиготности составлял 0,853. Согласно данным Barmintseva, A.E. с соавт. ожидаемая гетерозиготность также была высокой и находилась в диапазоне от 0,76 у шипа до 0,99 у сибирского осетра Acipencer baerii [5].
Согласно нашим данным группа сибирского осетра по локусу Afug135 характеризовалась довольно высокой ожидаемой и наблюдаемой гетерозиготностью – 0,846 и 0,950. Напротив, у озерного осетра Acipenser fulvescens наблюдался средний уровень данных показателей, причем наблюдаемая гетерозиготность (0,67) была ниже, чем ожидаемая (0,72).
Наши результаты соотносятся с данными B. Welsh et al., так, в исследуемой ими группе озерного осетра Acipenser fulvescens локус Afug37 также характеризовался высокими ожидаемой и наблюдаемой гетерозиготностью – 0,98 и 1,0. Кроме этого, также наибольшими значениями ожидаемой и наблюдаемой гетерозиготности обладал локус Afug41 – 0,99 и 1,0, соответственно.
Исследуемая группа сибирского осетра по локусу AoxD161 характеризовалась уровнем ожидаемой гетерозиготности равным 0,817. Полученные данные соотносятся с результатами Барминцевой А.Е. с соавт., согласно которым уровень ожидаемой гетерозиготности у российской популяции сибирского осетра Acipencer baerii также был высоким (0,99) [1].
По локусу AoxD165 у исследуемой нами группы сибирского осетра значение ожидаемой гетерозиготности составляло 0,815, это согласуется с данными Барминцевой А.Е. с соавт. по изучению сибирского осетра, обитающего в России – 0,96. В остальных исследуемых ими группах осетровых ожидаемая гетерозиготность была также высокая, за исключением белуги Huso huso – 0,34. Henderson-Arzapalo A., изучая атлантического осетра Acipenser oxyrinchus, выявили, что по локусу AoxD161 значение ожиадемой гетерозиготности составляло 0,853, а наблюдаемой было ниже – 0,733 [7].
В отношении ожидаемого уровня гетерозиготности (Не) характеризовался максимальным показателем – 0,878, а в локусе LS19 – минимальное значение 0,742, а локусы AoxD161, LS19, Afug37 и AoxD165 характеризовались наибольшей наблюдаемой гетерозиготностью (Но). Так, по этим локусам все исследуемые особи были гетерозиготными, соответственно, уровень наблюдаемой гетерозиготности равнялся 1. Наименьшее значение наблюдаемой гетерозиготности было в локусе Spl173 – 0,550 (Рисунок 1).
Анализ данных показателя индекса фиксации (Fis) показал, что только локус Spl173 отличался недостатка гетерозигот 0,374, максимальная в локусе LS19 до -0,348, а во всех остальных случаях наблюдалась различная степень преобладания показателей наблюдаемой гетерозиготности (Но) над ожидаемой (Не).
По данным рисунка 2 видно, что средний показатель уровня полиморфности исследуемых локусов составил 5,952. Наибольший уровень полиморфности наблюдался у локуса Spl173 (8,197), наименьший – у локуса LS19 (3,876).
Заключение. При оценке полиморфизма STR-локусов у исследуемой популяции сибирских осетров, выращиваемых в УЗВ, установлено, что соотношение ожидаемой и наблюдаемой гетерозиготности, а также показателя индекса фиксации, говорит о высоком «запасе» генетического разнообразия по STR-локусам, ввиду избытка гетерозигот.
Исследовательская работа выполнена в рамках грантового финансирования по научным и (или) научно-техническим проектам на 20222024 годы по проекту АР14870980 «Изучение специфических особенностей генетической структуры осетровых рыб и их гибридов, выращиваемых в установках замкнутого водоснабжения», источником финансирования которого является Комитет науки Министерства образования и науки Республики Казахстан.
В статье приведены результаты исследований по оценке полиморфизма по 7 микросателлитным локусам ДНК осетровых рыб, в которых идентифицировано 69 аллелей. Все исследуемые особи были гетерозиготными, соответственно, уровень наблюдаемой гетерозиготности равнялся 1. На основании полученных результатов оценки полиморфизма STR-локусов у сибирских осетров, выращиваемых в УЗВ, установлено, что в исследуемой группе соотношение ожидаемой и наблюдаемой гетерозиготности, а также показателя индекса фиксации (Fis), говорит об избытке гетерозигот в них и высоком «запасе» генетического разнообразия по STR-локусам.
Список литературы STR-генотипирование осетровых рыб, выращиваемых в УЗВ: значение и перспектива применения
- Барминцева, А. Е. Использование микросателлитных локусов для установления видовой принадлежности осетровых (Acipenseridae) и выявления особей гибридного происхождения / А. Е. Барминцева, Н. С. Мюге // Генетика. – 2013. – Т. 49. – № 9. – С. 1093-1105.
- Козлова, Н. Применение молекулярно-генетических исследований в аквакультуре осетровых рыб / Н. Козлова, Н. Базелюк, Д. Файзулина, Е. Стоногина // Вестник Астраханского государственного технического университета. Серия: Рыбное хозяйство. – 2013. – №. 3. – С. 113-117.
- Мюге, Н.С. Полиморфизм контрольного региона митохондриальной ДНК восьми видов осетровых и разработка системы ДНК-идентификации видов / Н. С. Мюге, А. Е. Барминцева, С. М. Расторгуев [и др.] // Генетика. 2008. – Т. 44. – С. 1-7.
- Чебанов, М. С. Формирование генетической коллекции осетровых в южном филиале ФГУП ФСГЦР / М. С. Чебанов // Материалы Первой Всероссийской конференции «Генетика, селекция и воспроизводство рыб», СПб, 2002. – С. 73-80.
- Шалгимбаева, Г.М. Генетическое разнообразие севрюги реки Урал / Г. М. Шалгимбаева, А. Е. Барминцева, Л. Н. Мюге, К. Б. Исбеков, Н. С. Мюге // Труды ВНИРО, 2018. – Т. 171. – С. 95-105.
- Barmintseva, A. E. Natural genetic polymorphism and phylogeography of Siberian sturgeon Acipenser baerii Brandt, 1869 / A. E. Barmintseva, N. S. Myuge, // Russian Journal of Genetics, 2017. – V. 53(3). – Р. 358-368. DOI: 10.1134/S1022795417030024
- Georgescu, S. E. Analysis of the microsatellite variation in the common hybrid between russian sturgeon (Acipenser Gueldenstaedtii) and siberian sturgeon (Acipenser Baerii) from aquaculture / S. E. Georgescu, O. Canareica, A. Dudu, M. Costache // Transylvanian Review of Systematical and Ecological Research. – 2013. – V. 15.2. – Р. 117-124.
- Henderson-Arzapalo, A. Novel microsatellite markers for Atlantic sturgeon (Acipenser oxyrinchus) population delineation and broodstock management / A. Henderson-Arzapalo, T. L. King // Molecular Ecology Notes. – 2002. – V. 2. – P. 437-439.
- Nazari, S. Analysis of the genetic structure of the Persian sturgeon (Acipenser persicus) populations: Comparison of control region sequencing and PCR-RFLP analysis of mitochondrial DNA / S. Nazari, M. Pourkazemi, M. R. Khoshkholgh // Iranian Journal of Fisheries Sciences. – 2020. – V. 19(6). – Р. 3201-3220.
- Nei, M. Accuracy of estimated phylogenetic trees from molecular data. II. Gene frequency data / M. Nei, F. Tajima, Y. Tateno // Journal of Molecular Evolution. – 1983. – Vol. 19. – P. 153-170.
- Welsh, A. B. Identification of microsatellite loci in lake sturgeon. Acipencer fulvescens, and their variability in green sturgeon, A. medirostris / A. B. Welsh, M. Blumberg, B. May // Mol. Ecol. Notes, 2003. – V. 3. – P. 47-55.
