Структура биомассы подроста ели в разновозрастных березняках

Автор: Зарубина Лилия Валерьевна, Пятовская Светлана Александровна
Журнал: Молочнохозяйственный вестник @vestnik-molochnoe
Рубрика: Биологические и сельскохозяйственные науки
Статья в выпуске: 4 (24), 2016 года.
Бесплатный доступ
В березняках черничных типов условий местопроизрастания изучено влияние возрастного состояния лиственного полога на биометрические показатели елового подроста в условиях Сямженского района Вологодской области. Установлено, что с увеличением возраста березы условия для роста и развития хвойного подроста ухудшаются, что сказывается на изменении структуры биомассы, морфометрических показателей и возраста хвои подпологовой ели.
Разновозрастные березовые насаждения, черничные типы условий местопроизрастания, подрост ели, структура биомассы
Короткий адрес: https://sciup.org/14998879
IDR: 14998879 | УДК: 574.36:630*5
Spruce undergrowth biomass structure in birch forests of various ages
In birch forests of bilberry type in vegetation conditions the influence of age statement of leaf-canopy on biometrical parameters of spruce undergrowth under the Syamzhenskiy district of the Vologda region is studied. It is established that with the birch age increase the conditions for growth and development of conifer undergrowth deteriorate, as a result it changes the biomass structure, morphometrical parameters and age of under canopy needles.
Текст научной статьи Структура биомассы подроста ели в разновозрастных березняках
Введение. Зарождение идеи об использовании производительных сил природы в целях восстановления леса при рубках произошло на заре отечественного лесоводства, в это время формируются принципы рациональности и постоянства пользования лесом [1]. О необходимости сохранения подроста и запрещения его вырубки указывалось еще в Правилах главного пользования в лесах СССР (1967 г). Однако, вопреки рекомендациям ученых и действующим правилам, на практике эти требования часто не выполнялись. По данным исследователей, внедрение технологий с сохранением подроста позволяло многим бригадам лесорубов не только систематически выполнять производственные задания, но и повышать производительность труда [2]. В настоящее время сбережение подпологового возобновления регламентируется «Правилами заготовки древесины» от 1 августа 2011 г. и «Правилами лесовосстановления» от 16 июля 2007 г. [3]. Большинство исследователей полагают, что появлению елового подроста, его закреплению в среде обитания, росту и развитию сопутствуют разновозрастные по структуре, низкополнотные насаждения. Еловый подрост с хорошей перспективой занимать лидирующие позиции в фитоценозе отмечается также под пологом лиственных и лиственно-хвойных насаждений [4].
Степень сомкнутости полога обратно пропорциональна наличию крупного подроста, т.е. чем больше сомкнутость полога, тем меньше доля среднего и крупного подроста ели [5]. Давно известен факт, что режим освещенности под пологом древостоев оказывает решающее влияние на состояние подроста ели. С возрастом и ростом особей ели постепенно снижается их теневыносливость и увеличивается светолюбивость. Оптимальная сомкнутость полога для развития жизнеспособного подроста ели – 0,5‒0,6 [6]. Жизнеспособный подрост ели в подавляющем большинстве случаев растет только в окнах полога елового древостоя, поскольку в окнах подрост ели не испытывает недостатка света, а к тому же напряженность корневой конкуренции там значительно ниже, нежели в приствольной части древостоя [7]. Хоть ель и относится к теневыносливым породам, подрост ели в высокоплотных насаждениях все же испытывает большие трудности при низкой освещенности. В результате качественные характеристики подроста в плотных насаждениях заметно хуже по сравнению с подростом, растущим в среднеплотных и низкоплотных насаждениях [8].
Своеобразие светового и теплового режимов в подчинённых ярусах насаждений определённым образом сказывается на формировании фотосинтетического аппарата деревьев и на его работе. Недостаток тепла и света, высокая корневая конкуренция за элементы почвенного питания и воду являются основной причиной слабого роста и функционирования [9].
Цель исследования состояла в изучении биомассы подроста ели при сукцессионном развитии березового насаждения черничных типов условий местопроизрастания.
Объекты и методы исследования. Исследование структуры биомассы подроста ели в березняках черничного типа условий местопроизрастания разной стадии онтогенеза проведены на территории Сямженского территориального отдела – государственного лесничества Вологодской области. Объектом исследований являлся подрост ели (Picea abies Karst. (Pinacea) в березняке черничном (Betuletum myrtillosum) разного возраста (табл. 1).
Таблица 1. Лесоводственно-таксационная характеристика исследуемых частков
|
Древостой |
Подрост |
|||||||
|
состав |
средние |
полнота |
бонитет |
общий запас, м3/ га |
состав |
кол- во, экз./га |
высота, м |
|
|
Н, м |
Д, см |
|||||||
|
29-летнее берёзовое насаждение |
||||||||
|
9Б1ЕедОседС |
14,0 |
10,6 |
0,92 |
3 |
107 |
10Е |
6014 |
1,45 |
|
42-летнее берёзовое насаждение |
||||||||
|
8Б2Е+Ос+С |
20,7 |
17,3 |
0,79 |
2 |
179 |
10Е |
4761 |
1,42 |
|
62-летнее берёзовое насаждение |
||||||||
|
9Б1Е+СедОс |
22,9 |
18,6 |
0,73 |
2 |
200 |
10Е |
4302 |
1,39 |
Закладка пробных площадей велась с учётом требований ОСТ 56-69-83 [10]. Перечет подроста проводился методом пробных площадей (ПП) с учетом требований ГОСТ 16128-70 [11].
Биомассу надземной части деревьев определяли методом модельных деревьев [12]. В конце вегетационного периода (август-сентябрь) на каждой пробной площадке подбирались по три модельных деревца ели средней высоты со средними таксационными показателями. Определялась масса отдельных фракций деревца (ствол, отмершие ветви, живые охвоенные (лапник) и неохвоенные ветви, хвоя разных лет). При разделке деревца учитывалась вся крона. После разделки и взвешивания от каждой фракции брались образцы для определения влажности (в 3-кратной повторности). Разделка стволов проводилась по четвертям (0; 0,25; 50; 75). Обработка полевых материалов осуществлялась общепринятыми в лесоводстве и таксации методами.
Результаты исследований и их обсуждение
В связи с активным вовлечением хвойных лесов в промышленную эксплуатацию без проведения мер содействия естественному возобновлению вырубки возобновляются лиственными породами и в дальнейшем на месте хвойных насаждений формируются леса с преобладанием лиственных пород. По данным учета лесов Вологодской области (на 1.01.2015) доля насаждений с преобладанием мелколиственных пород в лесном фонде области составляет 48,9%, в т.ч. березовые ‒ 37,9%. [13]
Под пологом березняков имеется достаточное количество подроста ели (до 5 тыс. экз. на 1 га и более), которое способно обеспечить в последующем господство в составе формирующихся древостоев при условии создания благоприятных экологических условий [7, 14, 15].
По вопросу роли лиственных пород в жизнедеятельности ели в литературе встречаются разные мнения. Основоположник учения о смене пород Г.Ф. Морозов (1949) [16] указывает, что лиственные породы не только не угнетают ель, но даже способствуют ее росту. И.С. Мелехов (1953) [17] считает, что примесь лиственных к ели улучшает плодородие почвы, активизирует обмен веществ и энергий и, в конечном счете, способствует лучшему росту ели. Наряду с этим А.С. Тихонов (1979, 1990) [18, 19] в своих обзорах приводит многочисленные примеры заглушения ели березой и осиной на бывших вырубках. По его данным даже двухразовое осветление культур ели на вырубках не дает нужного эффекта, поскольку численность бе- резы восстанавливается быстрее ели, и она вновь начинает угнетать ель. Одной из основных причин слабого роста подроста ели в березняках является то, что корни березы обладают большей энергией роста, они более интенсивны по сравнению с хвойными породами. На участках с одинаковым качеством лесорастительных условий корневая система березы гуще, она проникает в почву глубже, чем корневые системы хвойных деревьев, вытесняя их в верхние горизонты [20].
По исследованиям многих авторов [21, 22], недостаток света является основным внешним фактором, лимитирующим нормальный рост подроста в древостоях. Л.А. Иванов (1946) [23] свет и влагу считал главными внешними факторами в жизни древесных растений. Интенсивность света определяет видовой состав, габитус и численный состав растений нижних ярусов леса с умеренным климатом на почвах с достаточным увлажнением [24]. В условиях угнетения под пологом древостоев ель и пихта формируют характерную зонтикообразную форму кроны [25].
Адаптация подроста к экологическим условиям местопроизрастания на морфологическом уровне происходит через изменение целого ряда его биометрических показателей. Еловый подрост в березняках разного возраста имеет различную структуру фитомассы. Наши исследования показали, что у подпологовой ели при возрастном развитии березняков в соответствии со световым режимом изменяется величина главных и боковых побегов, количество и масса на них хвои. При примерно одинаковой высоте моделей у подроста наибольшая масса хвои накапливается в 29-летнем березняке. Крона каждого деревца при высоте 145 см здесь содержит 6,1 г вегетирующей абсолютно сухой хвои, что составляет от общей массы деревца 0,8%. В 62-летнем березняке деревце аналогичной высоты имеет вегетирующей хвои в 5 раза меньше. Это, по нашему мнению, объясняется недостатком фотосинтетической активной радиации, необходимой для образования органического вещества, в процессе фотосинтеза, поэтому более старые органы отмирают (табл. 2).
Таблица 2. Структура биомассы и содержание воды у подроста ели в берёзовых насаждениях черничного типа условий местопроизрастания на разных этапах онтогенеза
|
Структура биомассы |
Объект исследований |
|||||||
|
29-летнее берёзовое насаждение |
42-летнее берёзовое насаждение |
62-летнее берёзовое насаждение |
||||||
|
г \ |
% |
г |
% |
г |
% |
|||
|
Хвоя: |
||||||||
|
1-го года |
0,7 |
0,1 |
0,2 |
0,1 |
0,5 |
0,1 |
||
|
2-го года |
2,4 |
0,3 |
0,3 |
0,1 |
0,3 |
0,1 |
||
|
3-го года |
2,2 |
0,3 |
0,3 |
0,1 |
0,4 |
0,1 |
||
|
прочих лет |
0,8 |
0,1 |
0,3 |
0,1 |
0 |
0 |
||
|
Итого |
6,1 |
0,8 |
1,1 |
0,4 |
1,2 |
0,3 |
||
|
Побеги: |
||||||||
|
1-го года |
13,9 |
1,9 |
4,7 |
0,7 |
2,8 |
0,4 |
||
|
2-го года |
5,9 |
0,8 |
10,1 |
1,6 |
4,7 |
0,8 |
||
|
3-го года |
50,2 |
6,8 |
16,0 |
2,5 |
20,6 |
3,3 |
||
|
прочих лет |
241,9 |
32,7 |
181,3 |
29 |
155,0 |
24,8 |
||
|
Итого |
311,9 |
42,4 |
212,1 |
33,8 |
183,1 |
28,9 |
||
|
Живые ветви |
255,1 |
34,4 |
247 |
39,5 |
237,9 |
38,1 |
||
|
Сучья |
4,7 |
0,6 |
6,3 |
1,0 |
22,4 |
3,6 |
||
|
Древесина |
121,8 |
16,5 |
127,7 |
20,4 |
145,8 |
23,4 |
||
|
Кора |
40,4 |
5,5 |
30,8 |
4,9 |
34,3 |
5,5 |
||
|
Всего |
740 |
100 |
625 |
100 |
624,7 |
100 |
||
|
Высота, м |
1, |
45 |
1,42 |
1,39 |
||||
|
Возраст, лет |
18 |
27 |
42 |
|||||
Масса живых ветвей на деревцах в изучаемых насаждениях также оказалась подверженной значительной изменчивости в соответствии со световыми условиями. Наибольшая их масса нами отмечена в 29-летних насаждениях (255,1 г), наименьшая – в 62-летних березняках (237,9 г). Масса сухих сучьев в расчёте на одно деревце с ухудшением условий увеличивается с 4,7 г до 24,4 г (или на 80%). Отмечая факт уменьшения общей массы хвои на деревце в ухудшающихся условиях существования, следует отметить, что доля хвои в общей массе с увеличением возраста насаждения, то есть с ухудшением освещённости, так же снижается (с 0,8% до 0,3%).
С увеличением возраста подроста ели (с 18 до 42 лет) масса древесины увеличивается (на 7% больше в 62-летнем насаждении по сравнению с 29-летним). Наибольшая масса коры деревца наблюдается в 29-летнем березняке (40,4 г). Долевое участие коры в общей массе существенно не различается при изменении возраста основной породы (4,9‒5,5%).
Общая биомасса деревца ели наибольшая в 29-летнем берёзовом насаждении (740 г), в 62-летнем она составляет на 16% меньше (624,7 г). Это объясняется тем, что под полог поступает больше света и тепла, а так же за счёт менее мощно развитой корневой системы основных деревьев в молодых древостоях подрост ели получает больше количества питательных веществ и воды, это выражается и в более высоком содержании воды в деревцах. Так количество воды в 62-летнем насаждении уменьшилось на 28% по сравнению с 29-летним древостоем.
Долевое участие элементов структуры биомассы в общем объёме деревца при изменении возраста берёзового яруса представлено на рисунке.
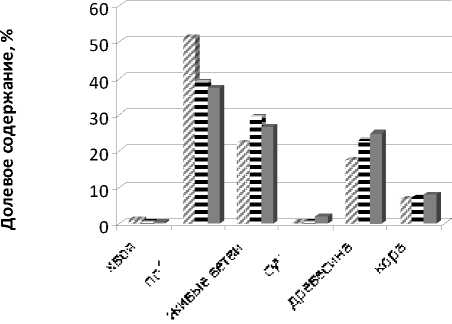
№ 29-летнее насаждение 42-летнее насаждение ■ 62-летнее насаждение
Рисунок. Структура биомассы элементов в общем объёме
Анализ данных показал, что побеги, живые ветви и древесина занимают наибольший объём, в среднем 30%. Доля фотосинтетического аппарата в общей структуре присутствует не более 1%, так как еловому подросту не хватает света и тепла для нормального функционирования. Доля сухих ветвей в общей массе около 5%.
Известно, что длина хвои, как и размер листьев, является важным диагностическим признаком вида, критерием оценки условий местопроизрастания, успешности его роста. Световые условия местообитания формируют ассимиляционный аппарат растений двояким образом: с одной стороны, влияя на ростовые процессы в листьях (Крамер, Козловский, 1963) [26], с другой – путем воздействия на его функциональные особенности [27]. Анализируя полученные данные, можно отметить, что масса 100 штук хвоинок наибольшая ‒ из верхней части кроны средней модели елового подроста, произрастающего в средневозрастном березовом насаждении (от 174 до 224 мг), наименьшая – из нижней части (в среднем на 14%). Условия светового режима оказывают влияние и на длину хвоинок. Наибольшая длина 100 штук хвоинок (126,7 см) наблюдается в верхней части кроны, наименьшая – в нижней части (в среднем на 7%) (табл. 3).
Таблица 3. Средние статистические показатели массы и длины хвои елового подроста в березняке черничного типа условий местопроизрастания
|
Показатель |
Объект исследований |
||||
|
29-летнее насаждение |
42-летнее насаждение |
62-летнее насаждение |
|||
|
М±m |
М±m |
t 1 |
М±m |
t 1 |
|
|
масса 100 штук хвоинок, мг |
208±0,3 |
188±0,2 |
50 |
168±0,3 |
100 |
|
длина 100 штук хвоинок, см |
122,3±2,34 |
87,8±3,31 |
8,75 |
81,2±2,28 |
12,8 |
По данным таблицы видно, что возраст берёзового древостоя оказывает влияние на массу и длину хвои, так как при вероятности безошибочного заключения 95% различия по данным показателям статистически доказаны (tst 0.95= 2,2). Аккумуляция подростом ели атмосферного углерода усиливается с улучшением условий местопроизрастания, ослаблением внутри- и межвидовой конкуренции. Основная часть его используется при формировании нового ассимиляционного аппарата и расходуется на создание общей биомассы деревца [28].
Результаты проведенных исследований позволяют сделать вывод, что в условиях средней тайги с увеличением возраста древостоя (см. табл. 2, 3), с ухудшением освещенности уменьшается общая масса хвои на деревце, доля хвои в общей массе кроны также снижается.
Характерным показателем текущих изменений в физиологическом состоянии хвойного подроста является средняя продолжительность жизни хвои (табл. 4).
Таблица 4. Средняя продолжительность жизни хвои елового подроста на исследуемых объектах
|
Группа высот, м |
Объект исследований |
||
|
29-летнее насаждение |
42-летнее насаждение |
62-летнее насаждение |
|
|
До 0,5 |
4,9 |
3,2 |
2,1 |
|
0,6–1,0 |
6,4 |
3,3 |
2,5 |
|
1,1–1,5 |
7,6 |
3,7 |
2,8 |
|
1,6–2,0 |
9,2 |
3,4 |
4,3 |
|
2,1–2,5 |
7,2 |
3,0 |
3,3 |
|
выше 2,5 |
5,6 |
2,8 |
2,2 |
|
Средняя продолжительность жизни хвои, лет |
6,8 |
3,2 |
2,9 |
|
Средняя протяженность кроны, см |
95,2 |
62,0 |
59,6 |
Наши исследования показали, что охвоённость деревца находится в прямо пропорциональной зависимости от возраста хвои: при снижении возраста сокращается протяженность кроны. Продолжительность жизни хвои подроста ели, произрастающего под пологом 62-летнего березового насаждения почти в 3 раза меньше, чем у естественного возобновления ели в 29-летнем березняке.
Заключение. Таким образом, по результатам проведенных в условиях Вологодской области исследований можно отметить, что при онтогенезе лиственного полога в березняках черничных типов условий местопроизрастания условия для роста и развития подпологовой ели ухудшаются. Влияние березового древостоя на возобновление ели и формирование еловой популяции неоднозначно и в значительной мере определяется плотностью березового насаждения в разные стадии его онтогенеза. Именно режим среды в основном обусловливает морфогенез и рост ели в процессе формирования еловой популяции.
Анализ роста и формирования подпологовой популяции ели свидетельствует, что для сохранения и ускорения роста ели, переформирования лиственно-еловых лесов в елово-лиственные и еловые насаждения необходимо изреживание лиственного яруса путем своевременного проведения выборочных форм рубок.
Список литературы Структура биомассы подроста ели в разновозрастных березняках
- Шерголь, О. Э. Лесоводственные результаты различных рубок, применявшихся в прошлом в Пермской области//Леса Урала и хозяйство в них/О. Э. Шерголь, С. А. Дыренков, Э. О. Кобак. -Свердловск, 1970. -№5. -С. 121-123.
- Некрасов, М. Д. Об экономической эффективности сохранения подроста/М. Д. Некрасов//Лесное хозяйство. -1990. -№9. -С. 18-19.
- Дебков, Н. М. Идеи о сбережении подроста: зарождение, течение и современные тенденции/Н. М Дебков//Устойчивое лесопользование. -2012. -№2. -С. 26-31.
- Писаренко, А. И. Лесовосстановление: монография/А. И. Писаренко. -М.: Лесная промышленность, 1977. -250 с.
- Чертовской, В. Г. Еловые леса европейской части СССР/В. Г. Чертовской. -М.: Лесная промышленность, 1978. -176 с.
- Алексеев, А. В. Световой режим леса/А. В. Алексеев. -Л.: Наука, 1975. -227 с.
- Грязькин, А. В. Возобновительный потенциал таёжных лесов/А. В. Грязькин. -СПб.: СПбГЛТА, 2011. -188с.
- Афанасьев, В. Н. О росте молодняка ели под пологом лиственных пород/В. Н. Афанасьев//Лесной журнал. -1962. -№3. -С. 5-8.
- Карпов, В. Г. Факторы регуляции экосистем еловых лесов/В. Г. Карпов. -Л.: Наука, 1983. -318с.
- ОСТ 56 69-83 Площади пробные лесоустроительные. Методы закладки.
- ГОСТ 16128-70. Площади пробные лесоустроительные. Метод закладки. -М.: Изд-во стандартов, 1971. -23 с.
- Молчанов, А. А. Методика изучения прироста древесных растений/А. А. Молчанов, В. В. Смирнов. -М.: Наука, 1967. -100 с.
- Официальный сайт Департамента лесного комплекса Вологодской области. -Режим досткпа: forestvologda.ru
- Зарубина, Л. В. Эколого-физиологические особенности ели в березняках черничных: монография/Л. В. Зарубина, В. Н. Коновалов. -Архангельск: ИД САФУ, 2014. -378с.
- Зарубина, Л. В. Оценка естественного лесовосстановления в мелколиственных лесах Севера и на вырубках из-под них/Л. В. Зарубина//Вестник Северного (Арктического) федерального ун-та: Сер. Естественные науки. -2015. -№ 2. -С. 35-45.
- Морозов, Г. Ф. Учение о лесе/Г. Ф. Морозов. -Л.: Гослесбумиздат, 1949. -456 с.
- Мелехов, И. С. Пути повышения производительности лесов в таежной зоне/И. С. Мелехов//Некоторые вопросы ведения лесного хозяйства на Севере. -Архангельск: Арханг. кн. изд-во, 1953. -С. 5-18.
- Тихонов, А. С. Лесоводственные основы различных способов рубок для возобновления ели/А. С. Тихонов. -Л.: ЛГУ, 1979. -248 с.
- Тихонов, А. С., Теория и практика рубок леса/А. С. Тихонов, С. С Зябченко. -Петрозаводск: Карелия, 1990. -224 с.
- Мартинович, Б. С. Особенности роста и жизнедеятельности ели и березы в смешанных елово-березовых насаждениях/Б. С. Мартинович, Г. И. Кабашникова, Л. А. Крот//Регулирование роста, развития и питания растений в фитоценозах/под ред. И. Н. Рахтеенко. -Минск: Наука и техника, 1982. -С. 49-78.
- Грязькин, А. В. Влияние факторов внешней среды на структуру и состояние подроста/А. В. Грязькин//Известия Санкт-Петербургской лесотехнической академии. Вып. 8 (166). -СПб.: СПбГЛТА, 2000. -С. 19-25.
- Мигунова, Е. С. Лес и лесные земли (количественная оценка взаимосвязей)/Е. С. Мигунова. -Харьков: Новое слово, 2010. -364 с.
- Иванов, Л. А. Свет и влага в жизни древесных растений/V Тимирязевские чтения/Л. А. Иванов. -М.: Из-во АН СССР, 1946. -46 с.
- Цельникер, Ю. Л. Влияние интенсивности света на параметры структуры кроны ели/Ю. Л. Цельникер//Лесоведение. -1995. -№ 5. -С. 73-78.
- Дружинин Ф.Н. Надземная фитомасса подпологовой ели после комплексных рубок//Известия высших учебных заведений. Лесной журнал. -2012. -№3. -С. 19-24.
- Крамер, П. Д. Физиология древесных растений/П. Д. Крамер, Т. Козловский. -М.: Гослесбумиздат, 1963. -627 с.
- Малкина, И. С. Фотосинтез сосны обыкновенной/И. С. Малкина//Лесоведение. -1981. -№ 4. -С. 83-89.
- Зарубина, Л. В. Динамика формирования пигментного фонда и рост ели в березняке черничном при онтогенезе древостоя/Л. В. Зарубина//Молочнохозяйственный вестник. -2016. -№ 2 (22). -С. 26-36.
