Структура популяций орловских рысаков по генотипам локусов пигментообразования MC1R и Туr и фенотипам масти

Автор: Князев С.П., Гуторова Н.В.
Журнал: Сельскохозяйственная биология @agrobiology
Рубрика: Генетико-селекционное улучшение животных
Статья в выпуске: 2 т.38, 2003 года.
Бесплатный доступ
Проводили сравнительный анализ распределения генотипов по локусам MC1R и Тyr и фенотипам масти на трех конных заводах, разводящих лошадей орловской рысистой породы (Шадринском, Пермском и Алтайском).
Короткий адрес: https://sciup.org/142132874
IDR: 142132874 | УДК: 636.1:636.082
Structure of the Orlov's trotters populations on pigment formation loci MCIR and Tyr, and paint phenotypes
The authors performed the comparative analysis of genotypes distribution on loci MCIR and Tyr, and paint phenotypes on three stud farms for breeding of the Orlov's trotters (Shadrinskii, Permskii and Altaiskii). Differentiation on the frequency of genotypes and phenotypes in horses from different stud farms was revealed. The reliable relationship between phenotypes distribution on paint and genotypes distribution on loci MCIR and Tyr was detected.
Текст научной статьи Структура популяций орловских рысаков по генотипам локусов пигментообразования MC1R и Туr и фенотипам масти
природно-климатических зонах России: Пермский (Средний Урал, n = 53), Шадрин-ский (Курганская область, n = 104) и Алтайский (степные районы Алтайского края, n = 93). Для генетического анализа полиморфизма по локусам MC1R и Tyr использовали образцы ДНК, выделенные из волос гривы 250 племенных особей: все жеребцы-производители, основные штатные конематки трех хозяйств и молодняк Шадринско-го и Алтайского конных заводов, находившийся в заводском тренинге на момент сбора материала.
Выделение и анализ ДНК с использованием полимеразной цепной реакции (ПЦР) проводили в лабораториях Гумбольдтского университета и Свободного университета в Берлине (Германия) в соответствии с программой содружества с Новосибирским государственным аграрным университетом.* Фенотипы лошадей по масти были детально описаны. Статистическую обработку данных проводили с использованием программ Ехсеl-97 и SPSS-2000.
Результаты. Оценка распределения лошадей в трех конных заводах в целом статистически подтвердила значительное преобладание серой и гнедой масти у лошадей орловской рысистой породы (рис. 1). Графическое отображение распределения фенотипов по масти и генотипов по маркерным локусам наглядно демонстрирует преобладание во всех изученных выборках частоты доминантных гомозигот MC1REE и TyrCC над частотой рецессивных гомозигот и соответственно количественное преобладание (среди рысаков несерых мастей) лошадей гнедой и вороной мастей. Такое соотношение генотипов и фенотипов может быть следствием косвенного отбора против мутантных рецессивных гомозигот по локусам MC1R и Туr.
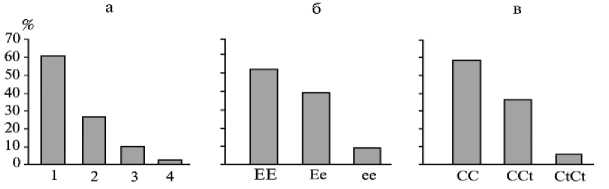
Рис. 1. Распределение частоты (%) фенотипов по масти (а) и генотипов по локусам MC1R (б) и Tyr (в) у лошадей орловской рысистой породы: 1, 2, 3 и 4 — соответственно серая, гнедая, вороная и рыжая масти.
Вероятно, это связано с более высокой жизнеспособностью, лучшими приспособительными качествами, а также б о льшим соответствием критериям отбора в производящий состав лошадей, имеющих в генотипе доминантные аллели обоих локусов. Однако для доказательства этих предположений необходимы дополнительные исследования.
Фенотипическое проявление на уровне масти рецессивной мутации
ТИРОЗИНАЗА
ТИРОЗИНАЗА
Фенилаланин
Тирозин
I
3,4-Дигидроксифенилаланин
Фенилпировиноградная кислота
р-Гидроксифенил----- пировиноградная А / кислота
ТИРОЗИНАЗА
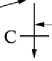
АЛЬБИНИЗМ
МЕЛАНИН
ФЕНИЛКЕТОНУРИЯ
ТИРОЗИНОЗ
2,5-Дигидроксифенил- . пировиноградная кислота
2,5-Дигидроксифенилаланин
Гомогентизиновая кислота
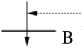
АЛЬКАПТОНУРИЯ
Ацетоуксусная кислота
со2 + Н2О
Рис. 2. Пути метаболизма фенилаланина (цит. по 8): широкие линии означают блокирование отдельных процессов метаболизма, вызывающих фенилкетонурию (А), алькаптонурию (В), альбинизм (С) и тирози-ноз (D).
По данным генетического анализа, частота гомозиготных генотипов MC1REE в популяциях лошадей на Пермском и Алтайском конных заводах была выше, чем на Шадринском заводе (Р < 0,001). Частота гетерозигот по обоим локусам (MC1R и Tyr) в двух первых популяциях имела промежуточные значения при сравнениях с доминантными и рецессивными гомозиготами, а в последнем случае частота гетерозиготных генотипов по локусу MC1R оказалась несколько выше, чем гомозиготных (табл. 1).
1. Частота фенотипов по масти и генотипов по локусам MC1R и Ty r в различных популяциях рысаков орловской породы (%)
|
Конный завод |
Масть ( n = 247) |
Локус MC1R ( n = 202) |
Локус Tyr ( n = 238) |
||||||
|
серая |
гнедая вороная |
рыжая |
ЕЕ |
Ее |
ее |
СС |
ССt |
СtСt |
|
|
Шадринский |
68,0 |
23,3 6,8 |
1,9 |
40,4 |
47,9 |
11,7 |
61,8 |
31,4 |
6,8 |
|
Пермский |
48,1 |
35,2 9,3 |
7,4 |
57,9 |
31,9 |
10,5 |
59,6 |
32,7 |
7,7 |
|
Алтайский |
57,8 |
27,8 14,4 |
0 |
62,5 |
31,9 |
5,6 |
55,9 |
41,9 |
2,2 |
При этом обращает на себя внимание низкая частота встречаемости фенотипа «рыжая масть» и рецессивного гомозиготного генотипа MC1Rее. Распределение генотипов по локусу MC1R не отклонялось от выявленной ранее детерминации мас- тей по локусу Extension, согласно которой гнедые и вороные лошади могут иметь гомо- или гетерозиготный генотип по доминантному аллелю MC1RЕ (без учета лошадей серой масти, так как эпистатичный ген Gray, обусловливающий эту масть, по-
2. Число гомо- и гетерозиготных генотипов по локусам MC1R и Tyr у рысаков орловской породы различной масти
давляет экспрессию других локусов окраски) (3, 10, 12) (табл. 2). Во всех трех выборках лошадей орловской рысистой породы обнаружено всего 6 особей рыжей масти, 19 — гомозигот MClRee, включая 14 серой и 5 рыжей масти.
Рыжая масть лошадей, золотистые окрасы собак (лабрадоры, ретриверы и др.), желтая окраска мышей проявляются у е-аллелю локуса Extension, синонимом кото- рого служит ген MC1R (3, 4, 6). Кроме того, у людей с рыжими волосами и светлой кожей обнаружены мутации в этом локусе (11). Поэтому вполне вероятно, что ген MC1R детерминирует у лошадей рыжую окраску.
В большинстве конских пород могут встречаться как рыжая, так и вороная масти. Однако иногда вследствие отбора, направленного против доминантного аллеля, происходит элиминация последнего (породы, для которых типична только рыжая масть) или, наоборот, при отборе по доминантному аллелю исчезает рецессивная мутация рыжей окраски. Так, у лошадей фризской, кливлендской гнедой и некоторых других пород произошла почти полная элиминация рецессивного е-аллеля, в то время как у лошадей других пород (например хафлингер, шварцвальдская, донская) практически отсутствуют доминантные варианты аллелей Е-локуса (13).
Нами выявлена дифференциация по частоте аллелей локусов MC1R и Tyr и по распределению фенотипов масти лошадей орловской рысистой породы различных популяций (табл. 3). Частота генотипов, имеющих доминантные аллели обоих локусов, незначительно отличалась от таковой по всей исследованной выборке лошадей производящего состава Пермского конного завода. Частота рецессивных гомозигот MC1Rее и ТуrCtCt у лошадей Шадринского конного завода соответствовала средним показателям по всей выборке.
3. Частота фенотипов по масти и генотипов по локусам MC1R и Ty r у лошадей орловской рысистой породы в производящем составе различных конных заводов (%)
|
Масть, генотип |
Конный завод |
Среднее |
||
|
Шадринский |
^ Пермский 1 |
Алтайский |
||
|
Масть: |
||||
|
серая |
65,0 |
49,0 |
56,4 |
57,1 |
|
рыжая |
0 |
5,9 |
0 |
2,3 |
|
вороная |
6,7 |
9,8 |
16,4 |
10,7 |
|
гнедая |
28,3 |
35,3 |
27,2 |
29,9 |
|
Генотип: |
||||
|
ЕЕ |
50,0 |
57,9 |
66,0 |
58,1 |
|
Ее |
42,3 |
31,6 |
30,0 |
35,1 |
|
ее |
7,7 |
10,5 |
4,0 |
6,8 |
|
СС |
66,1 |
58,5 |
63,3 |
60,3 |
|
ССt |
28,8 |
34,0 |
35,0 |
34,5 |
|
СtСt |
5,1 |
7,5 |
1,7 |
5,2 |
Обращает на себя внимание отсутствие лошадей рыжей масти среди производителей и маток Шадринского и Алтайского конных заводов. Наибольшее число лошадей несерой масти (а среди них, гнедой масти) содержалось на Пермском конном заводе.
Все конные заводы обмениваются жеребцами-производителями в зависимости от статуса: хозяйства, имеющие более высокий экономический и племенной статус, получают право чаще и дольше использовать ценных производителей, как, например, Пермский конный завод. Маточный состав всех конных заводов комплектуется на протяжении многих лет кобылами собственного разведения (за редким исключением), поэтому их влияние на генотипическую структуру популяции завода по сравнению с жеребцами-производителями относительно невелико. Производящий состав Алтайского и Шадринского конных заводов пополняется в основном жеребцами из этих же хозяйств; на Шадринский завод в последнее время часто поступают производители из Алтайского конного завода. Этим объясняется некоторая схожесть между генотипической (по локусам MC1R и Tyr ) и фенотипической (по масти) характеристикой производящего состава обоих хозяйств.
При анализе распределения генотипов по локусу Tyr и фенотипов по масти нами не выявлено детерминации окраски шерсти аллелями этого локуса, так как не вполне ясно фенотипическое проявление мутации Сt по сравнению с нормальным аллелем С. Возможно, эта мутация не оказывает влияния на окраску шерсти лошадей. Напротив, локус MC1R ( аллели Е и е ) детерминирует масть лошадей, так как дифференциация генотипической структуры по этому локусу соответствует распределению мастей в изученных выборках.
Дифференциация по частоте встречаемости генотипов, возможно, свидетельствует о том, что наряду с общими тенденциями в породе происходят локальные изменения генотипической структуры в пределах заводских типов. Фенотипически это проявляется в виде образования внутри крупнейших племенных репродукторов так называемых заводских типов — групп животных, обладающих только им присущим комплексом признаков (соответственно общему направлению отбора), в том числе приспособленностью к конкретным эколого-географическим условиям, и наряду с этим соответствующих стандарту породы по общему комплексу признаков отбора (экстерьер, промеры, работоспособность, плодовитость и т.д.). Соотношение лошадей по масти в каждом из трех заводов отражает существующую дифференциацию, которая возникла, видимо, как результат косвенного отбора, когда при селекции по признакам работоспособности, типа экстерьера и плодовитости элиминируются определенные фенотипы по масти, возможно, генетически сопряженные с приспособительными качествами.
Итак, комплексы признаков (селекционируемых и «нейтральных») обусловлены для каждого заводского типа лошадей орловской рысистой породы складывающейся в пределах микропопуляций структурой генофонда, которая, в свою очередь, может представлять собой не простое суммирование генов всех членов популяции, а совокупность взаимосвязанных генетических элементов. Своеобразие популяции зависит от модального по соответствующим локусам «комплексного генотипа» производителей, которые вносят наибольший вклад в формирование генофонда при селекции.
Таким образом, проведенное исследование выявило дифференциацию обследованных выборок орловских рысаков по фенотипам (масти) и генотипам молекулярно-генетических маркеров ( MC1R и Tyr ), участвующих в детерминации пигмен-тообразования. Изменчивость популяций по этим параметрам характеризуется однонаправленностью и комплементарностью. Обнаружена достоверная зависимость распределения лошадей по фенотипам масти от распределения их генотипов по локусам MC1R и Tyr . Это позволяет использовать апробированный метод генетического анализа для мониторинга динамики изменений генетической структуры «закрытой» орловской породы и ее отдельных популяций на заводах, где под влиянием разных факторов формируются и дифференцируются различные фено- и генофонды.
Л И Т Е Р А Т У Р А
-
1. К р а в ч е н к о Н.А. Разведение сельскохозяйственных животных. М., 1973.
-
2. A n d e r s s o n L., S a n d b e r g K. A linkage group composed of three coat color genes and three serum
protein loci in horses. J. Heredity, 1982, 73: 91-94.
-
3. R о b b i n s L.S., N a d e a u J.H., J о n s о n K.R. e.a. Pigmentation phenotypes of variant extension locus alleles result from point mutations that alter MSH receptor function. Cell, 1993, 72: 827-834.
-
4. M a r k l u n d L., M o l l e r Jo., S a n d b e r g K. e.a. A missense mutation in the gene for melanocytestimulating hormone receptor ( MC1R ) is associated with the chestnut coat color in horses. Mammalian Genome, 1996, 7: 895-899.
-
5. П р а с о л о в а Л.А., Т р у т Л.Н., В с е в о л о д о в Э.Б. и др. Морфология пигментации волос у диких рыжих, серебристо-черных лисиц и их гибридов. Генетика, 2002, 38, 4: 463-467.
-
6. J a c k s o n I.J. Molecular and developmental genetics of mouse coat color. Ann. Rev. Gentechnology, 1994, 28: 189-217.
-
7. R e i s s m a n n M., W a g n e r H.J., H a r d g e T. e.a. Sequencing and mutation analysis in exon 1 of horse tyrosinase gene. 27th Intern. Conf. on Animal Genetics. Univ. of Minnesota, 2000.
-
8. Х а т т Ф. Генетика животных. М., 1969.
-
9. В у о n g S., Н a q А.К., W a k u 1 с h i k M. e.a. Isolation, chromosomal mapping and expression of the
mouse tyrosinase gene. J. of Investigative Dermatology, 1989, 5: 93.
-
10. К н я з е в С.П., Г у т о р о в а Н.В., Р а й с с м а н н М. и др. Структура популяции орловского рысака по локусам рецептора меланоцитстимулирующего гормона ( MC1R ) и тирозиназы ( Туr ) . Докл. РАСХН, 2002, 4: 39-42.
-
11. V а 1 v е г d e P., H e a l y E., J a c k s o n I. e.a. Variants of the melanocyte-stimulating hormone recep
tor gene are associated with red hair and fair skin in humans. Nature Genetics, 1995, 11: 328-330.
-
12. К н я з е в С.П., Р а й с с м а н н М., В а г н е р X. и др. Анализ генетической структуры племенного
ядра Российской популяции чистокровных верховых лошадей по локусу Extension с использованием молекулярной ДНК-диагностики. Генетика, 2001, 37, 10: 1383-1387.
-
13. В о w l i n g Ann T. Horse genetics. Wallingford, 1996.
