Таксономическое разнообразие и ареалогия орибатид (Oribatei) европейского севера России
 европейского севера России")
Автор: Мелехина Е.Н.
Журнал: Известия Коми научного центра УрО РАН @izvestia-komisc
Рубрика: Биологические науки
Статья в выпуске: 2 (6), 2011 года.
Бесплатный доступ
Обобщены результаты исследований автора и имеющиеся литературные дан- ные о таксономическом разнообразии орибатид европейского Севера России. Для региона известно 354 вида, 132 рода, 67 семейств орибатид. Отмечена относительная бедность таксономического состава по сравнению с более низ- кими широтами и западными фаунами. По типу долготного распространения преобладают голарктические и палеарктические виды. В тундровой зоне по- вышается доля голарктических видов и снижается - палеарктических и ев- ропейских, по сравнению с таежной зоной. Характерна высокая степень сходства с сибирской фауной. Выделены пять типов широтного распростране- ния видов: арктический, аркто-бореальный, бореальный, температный, поли- зональный. Фауна орибатид равнинной и горной субарктической тундр сход- на с таежной.
Таксономическое разнообразие, биогеография, ареалогия, орибатиды, европейский север России
Короткий адрес: https://sciup.org/14992450
IDR: 14992450 | УДК: 595.423:591.1(470.1)
Taxonomic diversity and areology of oribatid mites (Oribatei) of the European North of Russia
354 species, 132 genera and 67 families of the oribatid mites are known in the European North of Russia. Reduced taxonomic diversity of oribatids is noted in comparison with lower latitudes and western faunas. Oribatids species with the wide areals predominate (holarctic and palaearctic). The characteristic feature is high degree of similarity to the Siberian fauna. According to latitudinal distribution, arctic, arctic-boreal, boreal, temperate and polizonal species are present. There is high degree of similarity between species composition of oribatids in plain and mountain areas of subarctic tundra and boreal zone.
Текст научной статьи Таксономическое разнообразие и ареалогия орибатид (Oribatei) европейского севера России
Для ряда групп наземных беспозвоночных европейской части России (чешуекрылых, двукрылых, жесткокрылых и других) разработаны типологии ареалов, подробно изучена зоогеографическая структура фауны. В отношении типично почвенных животных в этой области сделаны лишь первые шаги. Д.А. Криволуцкий (2004) указывал, что почвенная микрофауна является новой для биогеографии группой животных. Обозначая задачи современной биогеографии почвы, он писал: «К настоящему времени собрано достаточно данных для перехода от локального изучения фауны …, структуры экосистем почвы к обобщению и анализу их географического распространения, ареалогии, районированию» [1].
Орибатиды, или панцирные клещи, являются удобной модельной группой биогеографических исследований. Это мелкие членистоногие, обитающие в почве и других субстратах: мхах, лишайниках, разлагающейся древесине. Они составляют подотряд Oribatida отряда акариформных клещей (Acari-formes) [2]. Высоко таксономическое разнообразие группы. Известно около 9 тыс. видов, более 1200 родов, 169 семейств орибатид в современной мировой фауне [3]. Орибатиды распространены во всех природно-климатических зонах, всегда многочисленны (до 150 тыс. экз./м2 – в лесных биоценозах; 5-20 тыс. экз./ м2–в почвах субарктических тундр).
За последние пять десятилетий накоплен обширный материал о таксономическом разнообразии орибатид таежной и тундровой зон европейской части России, арктических островов, западного макросклона Уральского хребта. Целью настоящей работы было обобщение имеющихся данных, выявление закономерностей изменения состава и структуры фауны орибатид в широтно-зональном аспекте, особенностей географического распространения видов.
Материал
Исследования охватывают аркто-бореальную зону европейской части России (рисунок). В таежной зоне локальные фауны орибатид изучались в Мурманской области: на беломорских островах Кандалакшского заповедника [4], в Ленинградской [5, 6], Архангельской [7], Пермской [8], Кировской [9] областях, Карелии [10, 11], Республике Коми [12 – 19].
Для сравнения использованы данные по о. Западный Шпицберген (норвежский сектор Арктики), где достаточно полно изучена фауна почвенных мик-роартропод [26, 27].
Автором проведено изучение фаунистического разнообразия орибатид на европейском Северо-Востоке в период с 1988 по 2010 гг. Значительно пополнен фаунистический список панцирных клещей таежной зоны [17, 18], получены первые дан-
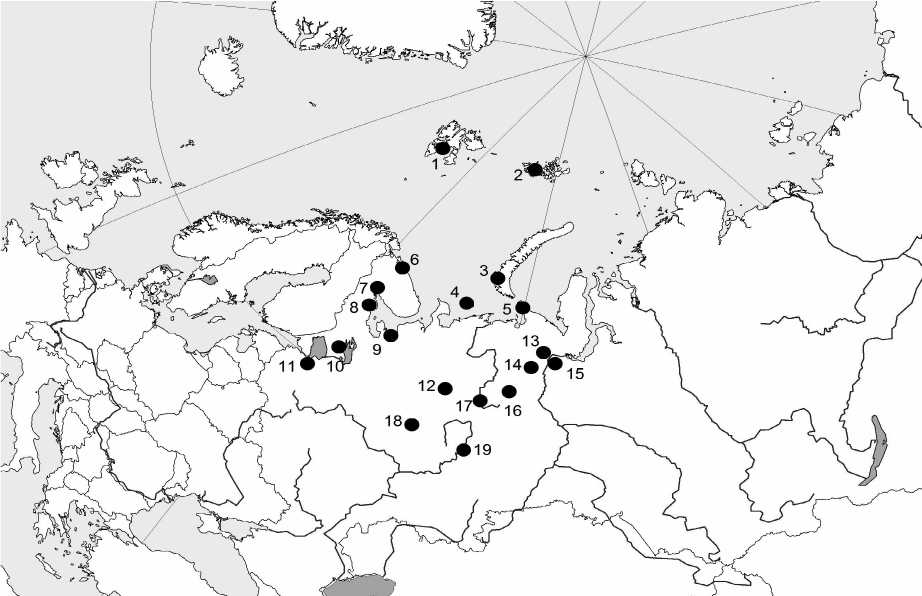
Рис. Локальные фауны орибатид европейского Севера.
Арктические острова и архипелаги:
1 – о. Западный Шпицберген, 2 – арх. Земля Франца-Иосифа, 3 – арх. Новая Земля, 4 – о. Вайгач, 5 – о. Колгуев;
Тундровая зона:
6 – пос. Дальние Зеленцы, 13 – г. Воркута;
Таежная зона:
7 – беломорские острова Кандалакшского заповедника, 8 – Беломорская биологическая станция МГУ, 9 – Архангельская обл., 10 – Карелия, 11 – Ленинградская обл., 12 – Республика Коми, 18 – Кировская обл., 19 – Пермская обл.;
Полярный Урал:
14 – окрестности оз. Пага-ты, 15 – окрестности пос. Полярный;
Северный Урал:
16 – пос. Якша, 17 – г. Яны-Пупу-Ньер.
В равнинной субарктической тундре исследования проведены вблизи пос. Дальние Зеленцы (Мурманская область) [20] и в окрестностях г. Воркуты [21]. Получены сведения о фауне орибатид Полярного Урала: в районе оз. Пага-ты [22] и неподалеку от пос. Полярный – в Лабытнангском районе Ямало-Ненецкого национального округа [23]. На Северном Урале сбор данных проведен в Печоро-Илычском заповеднике: в предгорных [12] и горных [17] сообществах.
Фауну орибатид изучали на островах (Вайгач, Колгуев) и архипелагах (Земля Франца-Иосифа, Новая Земля) российского сектора Арктики [24, 25].
ные о таксономическом составе орибатид тундровой зоны региона [21], западного макросклона Северного и Полярного Урала [16, 22]. При сборе полевого материала использованы стандартные методы [2]: в каждом конкретном местообитании отбирали не менее 10 проб площадью 100 см2 каждая. При этом применяли случайную выборку. Обследовали спектр зональных растительных сообществ, а также интразональных биогеоценозов.
Система орибатид и типы долготного распространения приводятся в работе по L. Subias [3]. При определении типов широтного распространения мы придерживались классификации К.Б. Городкова [28].
Результаты и обсуждение
Для европейского Севера России, по опубликованным данным, известно 354 вида, 132 рода, 67 семейств орибатид. Наибольшее число родов свойственно семействам Oppiidae, Ceratozetidae, Damaeidae, Brachichthoniidae (таблица). Более 56% семейств представлено одним родом. Около 15% семейств включает десять и более видов, около 24% – от пяти до девяти видов, 16 % – три-четыре, более 44 % – один-два вида.
Для фауны орибатид европейского Севера России свойственна относительная бедность таксономического состава. Для сравнения, в средней полосе известно около 700 видов орибатид, в фауне России – более 1300 [2]. Отмечено обеднение фауны по сравнению с западными регионами. На европейском Севере России не найдено по меньшей мере 150 видов орибатид, представленных в Скандинавских странах. В фауне региона не отмечено пять семейств, которые зарегистрированы на территории Швеции и Финляндии [29, 30], в то же время на территории России они присутствуют только в южных широтах: Atopochthoniidae, Cosmo-chthoniidae, Parhypochthoniidae, Mochlozetidae, Hap-lochthoniidae. Отсутствуют южные семейства, к примеру, такие, как Mesoplohporidae, Licnodamaeidae. Многие южные семейства (Zetorchestidae, Gymno-damaeidae, Xenillidae, Liodidae и др.) представлены единичными видами.
В таежной зоне обнаружено 320 видов ори-батид из 132 родов и 65 семейств. Наибольшей видовой насыщенностью характеризуются семейства Ceratozetidae, Suctobelbidae, Oppiidae, Camisii-dae, Phthiracaridae, Carabodidae, Damaeidae, Oriba-tulidae. Ряд семейств всегда присутствует в региональных списках и дополняет перечень семейств, типичных для таежной зоны: Nothridae, Namher-manniidae, Euphthiracaridae, Eremaeidae, Scheloriba-tidae, Chamobatidae, Achipteriidae, Galumnidae, Ori-botritiidae.
По типу долготного распространения выделены следующие группы видов: 1 – голарктические (34.3%), 2 – встречающиеся в Голарктике и некоторых регионах за ее пределами (7.2%), 3 – палеарктические (33.5%), 4 – встречающиеся в Палеарктике и некоторых регионах за ее пределами (4%), 5 – космополиты и полукосмополиты (9.5%), 6 – европейские (11.5%).
По типу широтного распределения в таежной зоне представлены четыре группы видов: полизо-нальные (62.7%), температные (31.0%), бореальные (3.9%) и аркто-бореальные (2.4%). Первые три группы названы по К.Б. Городкову [28]. Группа арк-то-бореальных включает виды, область распространения которых охватывает арктические острова, тундровую и таежную зоны. Группа бореальных видов, отмеченных только в таежной зоне – весьма малочисленна. В нее вошли, наряду с другими, шесть видов рода Ameronothrus: A. maculatus (Michael, 1882); A. bilineatus (Michael, 1888); A. nidicola (Sitnikova, 1975); A. nigrofemoratus (L. Koch, 1879); A. dubinini (Sitnikova, 1975); A. oblongus (Sitnikova, 1975).
Первые пять из них, а также аркто-бореальный A. lineatus (Thorell, 1871) найдены на беломорских островах Кандалакшского заповедника (северная подзона тайги) [4]; вид A. оblongus зарегистрирован в средней тайге Республики Коми. К этой же группе мы отнесли Spatiodamaeus boreus (Bulanova-Zachvatkina, 1957) , распространение которого, с учетом высотной составляющей ареала, можно охарактеризовать как борео-монтанное, поскольку помимо таежной зоны Севера вид встречается на Полярном Урале [23] и Кавказе [31]. Из бореальных видов в высоких широтах Неарктики отмечаются A. nigrofemoratus, Hermannia (Heterohermannia) subglabra (Berlese, 1910) .
В южной тундре Кольского полуострова (пос. Дальние Зеленцы) найдено 54 вида орибатид 34 родов и 19 семейств [4]. Отличительной чертой данной локальной фауны является высокое видовое богатство семейств Oppiidae, Suctobelbidae, Brachichthoniidae – мелких тонкопанцирных форм. В подзоне южной кустарниковой тундры европейского Северо-Востока обнаружено 47 видов 35 родов и 24 семейств орибатид [21]. В обследованных горнотундровых сообществах Полярного Урала, в окрестностях оз. Пага-ты, присутствовало 38 видов панцирных клещей 30 родов и 21 семейства [22]. В окрестностях пос. Полярный найдено 82 вида 34 семейств [23]. Фаунистический список орибатид западного макросклона Полярного Урала насчитывает 101 вид 34 семейств. В горно-тундровом поясе Северного Урала (г. Яны-Пупу-Ньер) обнаружено 25 видов орибатид 22 родов и 14 семейств. В целом таксономическое разнообразие панцирных клещей горно-тундрового пояса было ниже по сравнению с горно-лесным, где зарегистрирован 41 вид 34 родов и 20 семейств [17]. Всего для субарктической тундры европейской части России известно к настоящему времени около 100 видов орибатид 54 родов и 29 семейств. По сравнению с таежной зоной наблюдается значительное обеднение фауны панцирных клещей как на уровне видов, так и более высоких таксонов – родов и семейств. Ранее отмечалась четкая тенденция обеднения фауны орибатид при продвижении к северу [25].
Наибольшее видовое богатство характерно для семейств Camisiidae, Ceratozetidae, Oppiidae, Suctobelbidae, Brachichthoniidae. Эти же семейства, наряду с другими, отличаются значительной видовой насыщенностью в таежной зоне. В тундровой зоне высока доля широко распространенных видов, по сравнению с таежной. К голарктическим относится 48.9% видов, еще 7.4% – виды, которые распространены в Голарктике и встречаются за ее пределами, их суммарная доля – 56.3%. Палеарктические виды составляют 26.5% списка. Доля европейских видов (немногим более 4%) почти втрое меньше, чем в таежной зоне. Значительно число космополитов и полукосмополитов (около 11%).
Многие виды распространены циркумполярно (Danks, 1981) [32]. Среди них Liochthonius (Liochthonius) lapponicus (Trägardh, 1910), Liochtho-nius (L.) brevis (Michael, 1888) (= Brachychthonius perpusillus Berlese, 1910), Liochthonius (L.) sellnicki
Структура фауны орибатид европейского Севера России
|
№ |
Семейство |
Число |
|
|
видов —1 родов |
|||
|
1 |
Oppiidae Grandjean, 1951 |
22 |
12 |
|
2 |
Suctobelbidae Grandjean, 1954 |
22 |
2 |
|
3 |
Ceratozetidae Jacot, 1925 |
21 |
7 |
|
4 |
Damaeidae Berlese, 1896 |
19 |
7 |
|
5 |
Brachichthoniidae Balogh, 1943 |
19 |
6 |
|
6 |
Camisiidae Oudemans, 1900 |
17 |
2 |
|
7 |
Phthiracaridae Perty, 1841 |
16 |
3 |
|
8 |
Liacaridae Sellnick, 1928 |
11 |
3 |
|
9 |
Galumnidae Jacot, 1925 |
10 |
4 |
|
10 |
Carabodidae Koch, 1837 |
10 |
1 |
|
11 |
Punctoribatidae Thor, 1937 |
10 |
3 |
|
12 |
Achipteriidae Thor, 1929 |
9 |
3 |
|
13 |
Oribatulidae Thor, 1929 |
9 |
2 |
|
14 |
Malaconothridae Berlese, 1916 |
9 |
2 |
|
15 |
Phenopelopidae Petrunkevitch, 1955 |
9 |
2 |
|
16 |
Eremaeidae Oudemans, 1900 |
7 |
4 |
|
17 |
Nothridae Berlese, 1896 |
7 |
1 |
|
18 |
Humerobatidae Grandjean, 1970 |
6 |
2 |
|
19 |
Chamobatidae Thor, 1938 |
6 |
1 |
|
20 |
Euphthiracaridae Jacot, 1930 |
5 |
3 |
|
21 |
Oribotritiidae Grandjean, 1954 |
5 |
2 |
|
22 |
Autognetidae Grandjean, 1960 |
5 |
2 |
|
23 |
Nanhermanniidae Sellnick, 1928 |
5 |
1 |
|
24 |
Tectocepheidae Grandjean, 1954 |
5 |
1 |
|
25 |
Ameronothridae Willmann, 1931 |
5 |
1 |
|
26 |
Oribatellidae Jacot, 1925 |
5 |
1 |
|
27 |
Astegistidae Balogh, 1961 |
4 |
3 |
|
28 |
Ceratoppiidae Kunst, 1971 |
4 |
2 |
|
29 |
Scutoverticidae Grandjean, 1954 |
4 |
2 |
|
30 |
Haplozetidae Grandjean, 1936 |
4 |
2 |
|
31 |
Protoribatidae J. et P.Balogh, 1984 |
4 |
1 |
|
32 |
Oribellidae Kunst, 1971 |
3 |
3 |
|
33 |
Tegoribatidae Grandjean, 1954 |
3 |
3 |
|
34 |
Trhypochthoniidae Willmann, 1928 |
3 |
2 |
|
35 |
Hermanniidae Sellnick, 1928 |
3 |
1 |
|
36 |
Cepheidae Berlese, 1896 |
3 |
1 |
|
№ |
Семейство |
Число |
|
|
видов |
родов |
||
|
37 |
Scheloribatidae Grandjean, 1953 |
3 |
1 |
|
38 |
Passalozetidae Grandjean, 1954 |
2 |
2 |
|
39 |
Cymbaeremaeidae Sellnick, 1928 |
2 |
2 |
|
40 |
Parakalummidae Grandjean, 1936 |
2 |
2 |
|
41 |
Hypochthoniidae Berlese, 1910 |
2 |
1 |
|
42 |
Steganacaridae Niedbala, 1986 |
2 |
1 |
|
43 |
Hermanniellidae Grandjean, 1934 |
2 |
1 |
|
44 |
Palaeacaridae Grandjean, 1934 |
2 |
1 |
|
45 |
Thyrisomidae Grandjean, 1954 |
2 |
1 |
|
46 |
Gustaviidae Oudemans, 1900 |
2 |
1 |
|
47 |
Limnozetidae Thor, 1937 |
2 |
1 |
|
48 |
Hydrozetidae Grandjean, 1954 |
2 |
1 |
|
49 |
Liebstadiidae J. et P.Balogh, 1984 |
2 |
1 |
|
50 |
Hemileiidae J. et P.Balogh, 1984 |
1 |
1 |
|
51 |
Quadroppiidae Balogh, 1983 |
1 |
1 |
|
52 |
Xenillidae Woolley et Higgins, 1966 |
1 |
1 |
|
53 |
Liodidae Grandjean, 1954 |
1 |
1 |
|
54 |
Pheroliodidae Paschoal, 1987 |
1 |
1 |
|
55 |
Micreremidae Grandjean, 1954 |
1 |
1 |
|
56 |
Zetorchestidae Michael, 1898 |
1 |
1 |
|
57 |
Caleremaeidae Grandjean, 1965 |
1 |
1 |
|
58 |
Ctenobelbidae Grandjean, 1965 |
1 |
1 |
|
59 |
Damaeolidae Grandjean, 1965 |
1 |
1 |
|
60 |
Eniochthoniidae Grandjean, 1947 |
1 |
1 |
|
61 |
Eremobelbidae Balogh, 1961 |
1 |
1 |
|
62 |
Eulohmanniidae Grandjean, 1931 |
1 |
1 |
|
63 |
Gymnodamaeidae Grandjean, 1954 |
1 |
1 |
|
64 |
Licneremaeidae Grandjean, 1931 |
1 |
1 |
|
65 |
Tenuialidae Jacot, 1929 |
1 |
1 |
|
66 |
Euzetidae Grandjean, 1953 |
1 |
1 |
|
67 |
Heterozetidae Kunst, 1971 |
1 |
1 |
|
Всего |
354 |
132 |
|
(Thor, 1930), Nothrus borussicus Sellnick, 1928 , Noth-rus pratensis (Sellnick, 1928), Camisia (Camisia) hor-rida (Hermann, 1804) , Camisia (Ensicamisia) lapponica (Trägardh, 1910), Heminothrus (Heminothrus) long-isetosus Willmann, 1925, Heminothrus (Platynothrus) punctatus (L. Koch, 1879), Ceratoppia bipilis (Hermann, 1804) , Ceratoppia quadridentata (Haller, 1882),
Carabodes (Carabodes) labyrinthicus (Michael, 1879), Moritzoppia (Moritzoppiella) neerlandica (Oudemans, 1900 ), Suctobelbella (Suctobelbella) acutidens (Forss-lund, 1941), Liebstadia similis (Michael, 1888), Oriba-tula (Oribatula) tibialis (Nicolet, 1855), Peloribates ( Peloribates) pilosus (Hammer, 1952) , Melanozetes mollicomus (Koch, 1839) , Eupelops plicatus (Koch,
1844) (= Pelops auritus Koch, 1839) , Campachipteria nivalis (Hammer, 1952) , Tegoribates latirostris (Koch, 1844), Scutozetes lanceolatus (Hammer, 1952) , Neoribates (Neoribates) aurantiacus (Oudemans, 1914) . Этот перечень дополняют космополиты He-minothrus (Platynothrus) peltifer (Koch, 1839), Tecto-cepheus velatus (Michael, 1880 ), Oppiella (Oppiella) nova (Oudemans, 1902), Quadroppia (Quadroppia) quadricarinata (Michael, 1885), Ceratozetes (Cera-tozetes) gracilis (Michael, 1884).
Ареалы большинства видов охватывают более низкие широты и относятся к температному (17.5%) и полизональному (75.3%) типам. Немногочисленны аркто-бореальные виды – (7.2%). К ним относятся Hermannia (Heterohermannia) reticulata (Thörell, 1871) , Sphaerozetes (Sphaerozetes) arcticus (Hammer, 1952), Acrotritia loricata (Rathke, 1799), Propelops rossicus (Shaldybina, 1971) (=Hammeria rossica Shaldybina, 1971) , Svalbardia paludicola (Thor, 1930) , Edwardzetes (Edwardzetes) edwardsi (Nicolet, 1855), Diapterobates notatus (Thorell, 1871), Mela-nozetes sellnicki (Hammer, 1952) . Циркумполярными из них являются H. reticulata, S. arcticus, D. notatus . В изученных тундровых фаунах арктические виды пока не найдены, в отличие от субарктической тундры Сибири. Среди тундровых видов Сибири выявлены «северо-сибирские», условные эндемики Таймыра и Чукотки [33, 34]. Следует отметить, что материковые тундры европейской части России мало изучены. Возможно, при дальнейших исследованиях здесь будут обнаружены арктические виды орибатид. В целом по видовому составу ориба-тид равнинные и горные субарктические тундры близки к бореальным сообществам.
Список видов орибатид арктических островов и архипелагов (включая Шпицберген) составляет 100 видов из 55 родов и 37 семейств. В структуре фауны ведущими являются семейства Brachichthoniidae, Camisiidae, Oppiidae. К голарктическим (в том числе встречающимся в некоторых регионах за пределами Голарктики) относится 53.0% видов, к палеарктическим – 29.8%. Соотношение групп видов с разными типами долготного распространения сходно с таковым в тундровой зоне. Несколько повышается доля космополитов и полукосмополитов (12.6%), в то же время немного ниже доля европейских видов (3,4%). Один вид – Autogneta kaisilai (Karppinen, 1967) является условным эндемиком Шпицбергена. По типу широтного распределения преобладают полизональные виды (70.2 %). На островах выделяется комплекс арктических видов: Ceratozetes (C.) spitsbergensis Thor, 1934 , Murcia setiger (Trägårdh, 1910) , Oribatella (Oribatella) arctica (Thor, 1930) , Iugoribates gracilis ( Sellnick, 1944) , Autogneta kaisilai (Karppinen, 1967). К циркумполярным относятся O. arctica, I. gracilis.
С продвижением на север (от таежной зоны к арктическим островам) сохраняется ведущее значение семейств Oppiidae, Suctobelbidae, Brachich-thoniidae, Camisiidae, Ceratozetidae. Наблюдается общая тенденция снижения видовой насыщенности этих семейств в тундровой зоне и на арктических островах, однако доля участия некоторых семейств в структуре фауны может при этом увеличиваться. Это относится, прежде всего, к семействам Brachi-chthoniidae и Camisiidae. Из семейства Brachich-thoniidae увеличивается доля р. Liochthonuis. А семейства, которые относились в таежной зоне к ведущим, теряют свое значение в сложении фауны с продвижением на север. Так, видовое богатство семейства Liacaridae значительно убывает в тундровой зоне, на арктических островах виды этого семейства не найдены. Род Liacarus, представленный в таежной зоне пятью видами, в равнинной тундре не зарегистрирован; один вид (Liacarus (Dorycranosus) neonominatus Subias, 2004) обнаружен в горных тундрах Полярного Урала. В тундровой зоне остаются виды рода Adoristes указанного семейства. Видовое богатство семейства Phthira-caridae снижается в тундрах и на арктических островах. Семейство Damaeidae относится к числу ведущих в тайге и тундровой зоне, однако на островах теряет свои позиции. Из семейства Achipterii-dae, относящегося к числу богатых видами семейств в таежной зоне, в Арктике присутствует один вид – Parachipteria punctata (Nicolet, 1855). Сходная картина наблюдается для семейства Ma-laconothridae: из девяти видов, распространенных в таежной зоне, на арктических островах представлен только один – Malaconothrus (Malaconothrus) egregius (Berlese, 1904).
В таежной зоне доли первых десяти семейств, названных в таблице, составляют от 3,1 до 6,8 %, т.е. отмечается равномерное распределение числа видов между ведущими семействами. Доли наиболее богатых видами пяти семейств в тундровой зоне – от 10 до 15%, пяти семейств на арктических островах – от 5 до 10%.
Характерной чертой фауны орибатид европейского Севера является высокая степень сходства с сибирской фауной. Около 10% видов встречаются только на европейской территории России. Ареалы большинства видов простираются восточнее Уральского хребта.
Некоторые виды имеют дизъюнктивное распространение по территории Евразии. Выделяется три типа дизъюнкций: сибирская, западно-сибирская и восточно-сибирская. Виды с сибирской дизъюнкцией ареала, помимо европейской части России, зарегистрированы на Дальнем Востоке, при этом отсутствуют на территории всей Сибири: Liochtho-nius (Liochthonius) lapponicus (Trägardh, 1910), Noto-phthiracarus (Calyptophthiracarus) pavidus (Berlese, 1913), Phthiracarus (Archiphthiracarus) globosus (Koch, 1841), Mesotritia (Mesotritia) flagelliformis (Ewing, 1909) (=Mesotritia testacea Forsslund, 1963), Trhypochthoniellus longisetus (Berlese, 1904), Nan-hermannia (Nanhermannia) elegantula (Berlese, 1913), Eueremaeus quadrilamellatus (Hammer, 1952), Carabodes (Klapperiches) minusculus (Berlese, 1923), Lauroppia falcata (Paoli, 1908), Suctobelbella (Sucto-belbella) longicuspis Jacot, 1937, Suctobelbella (Suc-tobelbella) perforata (Strenzke, 1950), Ameronothrus oblongus (Sitnikova, 1975), Oribatula (Oribatula) pan-nonica (Willmann, 1949), Propelops rossicus (Shaldy-bina, 1971), Euzetes globulus (Nicolet, 1855), Eupelops plicatus (Koch, 1839), Scutozetes lanceolatus (Hammer, 1952), Galumna (G.) dimorpha Krivolutskaja, 1952, Pi-logalumna crassiclava crassiclava (Berlese, 1914).
Для видов Hafenrefferia gilvipes (Koch, 1839), Ramusella (Ramusella) clavipectinata (Michael, 1885), Banksinoma setosa Rjabinin, 1974, Oribatula (Zy-goribatula) glabra (Michael, 1890) (= Zygoribatula pro-pinqua Oudemans, 1902) отмечена дизъюнкция ареала в Западной Сибири, их ареал продолжается на территории Восточной Сибири и Дальнего Востока. Восточно-сибирская дизъюнкция имеется в ареалах видов Phthiracarus (Archiphthiracarus) piger (Scopoli, 1763); Nothrus pratensis Sellnick, 1928; Camisia (Ensicamisia) lapponica (Trägardh, 1910); Trimalaconothrus (Tyrphonothrus) maior (Berlese, 1910) (= Malaconothrus novus Sellnick, 1921); Nan-hermannia (Nanhermannia) dorsalis (Banks, 1896) (= Nanhermannia coronata Berlese, 1913); Autogneta (Autogneta) longilamellata (Michael, 1885); Suctobel-bella (Suctobelbella) palustris (Forsslund, 1953); Cym-baeremaeus cymba (Nicolet, 1855); Oribatula (Zy-goribatula) frisia (Oudemans, 1900); Diapterobates oblongus (L. Koch, 1879).
На европейском Северо-Востоке обнаружены виды, основной ареал которых находится в Сибири и на Дальнем Востоке. Так, вид Nothrus parvus Sitnikova, 1975, распространенный в Восточной Палеарктике, найден в средней тайге Республики Коми [18]. Данный вид отмечался также на Кавказе [31], Украине [35]. Вид Heminothrus (Platynothrus) sibiricus (Sitnikova, 1975) обнаружен на Северном Урале [16]. Это голарктический вид, в Палеарктике распространен в ее восточной части [3].
Находки ряда видов на Севере представляют интерес в зоогеографическом плане. Обращают на себя внимание виды с широтным разрывом ареала. Это комплекс так называемых «южных» видов. В Европе их основной ареал располагается в более низких широтах. К ним относится голарктический Trimalaconothrus (Trimalaconothrus) tardus (Michael, 1888), который, как указывает Л. Субиас [3], в пределах Палеарктики на севере не встречается. Распространен этот вид в Центральной Европе, на Кавказе, а также на Дальнем Востоке [2, 31, 36]. Находка данного вида на Полярном Урале [23] – пока единственная на севере Палеарктики. В еловых сообществах горно-лесного пояса Северного Урала найден степной вид Birsteinius perlongus (Krivolutsky, 1965) [16]. Представители рода Birstein-ius ( Krivolutsky, 1965) обычны в степях и полупустынях Палеарктики. B. perlongus распространен в степной зоне России: как европейской части, так и Сибири. Единичные находки известны из зоны хвойно-широколиственных лесов (Московская область) [2]. В Ухтинском районе Республики Коми зарегистрирован южный вид – Scutovertex sculptus Michael, 1879 (= Scutovertex (Neoscutovertex) rugosus Mihelčič, 1957).
В таежной зоне имеются единичные виды, основной ареал которых находится в более низких широтах. Севернее границы смешанных хвойно-ши- роколиственных лесов реже встречаются виды Diapterobates dubinini (Shaldybina, 1971), Ori-batula (Oribatula) pannonica Willmann, 1949, Phthiracarus (Phthiracarus) jacoti Feider y Suciu, 1958, Liebstadia humerata (Sellnick, 1928), Punctoribates (Punc-toribates) minimus (Shaldybina, 1969).
Есть виды, которые на европейской части России и в сопредельных странах распространены южнее бореальной зоны, а за Уральским хребтом – присутствуют на севере Сибири и Дальнем Востоке. Палеарктический вид Mycobates (Mycobates) monodactylus (Shaldybina, 1970), обнаруженный на Полярном Урале [22], обитает в зоне широколиственных лесов. В то же время вид встречается на севере Западной Сибири, а также на Алтае, в Иркутской области и на Сахалине [34, 36, 37]. Палеарктический вид Mycobates (Calyptozetes) patrius (Shaldybina, 1970) , находка которого сделана на Полярном Урале, отмечался на территории Украины [35]; распространен на Ямале, Таймыре, Чукотке [34], в горной тундре Хабаровского края [36]. На Северном и Полярном Урале обнаружен вид Cam-pachipteria nivalis (Hammer, 1952), область распространения которого охватывает Украину, Беларусь, Киргизию, север Восточной Сибири (Таймыр, Северную Эвенкию) и Дальний Восток.
Заключение
Для фауны орибатид Севера характерна относительная бедность таксономического состава по сравнению с более низкими широтами и западными регионами. С продвижением от таежной зоны к арктическим островам сохраняется ведущее значение семейств Oppiidae, Suctobelbidae, Brachichthoniidae, Camisiidae, Ceratozetidae.
По типу долготного распространения преобладают виды с широкими ареалами: голарктические и палеарктические. В тундровой зоне и на арктических островах повышается доля голарктических видов и снижается – палеарктических и европейских, по сравнению с таежной зоной. Отмечена высокая степень сходства с сибирской фауной.
По типу широтного распределения выделены арктические, аркто-бореальные, бореальные, тем-ператные и полизональные виды. Основу фауны составляют полизональные и температные виды. Группа бореальных видов весьма малочисленна. Типично арктические виды зарегистрированы на арктических островах. В субарктической тундре не обнаружены виды, нижняя граница распространения которых не выходит за пределы тундровой зоны. По видовому составу орибатид равнинные и горные субарктические тундры близки к бореальным сообществам. Фауна орибатид европейского Севера в целом имеет бореальный облик.
Настоящая работа выполнена при поддержке Программы ОБН РАН «Биологические ресурсы России: оценка состояния и фундаментальные основы мониторинга» (Проект 09-Т-4-1003).
Список литературы Таксономическое разнообразие и ареалогия орибатид (Oribatei) европейского севера России
- Криволуцкий Д.А. Становление биогеографии почвы/Вестн. Моск. ун-та. Сер. 5. География, 2004. № 6. С. 10-16.
- Панцирные клещи: Морфология, развитие, филогения, экология, методы исследования, характеристика модельного вида Nothrus palustris C.L. Koch, 1839./Д.А.Криволуцкий, Ф.Лебрен, М.Кунст и др. М.: Наука, 1995. 224 с.
- Subias L.S. Listado sistematico, sinonimico y biogeografico de los Acaros Oribatidos (Acariformes, Oribatida) del mundo (1758-2002)//Graellsia, 2004. Vol. 60. P. 3-305.
- Бызова Ю.Б., Уваров А.В., Губина В.Г. и др. Почвенные беспозвоночные беломорских островов Кандалакшского заповедника. М.: Наука, 1986. 312 с.
- Высоцкая С.О., Буланова-Захваткина Е.М. Панцирные клещи из гнезд грызунов и насекомоядных Ленинградской области//Паразитол. сб. Зоол. ин-та АН СССР, 1960. Т. 19. С. 194-219.
- Ситникова Л.Г. Краткий обзор фауны панцирных клещей (Acariformes, Oribatei) Ленинградской области//Тр. ЗИН АН СССР, 1962. Т. 31. С. 429-452.
- Бызова Ю.Б. Фауна почвенных ногохвосток и клещей севера среднеевропейской тайги//Pedobiologia, 1964. Bd. 3. S. 286-303.
- Воронова Л.Д. Почвенная фауна южной тайги Пермской области//Почвенная фауна Се-верной Европы. М.: Наука, 1987. С. 59-65.
- Животный мир Кировской области (беспозвоночные животные). Дополнение: сборник статей. Т.5. Киров: Изд-во ВГПУ, 2000. 232 с.
- Криволуцкий Д.А. Панцирные клещи из окрестностей Беломорской биологической станции Московского университета//Вестн. Моск. ун-та, 1966а. № 1. С. 42-45.
- Ласкова Л.М. Биоразнообразие панцирных клещей Карелии//Биогеография Карелии. Петрозаводск, 2001. С. 125 -132.
- Поспелов Л.Е., Солнцева Е.Л., Чугунова М.Н. Комплексы микроартропод в разных типах леса в подзоне северной тайги европейской части СССР//Проблемы почвенной зоологии. Минск: Наука и техника, 1978. С.189-190.
- Новожилова Э.П. Панцирные клещи (Oribatei) из гнезд млекопитающих Коми АССР//Кровососущие членистоногие Европейского Севера. Петрозаводск, 1980. С.164-168.
- Михальцова З.А. Новый вид панцирного клеща рода Passalozetes (Acariformes, Passalozetidae) из средней тайги Коми АССР//Зоол. журн., 1982. Вып. 11. С. 1756-1759.
- Друк А.Я., Вилкамаа П. Микроартроподы верховых болот севера европейской части СССР//Биология почв Северной Европы. М.: Наука, 1988. С. 190-198.
- Мелехина Е.Н. К фауне панцирных клещей (Acari: Oribatida) Печоро-Илычского заповедника//Труды Печоро-Илычского заповедника (Вып. 14). Сыктывкар, 2005а. С.113-117.
- Мелехина Е.Н. Разнообразие фауны и географическое распределение панцирных клещей (Oribatida) таежной зоны европейского Северо-Востока//Закономерности зональной организации комплексов животного населения на европейском Северо-Востоке. Сыктывкар, 2005б. С. 257-273. (Тр. Коми научного центра УрО РАН, № 177).
- Мелехина Е.Н., Криволуцкий Д.А. Список видов панцирных клещей Республики Коми. Сыктывкар, 1999. 24 с.
- Karppinen Eero et Krivolutsky D.A. List of oribatid mites (Acarina, Oribatei) of northern palaearctic region. 1. Europe//Acta ent. fenn. 1982. Vol. 41. 18 p.
- Криволуцкий Д.А. Панцирные клещи в почвах тундры//Pedobiologia, 1966б. Bd. 6. № 3. С. 277-280.
- Мелехина Е.Н. Влияние загрязнения почв на фауну панцирных клещей в Воркутинской тундре//Криопедология`97: Тез. докл. Междунар. конф. Сыктывкар, 1997. С. 176-177.
- Биоразнообразие экосистем Полярного Урала/Отв. ред. М.В. Гецен. Сыктывкар, 2007. 251 с.
- Сидорчук Е.А. К фауне панцирных клещей (Acariformes, Oribatida) Полярного Урала//Зоол. журн., 2009. Т. 88. № 7. С. 1-9.
- Криволуцкий Д.А., Калякин В.Н. Микрофауна почв в экологическом контроле на Новой Земле//Новая Земля. М., 1993. Т.2. С. 125-131.
- Криволуцкий Д.А., Дроздов Н.Н., Лебедева Н.В., Калякин В.Н. География почвенных микроартропод островов Арктики//Вестн. Моск. ун-та. Сер. 5. География, 2003. № 6. С. 33-40.
- Лебедева Н.В., Лебедев В.Д., Мелехина Е.Н. Новые данные о фауне орибатид (Oribatei) Шпицбергена//Доклады Академии наук, 2006. Т. 407. № 6. С. 845-849.
- Coulson S.J. Terrestrial and Freshwater Invertebrate Fauna of the High Arctic Archipelago of Svalbard//Zootaxa 1448. 2007. P. 41-68.
- Городков К.Б. Типы ареалов насекомых тундры и лесных зон европейской части СССР//Ареалы насекомых европейской части СССР. Л., 1984. С. 3-20.
- Lundqvist L. Bibliografi och checklist over Sveriges oribatider (Acari: Oribatei) Ent. Tidskr, 1987. Vol. 108. P. 3-12.
- Niemi R., Karppinen E., Uusitalo M. Catalogue of the Oribatida (Acari) of Finland//Acta Zool. Fenn, 1997. Vol. 207. P. 1-39.
- Shtanchaeva U. Ya. Catalog of oribatid mites of the Caucasus (Acari, Oribatida)//Acarina, 2001. Vol. 9. Nо. 2. P. 177-221.
- Danks H.V. Arctic artropods. A reviev of systematics and ecology with particular reference to the North American Fauna//Entomological Soc. Canada. Ottawa, 1981. 608 pp.
- Ананьева С.И., Криволуцкий Д.А., Чернов Ю.И. Панцирные клещи (Oribatei) подзоны типич-ных тундр Западного Таймыра//Биогеоценозы таймырской тундры и их продуктивность. Л., 1973. Вып. 2. С. 148-151.
- Гришина Л.Г. Панцирные клещи севера Сибири//Членистоногие Сибири и Дальнего Востока. Новосибирск: Наука, 1985. С. 14-23.
- Karppinen E., Melamud V., Mico L., Krivolutsky D.A. Further information on the oribatid fauna (Acarina, Oribatei) of the northern palaearctic region: Ukraina and Czechoslovakia//Entomol. Fenn. 1992. Vol. 3. P. 41-56.
- Рябинин Н.А., Паньков А.Н. Каталог панцирных клещей Дальнего Востока России. Часть II. Континентальная часть Дальнего Востока. Владивосток-Хабаровск: ДВО РАН, 2002. 92 с.
- Паньков А.Н., Рябинин Н.А., Голосова Л.Д. Каталог панцирных клещей Дальнего Востока России. Часть I. Каталог панцирных клещей Камчатки, Сахалина и Курильских островов. Владивосток -Хабаровск: Дальнаука, 1997. 87 с.
