Теоретическое обоснование и экспериментальное подтверждение антибактериальных и противогрибковых свойств природного пептида

Автор: Тихонов С.Л., Мерзлякова Н.В., Шихалев С.В., Тихонова Н.В., Смирнова А.В., Тихонова М.С.
Журнал: Вестник Восточно-Сибирского государственного университета технологий и управления @vestnik-esstu
Статья в выпуске: 2 (89), 2023 года.
Бесплатный доступ
Статья посвящена теоретическому обоснованию антибактериальной и противогрибковой активности природного пептида mpR2, выделенного из пепсинового гидролизата молозива коров, и экспериментальному подтверждению его эффективности in vitro. Выделенный полипептид состоит из 60 аминокислотных остатков и имеет молекулярную массу 23 кДа. Моделирование показало, что пространственная структураmpR2 представлена α-спиралью. Пептид имеет гидрофобный домен, его суммарный заряд составляет +2, а изоэлектрическая точка находится на уровне 8,9. Полученные физические характеристики позволили предположить, что выделенный пептид обладает антибактериальной и противогрибковой активностью, что было подтверждено в эксперименте in vitro. Установлено, что идентифицированный пептид mpR2 обладает антибактериальной активностью в отношении тест-штаммов бактерий грамотрицательной бактерии E. coli ATCC 25922 и грамположительной бактерии B. Subtilis, противогрибковой активностью в отношении дрожжевого диплоидного грибка C. Albicans, но более низкой в сравнении с активностью антибиотика и противогрибкового препарата.
Пептиды, молозиво коров, антимикробная активность, противогрибковая активность, структура пептидов, α-спиральная пространственная структура
Короткий адрес: https://sciup.org/142238242
IDR: 142238242 | УДК: 543.645.6 | DOI: 10.53980/24131997_2023_2_40
Theoretical rationale and experimental study confirmation of antibacterial and antifungal properties of natural peptide
This article focuses on theoretical explanation of the antibacterial and antifungal activity of the natural peptide mpR2 isolated from pepsin hydrolysate of cow colostrum, and experimental confirmation of its effects in vitro. The isolated polypeptide consisted of 60 amino acid residues and had a molecular weight of 23 kDa. Modeling showed that the spatial structure of mpR2 is represented by an α-helix. The peptide has a hydrophobic domain, its total charge is +2, and the isoelectric point is at the level of 8.9. The obtained biophysical characteristics suggested that the isolated peptide has antibacterial and antifungal activity, which was confirmed by the disco-diffusion method. It was found that the identified peptide mpR2 has antibacterial activity against test strains of bacteria of gram-negative bacterium E.coli ATCC 25922 and gram-positive bacterium B. Subtilis, as well as antifungal activity against yeast diploid fungus C. Albicans, but lower in comparison with the activity of synthetic antibiotics and antifungal drugs.
Текст научной статьи Теоретическое обоснование и экспериментальное подтверждение антибактериальных и противогрибковых свойств природного пептида
Антимикробные пептиды (AMП) представляют собой разнообразный класс молекул, встречающихся в природе и экспрессирующихся во всех организмах, включая млекопитающих [1]. Они являются незаменимыми компонентами врожденного иммунитета и обладают широким спектром активности против бактерий, грибков, дрожжей, вирусов, а также онкоклеток [2]. Как правило, АМП имеют относительно небольшую длину - от 10 до 100 остатков аминокислот, являются катионными, могут быть амфифильными, гидрофобными и гидрофильными [3]. АМП кодируются в геноме в виде препропептидов, где сигнальный пептид находится на N -конец, а домен антимикробного пептида находится на С -конце [4]. АМП являются альтернативой традиционным антибиотикам, так как обладают широким спектром действия и у микроорганизмов не развивается резистентность к пептидам. Первый АМП-препарат грамицидин А, выделенный из почвенных бактерий Bacillus subtilis , был произведен промышленным способом в 1940-х гг. Низин из Lactococcus lactis впервые был представлен в 1953 г. в качестве противомикробного средства и сейчас находит широкое применение в качестве безопасного пищевого биоконсерванта. Биоинженерные варианты низина эффективны при инфекционных заболеваниях, вызванных S. pneumoniae, энтерококками , C. Difficile , а также они выступают синергетиками по отношению к антибиотикам, таким как ципрофлоксацин и ванкомицин [5]. Пептиды полимиксин B (открытый в 1955 г.), полимиксин E (более известный как колистин, открытый в 1962 г.) и даптомицин (открытый в 2003 г.) используются в качестве антимикробных препаратов [6].
АМП классифицируют на основе их происхождения, свойств последовательности, структурных свойств, биологической активности и молекулярных мишеней воздействия [7]. Поскольку структурная организация АМП играет важную роль в механизме их действия [8], то целесообразно рассмотреть классификацию на основе структуры пептидов. АМП можно разделить на 4 класса в зависимости от их конформационной организации: 1) а-спираль; 2) в-слой; 3) одновременно а-спираль и в-слой (т. е. смешанный тип); 4) без а-спирали и в-слоя [9].
Процентное соотношение существующих пептидов с 1) а-спиралью, 2) в-слоем, 3) а-спи-ралью и в-слоем и 4) без а-спирали и в-слоя пептиды среди известных 3D-структур составляет 69, 12, 16 и 3 % соответственно. Большинство известных АМП являются катионными, незначительное число пептидов несет нейтральный или анионный заряд. Так, АМП с а-спиралью, в-слоем в структуре имеют средний заряд от +2 до +3,60, смешанные пептиды имеют более высокий средний заряд - на уровне + 5,40 [9].
В части структуры аминокислотной последовательности а-спиральные пептиды имеют в составе лизин, лейцин и аланин, тогда как пептиды с в-слоем имеют в составе цистеин и аргинин. Примечательно, что все 4 класса содержат высокие доли глицина, что позволяет предположить, что он важен как для структурной поддержки, так и для гибкости пептидов для выполнения антимикробных функций. а-спиральные пептиды представляют собой самый большой класс АМП и являются наиболее изученными [10].
Ввиду того что в статье описаны результаты исследования а-спиральных противомик-робных пептидов, целесообразно остановиться на механизме их действия.
Многие а-спиральные противомикробные пептиды линейны и амфипатичны, состоят из катионных и гидрофобных аминокислот, пространственно разделенных на противоположных сторонах спирали [10]. Яркими примерами этого класса являются кателицидины млекопитающих [11]. Эти пептиды проявляют сильное сродство к мембранам, тем самым ставя под угрозу стабильность β-слоя, нарушая организацию мембраны и (или) формируя поры. Первоначально пептиды находятся параллельно поверхности мембраны клетки, затем при увеличении концентрации вращаются вокруг мембраны, чтобы внедриться в мембрану под определенным углом наклона. Человеческий кателицидин LL-37 может принимать частично спиральную структуру в растворе, следовательно, заставляя его олигомеризоваться с другими пептидами, чтобы скрыть гидрофобную поверхность. Помимо индивидуальной активности, спиральные пептиды с различными последовательностями могут действовать синергически, что приводит к усилению цитолитического и антибактериального эффектов [12].
Путем посттрансляционной модификации зрелый (активный) биопептид образуется путем протеолитического расщепления белка [13].
Молочные белки являются легкодоступным источником ценных пептидов с разнообразными типами биологической активности, такими как антимикробная, гипотензивная, антиоксидантная, антитромботическая и иммуномодулирующая [14].
Так, авторами [15] в результате двустадийного гидролиза соевого белка пепсином и трипсином получены пептиды с антиоксидантными свойствами, установлены оптимальные параметры гидролиза соевого белка для каждого фермента. Ученые [16] доказали, что биопептиды могут быть выделены из мяса и мясопродуктов. В исследовании авторов [17] установлены оптимальные параметры гидролиза яичного альбумина для выделения биопептидов.
Цель работы – теоретически обосновать антимикробную и противогрибковую активность природного пептида, выделенного из пепсинового гидролизата молозива коров, и подтвердить его эффективность в эксперименте in vitro .
Материалы и методы исследования
В качестве объекта исследований использован пептид, выделенный из пепсинового гидролизата молозива коров. Пептиду дано условное название mpR2 . Молекулярно-массовое распределение пептида оценивали масс-спектрометрическим методом и идентифицировали методами MALDI-TOF и MS масс-спектрометрии на MALDI-времяпролетном масс-спектрометре Ultraflex («Bruker», Германия). Масс-спектры анализировали с помощью программы Mascot, опция Peptide Fingerprint («Matrix Science», США) с использованием базы данных Protein NCBI. Микросеквенирование осуществляли при помощи секвенатора MiSeq (Illumina, США) и системы для высокопроизводительного секвенирования (PGM Ion Torrent, США).
Моделирование пространственной структуры выделенного пептида осуществляли с помощью программы молекулярного моделирования Schrodinger Maestro (США).
Антимикробную активность пептида исследовали диско-диффузионным методом на грамположительных и грамотрицательных бактериях. В качестве тест-штаммов использовали Escherichia coli и грамположительную бактерию Bacillus subtilis .
Культивирование штаммов бактерий проводили на плотной питательной среде LB (агар – 1,5 %, триптон – 1, дрожжевой экстракт – 0,5, NaCl – 1 %) и жидкой питательной среде LB (триптон – 1 %, дрожжевой экстракт – 0,5, NaCl – 1 %) при температуре 37 ºC.
Диффузионный метод определения антимикробной активности гидролизатов заключался в следующем. Тест-штамм высевали на агаризованную питательную среду газоном, и одновременно на газон помещали пептид. В качестве контроля использовался бумажный диск с питательной средой, в качестве препарата сравнения – диск с антибиотиком (канамицин из стандартного набора). Чашки Петри инкубировали при температуре, соответствующей оптимальной температуре роста каждого тест-штамма микроорганизма, в течение 24,0 ± 0,5 ч. Результаты учитывались по наличию и размеру (в мм) прозрачной зоны отсутствия роста микроорганизмов вокруг диска.
Результаты исследования и их обсуждение
На рисунке представлены двухмерная (а) и трехмерная (б) структуры пептида mpR2 .

а
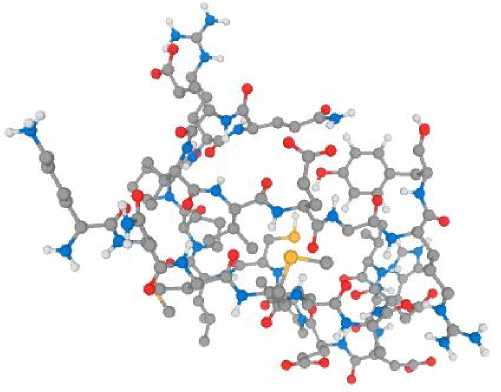
б
Рисунок – Двухмерная (a) и трехмерная (б) структуры пептида mpR2
При исследовании первичной структуры пептида с условным названием mpR2 установлено, что он состоит из 60 аминокислот и в следующей последовательности: LRQLSVVVAYKGKDVGLNDCEEADRHKSSHRDEVSSFRRNSYSIYENHGP SAKCAREVGR, и имеет молекулярную массу 23 кДа. Согласно базе данных, Protein NCBI сходен с пептидом под названием « EV861652 protein, sus scrofa », функции которого не исследованы.
Пептид mpR2 наряду с другими аминокислотами состоит из 5 аминокислотных остатков лизина, 2 лейцина и 5 аланина, что согласуется с базой данных APD3, состоящей из более чем 3 000 записей о структурно-аннотированных природных пептидах, из которой следует, что антимикробные α-спиральные пептиды имеют в составе лизин, лейцин и аланин [18].
По данным [19], в состав антимикробных пептидов входят специфические аминокислоты, такие как аргинин, глицин и гистидин. Их антимикробная активность обеспечивается водородными связями или силами Ван-дер-Ваальса между пептидами и мембранными липидами, независимо от водородных связей между остатками.
В антимикробном пептиде, который проявляет активность против бактерий и вируса SARS-CoV-2, по данным [20], присутствуют аминокислотные последовательности VV, VA и AK, которые имеются в исследуемом пептиде mpR2 .
При моделировании 2-и 3-D структур установлено, что пептид mpR2 относится к α-спиральным и имеет заряд +2, гидрофильность (гидрофобность) – +73,70 Ккал×моль-1, что позволяет сделать предположение о его антибактериальных свойствах, и согласуется с исследованием [21]. В исследовании [22] установлено, что АМП обычно содержат несколько основных сайтов, которые придают им суммарный положительный заряд при изоэлектрической точке близкой к нейтральной или слабощелочной, что подтверждается результатами наших исследований. Так, у пептида mpR2 изоэлектрическая точка на уровне 8,90 и имеется гидрофобный домен. По данным [23], электростатический заряд является еще одним движущим фактором, который влияет на притяжение пептида к микробной клетке и сворачивание пептида на границе раздела липид-пептид и большинство антимикробных пептидов имеют катионную природу с суммарным положительным зарядом от + 2 до + 9.
Теоретическое обоснование антимикробного действия пептида согласуется с исследованиями [24], в которых установлено, что гидролиз трипсина и пепсина способствует образованию большинства признанных АМП.
В таблице представлена антимикробная и противогрибковая активность пептида mpR2 .
Таблица
Антимикробная и противогрибковая активность пептида mpR2
|
Наименование образца |
Диаметр зоны лизиса, мм |
||
|
E. coli ATCC 25922 |
B. subtilis |
C. Albicans |
|
|
Пептид mpR2 |
7 |
9 |
5 |
|
Контроль |
0 |
0 |
0 |
|
Антибиотик канамицин |
22 |
24 |
не исследовали |
|
Противогрибковый препарат флуконазол |
не исследовали |
не исследовали |
26 |
Установлено, что пептид mpR2 обладает противомикробной активностью в отношении бактерий E.coli ATCC 25922 и B. Subtilis, а также противогрибковой активностью в отношении дрожжевого гриба C. Albicans . Следует отметить, что исследуемая активность пептида значительно ниже по сравнению с антибиотиком канамицином и противогрибковым препаратом флуканазолом.
Таким образом, теоретически прогнозируемая противомикробная активность пептида mpR2 подтверждена экспериментально.
Заключение
Несмотря на продолжающиеся усилия по разработке новых антибиотиков, возникновение резистентности у бактерий ограничивает эффективность их применения, что имеет значительные последствия для здоровья населения. По этой причине в последние годы исследования все больше фокусируются на выявлении альтернативных стратегий. В этом контексте особое внимание уделяется использованию АМП природного происхождения.
Проведены исследования по изучению взаимосвязи между структурной организацией и активностью АМП, вследствие чего из трипсинового гидролизата молозива коров выделен пептид с условным названием mpR2 , который идентифицирован по базе данных Protein NCBI как пептид «EV861652 protein, sus scrofa» с функциями, которые ранее не были исследованы.
На основании идентификации аминокислот в пептидной последовательности и определении их числа, а также на основании молекулярной массы пептида, заряда, изоэлектрической точки и гидрофильности сделано предположение, что исследуемый пептид относится к антимикробным катионным α-спиральным пептидам.
Экспериментально доказано, что исследуемый пептид обладает противомикробным и антигрибковым действием, но более низким в сравнении с антибиотиком и противогрибковым препаратом. Следовательно, можно рекомендовать использование исследуемого пептида в качестве дополнительного антибактериального и противогрибкового средства при условии доказательства его биодоступности и отсутствия токсичности.
Список литературы Теоретическое обоснование и экспериментальное подтверждение антибактериальных и противогрибковых свойств природного пептида
- Seyf R., Abarghooi F., Tahereh K. et al. Antimicrobial peptides (AMPs): roles, functions and mechanism of action // Int. J. Pept. Res. Ther. 2020. – P. 1451–1463.
- Huan Y., Kong Q., Mou H. et al. Antimicrobial peptides: classifcation, design, application and research progress in multiple felds // Front Microbiol. 2020. – N 11. – 582779.
- Lei J., Sun L., Huang S. et al. The antimicrobial peptides and their potential clinical applications // Am. J. Transl. Res. 2019. – N 11(7). – P 3919–3931.
- Shi Y., Li C., Wang M. et al Cathelicidin-DM is an antimicrobial peptide from duttaphrynus melanostictus and has wound-healing therapeutic potential // ACS Omega. 2020. – N 5(16). – P. 9301–9310.
- Shin J., Gwak J., Kamarajan P. et al. Biomedical applications of nisin // J. Appl. Microbiol. 2016. – N 120. – P. 1449–1465.
- Moretta A., Scieuzo C., Petrone A.M. et al. Antimicrobial Peptides: A New Hope in Biomedical and Pharmaceutical Fields. Front. Cell. Infect // Microbiol. 2021. – N 11. – 668632.
- Wang G., Li X., Wang Z. APD3: The antimicrobial peptide database as a tool for research and education // Nucleic Acids Res. 2016. – N 44. – P D1087–D1093.
- Koehbach J., Craik D.J. The Vast Structural Diversity of Antimicrobial Peptides // Trends Pharmacol. Sci. 2019. – N 40. – P. 517–528.
- Wang G. Unifying the classification of antimicrobial peptides in the antimicrobial peptide database (Chapter One) // In Methods in Enzymology. – Academic Press: Cambridge, MA, USA, 2022. – Vol. 663. – P. 1–18.
- Perumal P., Pandey V.P. Antimicrobial peptides: The role of hydrophobicity in the alpha helical structure // J. Pharm. Pharmacogn. Res. 2013. – N 1. – P. 39–53.
- Xhindoli D., Pacor S., Benincasa M. et al. The human cathelicidin LL-37 – A pore-forming antibacterial peptide and host-cell modulator // Biochim. Biophys. Acta (BBA)-Biomembr, 2016. – Vol. 1858. – P. 546–566.
- Pino-Angeles A., Leveritt J.M., Lazaridis T. Pore Structure and Synergy in Antimicrobial Peptides of the Magainin Family // PLoS Comput. Biol. 2016. – Vol. 12. – e1004570.
- Zhang L.J., Gallo R.L. Antimicrobial peptides // Curr. Biol. 2016. – N 26(1). – P. R14–R19.
- Guha S., Sharma H., Deshwal G.K. A comprehensive review on bioactive peptides derived from milk and milk products of minor dairy species // Food Prod Process and Nutr., 2021. – Vol. 3. – URL: https://doi.org/10.1186/s43014-020-00045-7
- Соколов Д.В. [и др.] Ферментативный гидролиз соевого белка // Техника и технология пищевых производств. – 2023. – Т. 53, № 1. – С. 86–96. – URL: https://doi.org/10.21603/2074-9414-2023-1-241
- Chernukha I.M., Mashentseva N.G., Afanasev D.A. et al. Biologically active peptides of meat and meat product proteins. Part 2. Functionality of meat bioactive peptides // Theory and practice of meat processing. 2020. – N 5(2). – P. 12–19.
- Жамсаранова С.Д., Лебедева С.Н., Болхонов Б.А. и др. Ферментативная конверсия пищевого белка и оценка антиоксидантной активности пептидов // Вестник ВСГУТУ. – 2021. – № 4(83). – С. 5–14. – DOI 10.53980/24131997_2021_4_5.
- Chen C.H., Lu T.K. Development and Challenges of Antimicrobial Peptides for Therapeutic Applications // Antibiotics. 2020. – N 9. – P. 24.
- Mishra A.K., Choi J., Moon E. et al. Tryptophan-Rich and Proline-Rich Antimicrobial Peptides // Molecules. 2018. – N 23. – 815 p.
- Liscano Y., Onate-Garzon J., Ocampo-Ibanez I.D. In Silico Discovery of Antimicrobial Peptides as an Alternative to Control SARS-CoV-2 // Molecules. 2020. – N 25. – P. 5535.
- Wang Y., Ding Y.,Wen H. et al. QSAR modeling and design of cationic antimicrobial peptides based on structural properties of amino acids // Comb. Chem. High Throughput Screen. 2012. – N 15. – P. 347–353.
- Wang G. The antimicrobial peptide database provides a platform for decoding the design principles of naturally occurring antimicrobial peptides // Protein Sci. 2020. – N 29(1). – P. 8–18.
- Chan D.I., Prenner E.J., Vogel H.J. Tryptophan- and arginine-rich antimicrobial peptides: Structures and mechanisms of action // Biochimica et Biophysica Acta (BBA) – Biomembranes. 2006. – Vol. 1758, Is. 9. – P. 1184-1202.
- Shivanna S.K., Nataraj B.H. Revisiting Therapeutic and Toxicological Fingerprints of Milk-Derived Bioactive Peptides // Food Biosci. 2020. – 100771.
