Теоретическое обоснование возможности отбора перспективных особей в сортовых популяциях самоопылителей на примере сои

Автор: Лукомец В.М., Зеленцов С.В., Мошненко Е.В.
Рубрика: Селекция и семеноводство сельскохозяйственных растений
Статья в выпуске: 2 (186), 2021 года.
Бесплатный доступ
Селекционная практика показывает, что созданные методами синтетической селекции сорта сои передаются на государственное сортоиспытание в поколении F10-F11. Но вновь выведенные сорта не являются абсолютно гомозиготными. Исследования были связаны с определением частот формирования нетипичных и пригодных для отбора перспективных особей в сортах сои, созданных методами синтетической селекции. Исследования проводили в 2019-2020 гг. на центральной экспериментальной базе ФГБНУ ФНЦ ВНИИМК, г. Краснодар. В экспериментах использовали сорта собственной селекции: Селена, Пума, Вита, Ирбис, Барс, Баргузин и Саяна, общим возрастом 11-15 лет с года получения гибридов F1. Для подтверждения практической возможности выделения в сортовых популяциях отличающихся от фенотипической сортовой нормы особей проводили сплошной осмотр посевов всех исследуемых сортов сои. В посевах всех сортов были выявлены особи, которые имели фенотипические отличия от сортовой нормы. Преимущественно, выделенные особи отличались повышенной высотой растений, более мощным габитусом, увеличенным количеством бобов на растении, устойчивостью к полеганию. Факты обнаружения нетипичных особей с улучшенными морфологическими признаками в разновозрастных сортах сои подтверждают возможность индивидуального отбора растений в сортовых популяциях по морфометрическим признакам. Статистическую динамику снижения гетерозиготных особей в нарастающих поколениях в гибридной самоопыляющейся популяции по n-числу парных генов, по которым гипотетически могут различаться родительские формы, рассчитывали по модифицированной формуле С. Бороевича (1984). Установлено, что частоты формирования в сортовых популяциях сои фенотипически отличающихся особей зависят от общего возраста сорта. Частота формирования морфологически отличающихся особей с увеличением количества поколений сорта снижается. Индивидуальный отбор особей с положительными фенотипическими отличиями от сортовой нормы, может быть рекомендован в качестве дополнительного источника перспективного и практически гомозиготного исходного материала для ускоренной аналитической селекции сои.
Соя, аналитическая селекция, сортовая популяция, индивидуальный отбор, динамика гомозиготизации сортовой популяции, частота встречаемости особей
Короткий адрес: https://sciup.org/142227758
IDR: 142227758 | УДК: 633.853.52:581.162.31:631.52 | DOI: 10.25230/2412-608X-2021-2-186-31-40
Theoretical justification of the possibility to select perspective individuals in cultivar populations of self-pollinators on the example of soybean
Breeding practice shows that soybean cultivars developed by synthetic breeding methods are submitted for the state variety testing in the F10-F11 generation. But the newly bred cultivars are not completely homozygous. The studies were related to the determination of the frequencies of formation of atypical and suitable for selection promising individuals in soybean cultivars developed by synthetic breeding methods. The studies were carried out in 2019-2020 at the central experimental base of V.S. Pustovoit AllRussian Research Institute of Oil Crops, Krasnodar. In the experiments, we used cultivars of our own breeding: Selena, Puma, Vita, Irbis, Bars, Barguzin and Sayana with a total age of 11-15 years from the year F1 hybrids were obtained. To confirm the practical possibility of isolating individuals differing from the phenotypic varietal norm in varietal populations, a complete examination of the crops of all studied soybean cultivars was carried out. In the fields of all cul-tivars, individuals were identified that differed from the varietal norm phenotypically. Mostly, the isolated individuals were distinguished by an increased plant height, a more powerful habit, an increased number of beans per plant, and resistance to lodging. The facts of the detection of atypical individuals with improved morphological traits in soybean cultivars of different ages confirm the possibility of individual selection of plants in varietal populations based on morphometric traits. The statistical dynamics of a decrease in heterozygous individuals in increasing generations in a hybrid self-pollinating population in terms of the n-number of paired genes, by which the parental forms can hypothetically differ, were calculated using the modified formula of S. Borojevic (1984). It was found that the frequency of formation of phenotypically different individuals in varietal populations of soybeans depends on the total age of the cultivar. The frequency of the formation of morphologically different individuals decreases with an increase in the number of generations of the cultivar. Individual selection of individuals with positive phenotypic differences from the varietal norm can be recommended as an additional source of promising and practically homozygous starting material for accelerated analytical breeding of soybean.
Текст научной статьи Теоретическое обоснование возможности отбора перспективных особей в сортовых популяциях самоопылителей на примере сои
Введение. Основным методом современной селекции большинства сельскохозяйственных культур в мире остаётся классическая синтетическая селекция, основанная на объединении в гибридном потомстве, за счёт искусственной гибридизации или естественного переопыления, признаков родительских форм. Этот метод позволяет получать в гибридном потомстве максимально широкую вариативность сочетаний родительских, прежде всего количественных, признаков [1; 2; 3; 4; 5]. Главным, и пока непревзойдённым, преимуществом методов традиционной синтетической селекции является возможность выделения и отбора исходного материала по количественным хозяйственно ценными признакам [4; 6]. Однако эти методы селекции имеют ряд 32
объективных ограничений, одним из которых является продолжительность селекционного процесса, от гибридизации до передачи нового сорта на Государственную регистрацию занимающая у самоопыляющихся культур не менее 9–11 поколений/лет [2; 3; 4; 6].
Тем не менее в методологии классической селекции имеются вполне доступные резервы ускорения процесса создания сортов, в частности, методы аналитической селекции, основанные на индивидуальном отборе перспективных по хозяйственно ценным признакам особей в уже имеющихся сортовых популяциях.
Технологической особенностью аналитической селекции является её почти вдвое сокращённая (на 4–5 лет) общая продолжительность селекционного процесса, по сравнению с синтетической селекцией, за счёт исключения этапа гибридизации и отборов в гибридных популяциях F 1 –F 5 . Селекционный процесс самоопыляющихся культур, включая сою, методами аналитической селекции начинается с индивидуального выделения перспективных высокогомозиготных особей в уже имеющихся сортах. Их селекционная проработка начинается с питомника оценки потомств (ПОП) – аналога селекционного питомника (СП) в схеме синтетической селекции. Поэтому общее время селекционного процесса в рамках аналитической селекции будет занимать не более 5–6 поколений/лет (рис. 1).
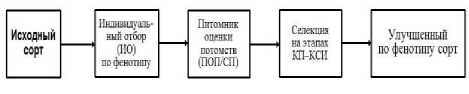
Рисунок 1 – Базовая схема ускоренной аналитической селекции растений-самоопылителей
Однако успех аналитической селекции будет определяться генетической дивер-гентностью родительских форм сорта, среди которых проводится индивидуаль- ный отбор нетипичных особей, фактическим наличием в сортовых популяциях нетипичных особей, а также возможностью их визуального выделения среди фенотипически типичных для сорта растений.
Общее количество генов культурной сои, установленное при секвенировании её генома в 2010 г., насчитывало 46430 шт. [7]. По более поздним (2015 г.) данным, при полном секвенировании генома сои общее количество экспрессирующих генов у сои составило 55582 шт. [8]. При этом у сортов сои фенологическими, морфометрическими, физиологическими и биохимическими методами достоверно фиксируется до 40–50 качественных и количественных признаков. В гибридных потомствах, полученных от скрещивания родительских форм с разным фенотипическим проявлением всех фиксируемых признаков, общее количество сочетаний будет соответствовать формуле (1) [9]:
X = Y n , (1)
где Х – общая комбинативная изменчивость признаков в гибридном потомстве;
Y – количество аллелей генов, кодирующих признак;
n – общее количество фиксируемых признаков.
По мнению С. Бороевича (1984), успех в селекции растений-самоопылителей зависит от времени, в течение которого можно достигнуть гомозиготности по признакам, требующим улучшения, а это обусловлено числом и действием генов, контролирующих данные признаки [1]. При этом только при моногенном наследовании признаков, контролируемых одной парой аллелей ( Y = 2), общая комбинативная изменчивость аллелей, кодирующих те же фиксируемые у сои 40– 50 признаков, в гибридном потомстве может достигать величин 1,09951 × 1012 – 1,1259 × 1015.
В практической синтетической селекции сои, в подавляющем большинстве случаев, в гибридизацию вовлекаются родительские формы, отличающиеся друг от друга намного более чем по одной паре аллельных генов. Поэтому рабочая схема селекционного процесса также обязательно учитывает скорость перехода полученных гибридных гетерозиготных популяций в гомозиготное состояние с учётом предполагаемой внутривидовой генетической дивергентности их родительских форм.
Вполне очевидно, чем больше генетические различия между родительскими формами, тем выше комбинационная изменчивость гибридного потомства, тем ниже скорость гомозиготизации нового сорта, тем выше вероятность формирования особей с отличающимися от сортовой нормы признаками в уже созданных сортах. В связи с этим сформулирована задача теоретически обосновать вероятность наличия нетипичных особей в созданных методами синтетической селекции сортах, а также оценить возможность их практического обнаружения и выделения в реальных сортах сои разного возраста.
Материалы и методы . Исследования проводили в 2019–2020 гг. на центральной экспериментальной базе ФГБНУ ФНЦ ВНИИМК в питомниках предварительного размножения сортов сои. В экспериментах использовали сорта собственной селекции: Селена, Пума, Вита, Ирбис, Барс, Баргузин и Саяна. Посев всех сортов проводили в оптимальные весенние сроки (3-я декада апреля) с густотой 350 тыс. раст./га. Густота стояния растений к уборке на изучаемых сортах составляла 294–309 тыс. раст./га.
В 2019 г. подсчёт и выделение перспективных особей с отличающимися от сортовой фенотипической нормы признаками, проводили на сортах Ирбис и Пума; в 2020 г. – на сортах Селена, Пума, Барс, Вита, Баргузин и Саяна. Общий возраст изучаемых сортов сои на момент проведения индивидуальных отборов устанавливали по количеству поколений (F n ) от года получения гибридов F 1 .
Динамику увеличения частот гомозиготных особей в гибридной самоопыляющейся популяции по n -числу парных генов, по которым гипотетически могут различаться родительские формы, рассчитывали по формуле С. Бороевича [1]:
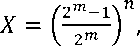
встречае-самоопы-
где Х - коэффициент частоты мости гомозиготных особей;
m - число поколений после ления в F | ;
n - предполагаемое число парных генов, по которым различаются родительские формы.
Поскольку при использовании формулы (2) расчётные значения Х имеют размерность в диапазоне от 0 до 1, этот алгоритм был модифицирован нами до следующего вида:
-
( 7т — 1 \
-
V) , (3)
где коэффициент «100» позволяет перевести частоты образования гомозиготных особей в гибридной популяции в проценты.
Для статистического определения возможных частот встречаемости примесных растений в сортовых самоопыляющихся популяциях сои рассчитывали динамику снижения частот гетерозиготных особей в нарастающих (до общего возраста F15) поколениях модельной гибридной самоопыляющейся популяции. Алгоритм расчёта этих частот X(h) может быть построен на вычитании ожидаемых частот гомозигот от 100 % на основе модифицированной формулы (3) до вида (4):
X(й) = 100 - (100 * (^) ). (4)
Теоретически ожидаемые частоты формирования особей с отличающимися (+/-) от сортовой нормы признаками в 34
сортовых популяциях сои 11-15-летнего общего возраста при стандартной густоте стояния растений сои 300 тыс./га рассчитывали по скомпилированной нами формуле (5):
(Х(Һ)*Е)
где к - теоретически ожидаемая частота встречаемости в популяции отличающихся по морфометрическим признакам особей, шт./га;
Х(һ) - расчётные значения частот гетерозиготных особей (в %) в самоопыляющихся гибридных популяциях разного возраста, рассчитанные по формуле (4);
Е - густота стояния растений (плотность посева), тыс. раст./га.
Результаты и обсуждение . Мировая и отечественная практика показывает, что созданный в результате классической синтетической селекции сорт сои передаётся на государственную регистрацию в поколении F io -F ii [2; 3; 4; 6]. В этом возрасте в подавляющем большинстве случаев сорт фенотипически выровнен и обладает уникальным набором визуально фиксируемых количественных и качественных признаков, позволяющих успешно пройти общепринятую на международном уровне независимую оценку на ООС (однородность, отличимость и стабильность). Уникальная совокупность этих признаков позволяет отличать новый сорт от других сортов, а также обеспечивать поддержание сортовой чистоты в звене первичного семеноводства.
Результаты расчёта по формуле (3) теоретических частот образования и накопления гомозиготных особей в гибридных популяциях сои поколений от F1 до F 15 , полученных от скрещивания родительских форм и гипотетически отличающихся друг от друга n -числом парных генов, для частных случаев n = 1, 5, 10, 50 и 100, представлены в таблице 1.
Таблица 1
Динамика увеличения частот гомозиготных особей в нарастающих поколениях модельной гибридной популяции сои
|
Гибридное поколение, F n |
Степень гомозиготности гибридной популяции при n -числе парных генов, по которым могут отличаться родительские формы, % |
||||
|
при 1 паре генов |
при 5 парах генов |
при 10 парах генов |
при 50 парах генов |
при 100 парах генов |
|
|
F 1 |
0 |
0 |
0 |
0 |
0 |
|
F 2 |
50 |
3,125 |
0,098 |
8,882 × 10-14 |
7,389 × 10-29 |
|
F 3 |
75 |
23,730 |
5,631 |
5,663 × 10-5 |
3,207 × 10-11 |
|
F 4 |
87,5 |
51,291 |
26,308 |
0,126 |
1,588 × 10-4 |
|
F 5 |
93,75 |
72,420 |
52,446 |
3,968 |
0,157 |
|
F 6 |
96,875 |
85,322 |
72,798 |
20,445 |
4,180 |
|
F 7 |
98,438 |
92,428 |
85,429 |
45,502 |
20,704 |
|
F 8 |
99,219 |
96,154 |
92,457 |
67,560 |
45,643 |
|
F 9 |
99,609 |
98,062 |
96,162 |
82,226 |
67,612 |
|
F 10 |
99,805 |
99,027 |
98,064 |
90,687 |
82,242 |
|
F 11 |
99,902 |
99,513 |
99,028 |
95,232 |
90,692 |
|
F 12 |
99,951 |
99,756 |
99,513 |
97,588 |
95,233 |
|
F 13 |
99,976 |
99,878 |
99,756 |
98,787 |
97,588 |
|
F 14 |
99,988 |
99,939 |
99,878 |
99,391 |
98,787 |
|
F 15 |
99,994 |
99,969 |
99,939 |
99,695 |
99,391 |
Анализ расчётных данных, представленных в таблице 1, показывает, что при селекционном процессе на методологической основе синтетической селекции на момент передачи нового сорта на государственное сортоиспытание его генетическая чистота, статистически, не будет 100-процентной.
Например, при различии родительских форм по 1-й паре генов гомозиготность нового сорта на момент передачи на гос-сортоиспытание (F10–F11) теоретически будет составлять 99,8–99,9 %. После завершения обязательного двухлетнего сортоиспытания и достижения общего (от момента получения гибридов F1) возраста нового сорта F12–F13 степень его гомози-готизации достигнет 99,95–99,98 %. При теоретическом генетическом различии родительских форм уже по 10 парам генов степень гомозиготности нового сорта к поколениям F10–F11 будет составлять уже 98,1–99,0 %, к поколениям F12–F13 увеличится до 99,5–99,8 %. Такой уровень гомозиготности сорта вполне соответствует критериям ГОСТ Р 52325–2005 на сортовую чистоту для семян сортов сои высших репродукций (ОС и ЭС), составляющую 99,5 % [10].
Однако при исходных генетических различиях родительских форм сорта по большему количеству генов, например, по 50 парам генов, к поколениям F 10 –F 11 его гомозиготность будет составлять всего 90,7–95,2 %. К поколениям F 12 –F 13 , с такими генетическими различиями между родительскими формами, гомозиготность нового сорта составит всего 97,6–98,8 %.
Безусловно, регулярное проведение сортопрочисток по фенотипу и поддержание сортовой чистоты будут последовательно повышать гомозиготность сорта. Но в первые годы промышленного возделывания нового сорта, непосредственно после его регистрации, ещё будет формироваться небольшая и непрерывно снижающаяся доля растений с некоторыми отличиями от базового фенотипа сортовой популяции.
Таким образом, статистические основания для отбора в популяциях достаточно молодых (до F 15 ) сортов индивидуальных особей сои с отличающимися по отдельным, визуально фиксируемым морфометрическим признакам, например, по высоте растений, габитусу куста, количеству бобов и семян на растении, имеются.
Более наглядно вероятность выделения нетипичных особей может быть представлена в виде частот остаточной гетерозиготности сорта (табл. 2).
Как следует из приведённых в этой таблице данных, рассчитанных по формуле (4), при гипотетическом различии родительских форм по одной паре генов гетерозиготность нового сорта на момент передачи на государственное сортоиспытание (F10–F11) теоретически будет составлять 0,195–0,098 %. После двухлетнего сортоиспытания на сортоучастках и увеличения общего возраста сорта до 12–13 лет степень его гетерозиготности естественно снизится до уровня 0,049– 0,024 %. В поколении F15 естественная гетерозиготность сорта снизится до 0,012 %.
Таблица 2
Динамика снижения частот гетерозиготных особей в нарастающих поколениях гибридной популяции сои
|
Гибридное поколение, F n |
Степень гетерозиготности гибридной популяции при n -числе парных генов, по которым могут отличаться родительские формы, % |
||||
|
при 1 паре генов |
при 5 парах генов |
при 10 парах генов |
при 50 парах генов |
при 100 парах генов |
|
|
F 1 |
100 |
100 |
100 |
100 |
100 |
|
F 2 |
50,000 |
96,875 |
99,902 |
≈100,000 |
≈100,000 |
|
F 3 |
25,000 |
76,270 |
94,369 |
≈100,000 |
≈100,000 |
|
F 4 |
12,500 |
48,709 |
73,692 |
99,874 |
≈100,000 |
|
F 5 |
6,250 |
27,580 |
47,554 |
96,032 |
99,843 |
|
F 6 |
3,125 |
14,678 |
27,202 |
79,555 |
95,820 |
|
F 7 |
1,563 |
7,572 |
14,571 |
54,498 |
79,296 |
|
F 8 |
0,781 |
3,846 |
7,543 |
32,440 |
54,357 |
|
F 9 |
0,391 |
1,938 |
3,838 |
17,774 |
32,388 |
|
F 10 |
0,195 |
0,973 |
1,936 |
9,313 |
17,758 |
|
F 11 |
0,098 |
0,487 |
0,972 |
4,768 |
9,308 |
|
F 12 |
0,049 |
0,244 |
0,487 |
2,412 |
4,767 |
|
F 13 |
0,024 |
0,122 |
0,244 |
1,213 |
2,412 |
|
F 14 |
0,012 |
0,061 |
0,122 |
0,609 |
1,213 |
|
F 15 |
0,006 |
0,031 |
0,061 |
0,305 |
0,609 |
При предполагаемом генетическом различии родительских форм по 10 парам генов степень гетерозиготности нового сорта к поколениям F 10 –F 11 будет составлять 1,94–0,97 %. К поколениям F 12 –F 13 этот показатель снизится до 0,49–0,24 %. При достижении сортом возраста 15 лет его естественная гетерозиготность снизится до 0,06 %.
При генетических различиях родительских форм по 50 парам генов к поколениям F 10 –F 11 гетерозиготность сорта будет составлять 9,31–4,77 %. К поколениям F 12 –F 13 остаточная гетерозиготность нового сорта снизится до 2,41–1,21 %, а к поколению F 15 – до 0,31 %.
Следует отметить, что основные хозяйственно ценные признаки у сои и других культур, такие как урожай семян с единицы площади, масса семян с одного растения, масса 1000 семян, высота растений, вегетационный период, содержание белка и масла в семенах и т.д., являются количественными. При этом включённые в программы искусственной гибридизации родительские формы, в по- давляющем большинстве случаев, отличаются друг от друга более чем одной парой аллельных генов. Поэтому процесс гомозиготизации полученных с их участием гибридных популяций будет ещё более продолжительным, а частичная естественная гетерозиготность будет сохраняться даже при возрасте сорта 13–15 лет.
Объективным ограничением результативности выделения отличающихся от основной сортовой популяции особей является лимитированный генотипами родительских форм суженый внутрисор-товой генофонд признаков исходного материнского сорта. По качественным признакам, которые быстрее всего гомо-зиготизируются в процессе селекции, различий между родственными генотипами в пределах выведенного методами классической селекции синтетического сорта почти не наблюдается. Более широкие возможности для отбора в сортовых синтетических популяциях обеспечивают количественные признаки.
Однако и количественные признаки в пределах сортовой популяции будут иметь ограниченную вариативность, в основном обеспечиваемую расщеплением остаточного количества гетерозиготных особей. Поэтому стандартные алгоритмы вычисления вариативности полигенных признаков и корректное определение частот их трансгрессий в таких генотипически суженных сортовых популяциях уже не применимы.
Тем не менее в сортовых популяциях сои общим возрастом 11–15 лет, на основании расчётных данных таблиц 1 и 2, в результате остаточного расщепления гетерозигот вполне можно ожидать формирования некоторого количества особей с отличающимися (+/-) от сортовой нормы признаками.
Рассчитанные по формуле (5) теоретически ожидаемые частоты встречаемости особей с генетически детерминированными отличиями по отдельным признакам от фенотипической нормы сорта, при частном случае гипотетических генетиче- ских различий родительских форм по одной паре генов (n = 1), без модифицирующего влияния возможного спонтанного переопыления и механического засорения другими сортами, представлены в таблице 3.
Анализ представленных в таблице 3 данных позволяет предположить, что в сортовых популяциях 11-летнего возраста ожидаемая частота встречаемости особей с отдельными улучшенными морфометрическими признаками может составлять не менее 147 шт./га. У 12-летних сортов ожидаемое количество отличающихся от базового фенотипа особей может составлять не менее 73 шт./га. У 13-летних сортов вероятность обнаружения растений с генетически детерминированным превышением отдельных хозяйственно ценных признаков по сравнению с сортовой нормой теоретически может составлять не менее 36 раст./га. У сортов 14-летнего возраста этот показатель может быть не менее 18 раст./га. А у 15-летних сортов частота встречаемости особей с положительными отличиями от фенотипической нормы по отдельному моногенно наследуемому признаку может составлять не менее 9 раст./га.
Таблица 3
Моделирование теоретического ожидания частот формирования особей, не типичных по гипотетически контролируемому одной парой аллелей морфометрическому признаку, в сортовых популяциях сои разного возраста (при средней густоте стояния 300 тыс. раст./га)
|
Общий возраст сорта (поколение F n ) по состоянию на момент отбора |
Теоретически ожидаемая гетерозиготность сортовой популяции при 1-й паре генов, Х(һ) % (см. табл. 2) |
Теоретически ожидаемое количество особей с генетически детерминированными отличиями от сортовой фенотипической нормы отдельного признака, шт./га |
|
|
общее ( k ) |
только с превышением от базового фенотипа сорта, ( k/2) |
||
|
F 11 |
0,098 |
294 |
147 |
|
F 12 |
0,049 |
147 |
73,5 |
|
F 13 |
0,024 |
72 |
36 |
|
F 14 |
0,012 |
36 |
18 |
|
F 15 |
0,006 |
18 |
9 |
В целом биотипы сои с отличающимися от сортовой нормы отдельными фенологическими или морфометрическими признаками, даже при минимальных частотах их встречаемости, должны быть хорошо различимы в сортовых популяциях F 11 –F 15 . Это позволит их индивидуально отобрать для размножения и последующей селекционной проработки ускоренными методами аналитической селекции по остальным хозяйственно ценным признакам.
Поскольку в рамках классической схемы аналитической селекции отбор будет проводиться из уже имеющихся сортов, то особи, выделенные по отдельным морфометрическим признакам в целом должны обладать тесным генетическим родством к исходному сорту. И, следовательно, по большинству признаков, прежде всего по качественным (форма листовой пластинки, окраска цветка, окраска опушения растений, и т.п.), выделенные особи фенотипически будут аналогичны материнскому сорту. Поэтому индивидуальный отбор в сортовых популяциях сои преимущественно будет основан на выделении перспективных особей с улучшенными количественными морфометрическими признаками, например, по высоте растений, повышенному количеству бобов и семян, устойчивости к полеганию и т.п.
Для практического подтверждения возможности выделения и отбора отличающихся от фенотипической сортовой нормы особей из сортов сои в 2019– 2020 гг. был проведён тщательный осмотр питомников предварительного размножения исследуемых сортов сои.
Фенотипически отличающиеся от сортовой нормы особи были выявлены в посевах всех сортов. Эти особи, преимущественно, отличались повышенной высотой, более мощным габитусом и увеличенным количеством бобов на растении, в отдельных случаях – повышенной устойчивостью к полеганию (рис. 2).

а б
Рисунок 2 – Примеры фенотипов нетипичных особей, выделившихся в питомниках предварительного размножения сортов сои, ФГБНУ ФНЦ ВНИИМК, Краснодар, 2020 г.: а – высокорослое растение в сорте сои Селена; б – устойчивое к полеганию растение в сорте сои Барс
Подсчёт всех индивидуально выделенных особей в питомниках предварительного размножения изучаемых сортов сои показал следующие результаты (табл. 4).
Таблица 4
Количество особей, выделившихся по визуальным морфометрическим признакам в питомниках предварительного размножения сортов сои
ФГБНУ ФНЦ ВНИИМК, Краснодар, 2019–2020 гг.
|
Сорт |
Год отбора |
Общий возраст, лет (F n ) |
Учётная площадь отбора, га |
Теоретически ожидаемое кол-во отобранных растений при n = 1 на 1 га |
Выделено, всего |
|||
|
с учётной площади, шт. |
в пересчёте на 1 га, шт. |
к общему кол-ву растений, % |
||||||
|
шт. |
% |
|||||||
|
Ирбис |
2019 |
13 |
0,21 |
36 |
0,024 |
23 |
109,5 |
0,037 |
|
Пума |
«» |
13 |
0,18 |
36 |
0,024 |
19 |
105,6 |
0,057 |
|
Селена |
2020 |
15 |
0,32 |
9 |
0,006 |
5 |
15,6 |
0,005 |
|
Пума |
«» |
14 |
0,39 |
18 |
0,012 |
18 |
46,2 |
0,015 |
|
Барс |
«» |
13 |
0,22 |
36 |
0,024 |
30 |
136,4 |
0,045 |
|
Вита |
«» |
12 |
0,32 |
73,5 |
0,049 |
27 |
84,4 |
0,028 |
|
Баргузин |
«» |
11 |
0,07 |
147 |
0,098 |
7 |
100,0 |
0,033 |
|
Саяна |
«» |
11 |
0,03 |
147 |
0,098 |
5 |
166,7 |
0,056 |
Как следует из представленных в таблице 4 данных, количество выделенных в посевах особей, визуально отличавшихся от фенотипической нормы, в целом снижалось с увеличением возраста сорта.
В 2019 г. у 13-летних на тот период сортов Ирбис и Пума по признакам более мощного габитуса куста с площади 0,21 и 0,18 га было выделено 23 и 19 соответственно фенотипически отличающихся особей. Эти величины составили соответственно 0,037 и 0,057 % от общего количества растений в питомниках
В 2020 г. у самого возрастного (15 лет) сорта сои Селена с площади 0,32 га было выделено всего 9 нетипичных растений с более мощным габитусом, что составило 0,005 % от общего количества растений на учётной площади. Эти величины оказались довольно близки к теоретически рассчитанным для частного случая (n = 1), составлявшим 9 особей и 0,006 % от общей площади соответственно.
У сорта Пума, возраст которого в 2020 г. увеличился до 14 лет в пересчёте на 1 га количество особей, выделенных по признаку более мощного габитуса куста, снизилось почти в 2 раза по сравнению с предыдущим годом и составило 46,2 шт., или 0,015 %. Статистическое моделирование при n = 1 предсказывало вероятность выделения 18 нетипичных растений и 0,012 % от общего количества растений на всей площади.
У 12-летнего сорта Вита фактическая частота выделенных растений в пересчёте на 1 га составила 84,4 шт., или 0,028 %, при теоретическом ожидании при частном случае отличий по одному моногенно наследуемому признаку – 73,5 особей, или 0,049 %.
У 11-летних сортов Баргузин и Саяна с фактической площади 0,07 и 0,03 га было выделено соответственно 7 и 5 растений с более мощным габитусом куста, что в пересчёте на 1 га составило 100,0 и 166,7 растений, или 0,033 и 0,068 % от общего количества растений на всей учётной площади. Теоретическое моделирование для сортовых популяций 11-летнего возраста предполагало формирование 147 растений на 1 га, или 0,098 %.
13-летний сорт Барс по количеству выделенных примесных растений в 2020 г.
несколько отличался от других сортов. Теоретические ожидания объёма примесных растений по этому сорту составляли 36 шт./га, или 0,024 % от общего количества растений на учётной площади. Фактически с учётной площади 0,22 га было отобрано 30 растений, что в пересчёте на 1 га составило более 136 шт., или 0,045 %. Наиболее вероятной причиной увеличенного количества отобранных нетипичных растений могли стать иные критерии отбора. В погодных условиях 2020 г. из-за обильных осадков во 2-й половине июля основная масса растений этого сорта полегла. Поэтому в посевах сорта Барс, в первую очередь, выделились более устойчивые к полеганию растения, которые преимущественно и отбирались (см. рис. 2 б ). В предыдущие годы этот сорт не полегал, поэтому при сортопрочистках этот признак практически не учитывался, что, возможно, повлекло за собой постепенное накопление в сортовой популяции особей с повышенной устойчивостью к полеганию за счёт их более высокой семенной продуктивности.
В целом анализ фактических частот формирования нетипичных растений в сортах разного возраста подтверждает теоретически ожидаемую динамику уменьшения гетерозиготности с возрастанием числа репродукционных поколений. Корреляционная зависимость количества выделенных нетипичных особей от возраста сорта, рассчитанная с использованием алгоритма прямолинейной корреляции Пирсона в целом по всем изучаемым сортам составила r = -0,752 при р -уровне (критерии значимости) этой корреляции р = 0,032, что на 5%-ном уровне является статистически значимым. При коэффициенте детерминации r 2 = 0,566 можно заключить, что именно возраст сорта при не менее чем 56,6 % всех наблюдений является определяющим фактором динамики частот формирования нетипичных особей.
При этом корреляционная взаимосвязь между теоретической моделью при частном гипотетическом случае генетических различий родительских форм по 1-й паре генов (n = 1) и реальными частотами выделения нетипичных особей хотя и была положительной (r = 0,596 и r2 = 0,355), но вполне ожидаемо оказалась статистически незначимой (р-уровень = 0,119). Причина незначимости этой корреляционной связи, вполне очевидно, определялась тем, что на частоты формирования нетипичных особей в сортовых популяциях изучаемых сортов оказывала влияние остаточная гетерозиготность более чем по одной паре генов. К тому же такие морфометрические признаки, как высота растений, габитус куста, количество бобов и семян на растении и устойчивость к полеганию, являются полигенными признаками, кодируемыми неаллельными комплексами генов. Эти количественные признаки имеют определённые вариационные ряды фенотипического проявления, что также вносило статистические шумы в значимость корреляционных отношений.
Выводы . Таким образом, возможность индивидуального отбора растений в сортовых популяциях по морфометрическим признакам, на примере разновозрастных сортов сои, в целом подтверждается.
Частоты формирования в сортовых популяциях сои фенотипически отличающихся особей зависят от общего возраста сорта с момента получения гибридов F 1 . С увеличением количества поколений сорта частота формирования, за счёт остаточного расщепления гетерозигот, морфологически отличающихся особей снижается.
Индивидуальный отбор особей с положительными фенотипическими отличиями от сортотипа может быть рекомендован в качестве дополнительного источника перспективного и практически гомозиготного исходного материала для ускоренной аналитической селекции сои.
Список литературы Теоретическое обоснование возможности отбора перспективных особей в сортовых популяциях самоопылителей на примере сои
- Бороевич С. Принципы и методы селекции растений / Пер. с сербохорв. В.В. Иноземцева. - М.: Колос, 1984. - С. 37-39.
- Scaboo A.M., Chen P., Sleper D.A., Clark K.M. Classical breeding and genetics of soybean // In: Genetics, Genomics and Breeding of Soybean / Ed. by K. Bilyeu, M.B. Ratnaparkhe and Ch. Kole - Science Publishers, USA, 2010. - P. 27-48.
- Ващенко А.П., Мудрик Н.В., Фисенко П.П., Дега Л.А., Чайка Н.В., Капустин Ю.С. Соя на Дальнем Востоке. - Владивосток: Дальнаука, 2010. - С. 124-130.
- Бабич А.О., Бабич-Побережна А.А. Селекщя, виробництво, торпвля i використання со! у свт. - Ки!в: Аграрна наука, 2011. - С. 149-154.
- Acquaah G. Principles of plant genetics and breeding. Second edition - Wiley-Blackwell, A. John Wiley & Sons, UK, 2012. - P. 64-66.
- Зеленцов С.В. Методические основы селекционного процесса у сои и его улучшающие модификации во ВНИИМК (обзор) // Масличные культуры. - 2020. - Вып. 2 (182). -С. 128-143. DOI: 10.25230/2412-608X-2020-2-182-128-143
- Schmutz J., Cannon S.B., Schlueter J. [et al.] [всего 45 авторов]. Genome sequence of the palaeopolyploid soybean // Nature. - 2010. -Vol. 463. - P. 178-183. DOI: 10.1038/natu-re08670
- Liu Z.Z., Yao D., Zhang J., Li Z.L., Ma J., Liu S.Y., Qu J., Guan S.Y., Wang D.D., Pan L.D., Wang D., Wang P.W. Identification of genes associated with the increased number of four-seed pods in soybean (Glycine max L.) using tran-scriptome analysis // Genetics and Molecular Research. - 2015. - Vol. 14 (4). - P. 1889518912. DOI: 10.4238/2015.December.28.39
- Инге-Вечтомов С.Г. Генетика с основами селекции. - М.: Высшая школа, 1989. - С. 41-42.
- ГОСТ Р 52325-2005 Семена сельскохозяйственных растений. Сортовые и посевные качества. Общие технические условия. - М.: Стандартинформ, 2005. - 24 с.
