Териофауна из голоценовых отложений пещеры Каминная (Северо-Западный Алтай)
")
Автор: Васильев С.К., Колясникова А.С.
Журнал: Проблемы археологии, этнографии, антропологии Сибири и сопредельных территорий @paeas
Рубрика: Археология каменного века палеоэкология
Статья в выпуске: т.XXIX, 2023 года.
Бесплатный доступ
Голоценовые слои Каминной пещеры охватывают период времени от мезолита до Средневековья. Из 11 слоев удалось определить более 7 тыс. костных остатков. Они принадлежат как минимум к 33 видам домашних и диких млекопитающих. Также в слоях присутствуют кости рыб, амфибий, птиц и человека. Почти 2/3 костных остатков найдено в слоях 1-3 (от 3,2 тысяч лет назад до этнографического времени). Количество остатков охотничье-промысловых видов более чем в 4 раза превосходит число костей домашних животных. Этим Каминная пещера резко отличается от других пещерных местонахождений Алтая. Очевидно, эта относительно удаленная пещера посещалась в основном группами охотников, реже пастухами. В среднем по слоям среди домашних животных преобладают остатки мелкого рогатого скота (44,3 %), лошади (38,6 %), менее многочисленны остатки коровы (10,8 %) и собаки (4,2 %). Среди диких млекопитающих абсолютно преобладают остатки благородного оленя (48,6 %) и косули (45,7 %). По нескольку десятков костей принадлежат волку, бурому медведю, соболю, единичные кости кабарге, лосю, туру (?), сибирскому горному козлу и архару. Большинство из остатков свиней относится, вероятно, к дикой форме. Почти все кости медведя, судя по их размерам, происходят от мелких взрослых самок. Голоценовый соболь Алтая размерами тела превосходил одновременного ему соболя Средней Сибири. По размерам костей скелета голоценовый марал был на 5,7 % крупнее современного паркового марала. Голоценовые маралы Северо-Западного Алтая (Каминная) и Центрального Алтая (Кучерла) в размерах тела не различались.
Каминная пещера, голоцен, костные остатки, крупные млекопитающие
Короткий адрес: https://sciup.org/145146715
IDR: 145146715 | УДК: 569+903.32 | DOI: 10.17746/2658-6193.2023.29.0081-0089
Theriofauna from the holocene deposits of Kaminnaya cave (Northwestern Altai)
The Holocene layers in Kaminnaya Cave comprise the chronological period from the Mesolithic to the Middle Ages. Over 11,000 bone remains from 11 layers have been identified. They belonged to at least 33 species of domestic and wild mammals. The layers also contained bones offish, amphibians, birds, and humans. Almost 2/3 of bone remains occurred in layers 1-3 (from 3200 BP to ethnographic period). The number of bones which belonged to hunting species was more than four times greater than number of bones of domestic animals, which makes Kaminnaya cave very different from other cave sites in the Altai. Apparently, this relatively remote cave was mostly visited by groups of hunters and less often by shepherds. In terms of domestic animals, on average, the layers contained the following remains (in the descending order): small cattle (44.3 %), horses (38.6 %), cows (10.8 %), and dogs (4.2 %). Remains of red deer (48.6 %) and roe deer (45.7 %) were absolutely predominant among wild mammals. Several dozen bones each belonged to a wolf, brown bear, and sable. Individual bones belonged to musk deer, elk, aurochs (?), Siberian mountain goat, and argali. Most of pig remains probably belonged to their wild form. Judging by their size, almost all bear bones belonged to small adult females. The Holocene sable of the Altai had a larger size of the body than the contemporaneous sable of Central Siberia. Based on the size of skeleton bones, the Holocene red deer was 5.7 % larger than modern park red deer. Holocene red deer in the Northwestern Altai (Kaminnaya cave) and Central Altai (Kucherla) did not differ in body size.
Текст научной статьи Териофауна из голоценовых отложений пещеры Каминная (Северо-Западный Алтай)
Пещера Каминная расположена в среднегорье Северо-Западного Алтая, на высоте 1 100 м над ур. м., в 30 км от Денисовой пещеры. Стационарные раскопки пещеры проводились в 1983–1988 гг. Ю.В. Гри-чаном [Деревянко, Гричан, 1990] и позднее, в 1994– 2004 гг., С.В. Маркиным [Деревянко и др., 1998]. Исследованию фауны крупных млекопитающих из плейстоценовых слоев посвящена отдельная статья [Васильев, Деревянко, Маркин, 2006]. Голоценовая териофауна Каминной пещеры до настоящего времени не была опубликована. Основной объем исследованной остеологической коллекции происходит из раскопок Ю.В. Гричана в 1983–1988 гг. на пред-входовой площадке. Кроме того, были привлечены также менее значительные по объему материалы из раскопок С.В. Маркина 1994 г., охватывающие слои с 6 по 11. Необходимо отметить, что остеологические материалы из голоценовой толщи в конце 1980-х гг. уже были предварительно разобраны и определены Н.Д. Оводовым и автором ( С.В. ). Неопределимые обломки костей в то время подсчитывались, но не сохранялись. К сожалению, все карточки с определениями остались, вероятно, где-то в архивах Н.Д. Оводова. Таким образом, авторы имели дело уже в основном с разложенным по видам материалом, не содержащим неопределимых остатков. Определимые ко стные остатки в количестве 7 018 экз. отнесены к представителям как минимум 33 видов млекопитающих. Кроме того, присутствуют остатки рыб, амфибий, птиц и человека (табл. 1).
Мощно сть рыхлых голоценовых отложений достигает 1,5 м и включает ряд слоев, охватывающих период от мезолита до Средневековья [Деревянко, Дергачёва, 2002]. Основной объем материала (63,7 %) происходит из 1–3 слоев. При этом количество костных остатков с каждым слоем прогрессивно убывает. Так, слой 1 включает 38,7 % всех определимых остатков, слой 2 – 14,7 %, слой 3 – 10,9 %, слой 4 – 8,6 %, слой 5 – 4,3 %, слой 6 – 1,5 %. Слой 1, относящийся к Средневековью, датирован 410 ± 65 л.н., слой 2 (ранние кочевники) – 1 790 ± 65 л.н., слой 3 (позднеафанасьевское время) – 3 120 ± 80 л.н., слой 4 (развитое афанасьево) имеет три датировки – от 4 130 ± 85 л.н. до 4 335 ± 130 л.н. Для 5 слоя, охватывающего период от мезолита до энеолита, получены 4 радиоуглеродные даты – от 5 100 ± 100 л.н. до 6 620 ± 600 л.н. [Деревянко, Гричан, 1990]. Костные остатки из слоев 10 и 11 (раскопки С.В. Маркина 1994 г.) имеют типичную голоценовую сохранность и соответствующий видовой состав (табл. 1).
Тафономические наблюдения. Подавляющее большинство костных остатков из голоценовых слоев являются кухонными отбросами. Все без исключения крупные трубчатые кости расколоты для извлечения костного мозга. Разбиты были даже большинство первых и более половины вторых фаланг маралов и коров. Крайне слабо представлены в коллекции наиболее массивные и габаритные фрагменты крупных ко стей скелета, такие как дистальные отделы бедренных, проксимальные концы плечевой и большеберцовой костей, позвонки маралов, лошадей и коров (табл. 2). Шанс захорониться, быть втоптанным в почвенный горизонт на предвходовой площадке либо под сводами пещеры имели только сравнительно небольшие либо уплощенные кости (такие, как нижние челюсти) или их фрагменты, изолированные зубы (табл. 2). Все остальные остатки от разделанных и съеденных туш неизбежно выпадали из захоронения – растаскивались собаками либо другими хищниками, отбрасывались прочь самим человеком.
У маралов (реже коров) до половины дистальных концов пястных и плюсневых костей, нижние поверх-но сти астрагалов, центральнокубовидных костей, верхние концы лучевых костей и других оказываются затерты в той или иной степени, часто до субовального состояния. Отдельно взятый астрагал, центральнокубовидную кость или уже отбитый нижний конец ме-таподии марала физически крайне трудно сточить до подобного состояния. Скорее всего, для этого использовались еще целые свежие кости, часто с причленя-ющимся к ним набором запястных и заплюсневых костей, удерживающимся на прочных связках. Лучевые, берцовые кости и метаподии разбивались уже после подобных манипуляций. В противном случае трудно представить, как можно сточить до состояния овала крайне прочный в свежем виде нижний конец астрагала либо проксимальный ряд запястных костей. Точно такие же повреждения костей наблюдаются также в материалах Кучерлы и Денисовой пещеры. Аналоги подобной утилизации костей, возможно, известны специалистам из этнографической литературы.
В целом по слоям количество остатков охотничье-промысловых видов более чем в 4 раза превосходит таковое для видов домашних животных. Этим Каминная пещера резко отличается от пещер Денисовой, Кучерлы или Чагырской, где число костей домашних животных в среднем составляет 45, 54 и 71 % соответственно (см. рисунок ). Очевидно, пещера Каминная была относительно удалена от поселений или традиционных маршрутов кочевок и посещалась не так активно, в основном группами охотников, реже пастухами. Доля остатков домашних животных снижается с 29–23 % в слоях 1–2, до 16–17 % в слоях 4–5 и 7 % в слое 6. Исключение составляет 3 слой, где она наиболее высока – 42 %.
Среди домашних животных преобладают остатки мелкого рогатого скота (44,3 %) и лошади (38,6 %), менее многочисленны остатки коровы (10,8 %) и собаки (4,2 %). Среди охотничье-промысловых видов абсолютно доминируют остатки марала (48,6 %) и косули (45,7 %). Другие виды, такие как волк, медведь, представлены 3–4 десятками находок, а кабарга, лось, тур (?), сибирский горный козел, архар – единичными находками.
Таблица 1. Видовой состав и количество костных остатков в голоценовых отложениях Каминной пещеры
Слои
|
Таксоны |
1 |
2 |
3 |
4 |
5 |
6 |
7 |
9 |
10 |
11 |
б/ слоя |
Всего |
|
Homo sapiens |
4/2* |
1/1 |
1/1 |
– |
3/1 |
– |
10/1 |
1/1 |
1/1 |
2/1 |
6/2 |
29/11 |
|
Canis familiaris |
16/1 |
2/1 |
24/1 |
2/1 |
1/1 |
– |
– |
– |
– |
– |
4/1 |
49/6 |
|
Sus scrofa |
7/2 |
6/1 |
– |
– |
10/1 |
– |
– |
1/1 |
– |
1/1 |
6/1 |
31/7 |
|
Equus caballus |
200/5 |
70/3 |
115/3 |
33/1 |
– |
1/1 |
– |
1/1 |
– |
– |
35/2 |
455/16 |
|
Bos taurus |
68/2 |
18/2 |
25/2 |
5/1 |
– |
– |
– |
– |
– |
– |
11/1 |
127/8 |
|
Capra-Ovis (дом.) |
264/5 |
82/3 |
46/6 |
31/3 |
20/2 |
4/2 |
– |
– |
– |
10/2 |
65/2 |
522/25 |
|
Asioscalops altaica |
3/1 |
4/2 |
1/1 |
7/4 |
2/2 |
– |
1/1 |
4/2 |
3/1 |
11/4 |
1/1 |
37/19 |
|
Ochotona sp . |
1/1 |
– |
– |
1/1 |
– |
– |
– |
2/2 |
– |
– |
– |
4/4 |
|
Lepus timidus |
12/2 |
3/1 |
2/1 |
5/1 |
3/1 |
1/1 |
– |
4/1 |
1/1 |
2/1 |
– |
33/10 |
|
Tamias sibiricus |
5/3 |
– |
– |
2/1 |
2/1 |
– |
– |
1/1 |
– |
– |
– |
10/6 |
|
Sciurus vulgaris |
19/4 |
2/1 |
1/1 |
– |
– |
– |
– |
1/1 |
– |
1/1 |
4/1 |
27/9 |
|
Spermophilus sp . |
20/4 |
3/1 |
4/1 |
22/6 |
7/3 |
2/1 |
2/1 |
12/4 |
1/1 |
8/2 |
– |
81/24 |
|
Marmota baibacina |
69/8 |
14/3 |
18/2 |
67/4 |
52/6 |
11/1 |
6/1 |
141/4 |
7/1 |
97/8 |
87/10 |
569/48 |
|
Castor fiber |
2/1 |
1/1 |
– |
1/1 |
1/1 |
– |
– |
– |
– |
– |
– |
5/4 |
|
Cricetus sp. |
1/1 |
1/1 |
3/2 |
– |
– |
– |
– |
2/1 |
– |
1/1 |
1/1 |
9/7 |
|
M. myospalax |
29/4 |
9/2 |
5/1 |
6/1 |
1/1 |
1/1 |
– |
2/1 |
4/1 |
6/1 |
8/2 |
71/15 |
|
Arvicola terrestris |
2/2 |
1/1 |
– |
1/1 |
– |
– |
– |
– |
– |
1/1 |
1/1 |
6/6 |
|
Canis lupus |
12/1 |
10/3 |
7/1 |
– |
– |
– |
– |
– |
– |
3/1 |
4/1 |
36/7 |
|
Vulpes vulpes |
17/2 |
3/1 |
2/1 |
4/1 |
4/1 |
– |
1/1 |
4/1 |
– |
4/1 |
3/1 |
42/10 |
|
Ursus arctos |
13/2 |
4/1 |
4/2 |
– |
– |
1/1 |
– |
– |
– |
3/2 |
7/2 |
32/11 |
|
Martes zibellina |
31/5 |
14/3 |
3/1 |
1/1 |
1/1 |
3/1 |
– |
7/1 |
1/1 |
12/2 |
10/2 |
83/18 |
|
Mustela nivalis |
– |
– |
– |
– |
– |
– |
– |
– |
– |
– |
1/1 |
1/1 |
|
Mustela sibirica |
– |
– |
– |
1/1 |
– |
– |
– |
– |
– |
– |
– |
1/1 |
|
Mustela altaica |
– |
– |
1/1 |
1/1 |
1/1 |
– |
– |
1/1 |
1/1 |
1/1 |
– |
6/6 |
|
Meles anakuma |
– |
– |
– |
– |
– |
– |
– |
1/1 |
– |
1/1 |
– |
2/2 |
|
L. lutra |
– |
– |
– |
– |
– |
– |
– |
– |
– |
2/2 |
– |
2/2 |
|
L. lynx |
7/1 |
– |
5/2 |
– |
– |
– |
– |
– |
– |
1/1 |
– |
13/4 |
|
Moschus moschiferus |
– |
1/1 |
– |
– |
– |
– |
– |
– |
– |
2/1 |
– |
3/2 |
|
Cervus elaphus |
1117/39 |
453/20 |
281/24 |
117/3 |
42/2 |
21/2 |
3/1 |
35/2 |
6/2 |
72/2 |
180/7 |
2327/102 |
|
Alces alces |
6/1 |
3/1 |
2/1 |
– |
– |
– |
– |
– |
– |
– |
– |
11/3 |
|
Caprolus pygargus |
709/23 |
280/18 |
193/11 |
289/2 |
142/8 |
47/4 |
16/1 |
18/7 |
27/2 |
250/4 |
218/7 |
2189/86 |
|
Bos-Bison sp. |
– |
– |
– |
– |
– |
– |
– |
1/1 |
– |
– |
– |
1/1 |
|
Capra sibirica |
7/1 |
5/1 |
3/1 |
– |
2/1 |
2/1 |
– |
6/1 |
– |
5/1 |
– |
30/7 |
|
Ovis ammon |
– |
– |
1/1 |
– |
1/1 |
– |
– |
1/1 |
– |
– |
– |
3/3 |
|
Pisces |
4 |
– |
– |
3 |
1 |
1 |
3 |
– |
3 |
1 |
– |
16 |
|
Amphibia |
– |
– |
– |
– |
– |
– |
– |
1 |
– |
– |
– |
1 |
|
Aves |
67 |
9 |
18 |
4 |
5 |
9 |
– |
– |
4 |
17 |
21 |
154 |
|
Всего костных остатков |
2712 |
999 |
765 |
603 |
301 |
104 |
39 |
247 |
59 |
514 |
673 |
7018 |
*число костей / минимальное число особей
Таблица 2. Распределение фрагментов костей фоновых видов крупных млекопитающих в голоценовых отложениях Каминной пещеры
|
Элементы скелета |
Марал |
Косуля |
Лошадь |
Корова |
Овцы-козы |
|||||
|
экз. |
% |
экз. |
% |
экз. |
% |
экз. |
% |
экз. |
% |
|
|
1, 2 шейные позвонки, обломки таза |
39 |
1,64 |
38 |
1,63 |
12 |
2,65 |
1 |
0,78 |
10 |
1,83 |
|
Фрагменты черепа, рогов, верхней и нижней челюсти |
272 |
11,46 |
481 |
20,64 |
17 |
3,76 |
15 |
11,72 |
68 |
12,45 |
|
Изолированные зубы |
285 |
12,01 |
498 |
21,37 |
245 |
54,20 |
24 |
18,75 |
152 |
27,84 |
|
Обломки лопатки, крупных трубчатых костей |
454 |
19,12 |
348 |
14,94 |
37 |
8,18 |
23 |
17,97 |
135 |
24,73 |
|
Обломки метаподий, кости запястья и заплюсны |
1324 |
55,77 |
965 |
41,42 |
144 |
31,86 |
65 |
50,78 |
181 |
33,15 |
Каминная, 6 052 кости
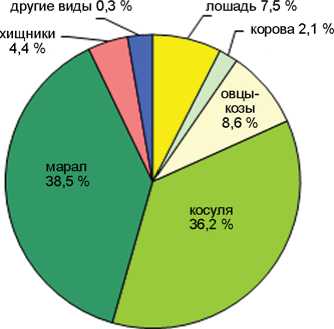
Кучерла, 9 862 костей
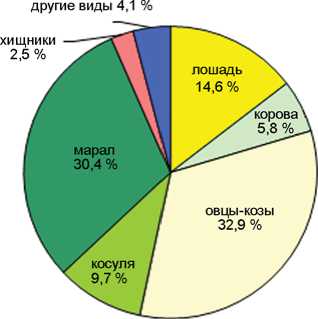
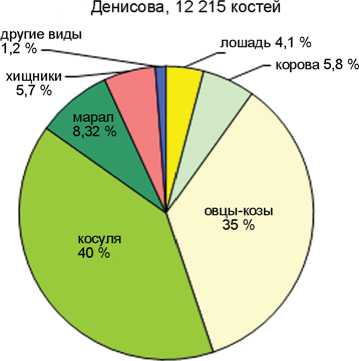
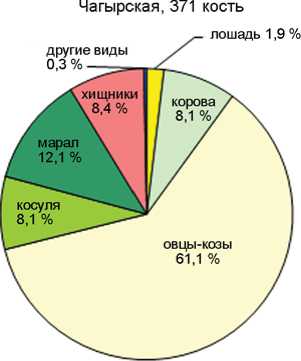
Соотношение домашних и охотничье-промысловых видов в голоценовых слоях пещер Каминная, Кучерла (по: [Молодин, Васильев, Оводов, 2008]), Денисова (по: [Васильев, Гребнев, 1994; Васильев, Шуньков, 2010; Васильев и др., 2019]) и Ча-гырская (по: [Деревянко и др., 2018]).
Наибольший интерес представляют костные остатки следующих видов.
Бобр ( Castor fiber ). В слое 1 найден неполный диафиз бедренной кости, в слое 2 – дистальная половина плечевой кости. В слоях 4 и 5 – обломки резцов.
Остатки волка (Canis lupus) обнаружены в 1–3 и 11 слоях. Преобладают фрагменты крупных костей посткраниального скелета (33 %), изолированные зубы (28 %), обломки верхней и нижней челюсти (22 %). Кости дистальных отделов конечностей немногочисленны (13 %). Три неполные ветви нижней челюсти дают некоторое представление о размерах голоценовых волков Алтая. Так, длина челюсти составляет ок. 181 и 183 мм (163,3-М 180,0–195 мм, n = 21 у современных Canis lupus юга Западной Сибири). Длина Р1-М3 – 100 и 93,6 мм (84,8-М 94,4–102 мм, n = 19). Длина ряда М – 46, 46,3 и 48 мм (42-М 45,6–50,4 мм, n = 19). Высота посредине М1 – 33, 31,4 и 30 мм (27,5М 31,3–37,9 мм, n = 21). Высота за М3 – 33,5, 33,4 и 34,4 мм (29,8 М 34,2–40 мм, n = 21). Таким образом, голоценовые волки из Каминной по размерам нижней челюсти близки к средним значениям соответствующих промеров современных Canis lupus.
Кости бурого медведя (Ursus arctos) присутствуют в 5 из 10 слоев (см. табл. 1), составляя в среднем 0,5 % от числа всего определимого материала. Наиболее представлены в слоях целые и фрагментарные ме-таподии, фаланги, кости запястья и заплюсны (56 %) и изолированные зубы (22 %). Ширина 2 астрагалов в проекции составляет 44 и 49 мм при поперечнике 37,8 и 45,2 мм. Те же промеры у голоценового медведя из пещеры Памятная в Хакасии [Васильев, Гребнев, 2009] – 49-М 55,13–61,6 мм и 42,6-М 47,98–52,5 мм, n = 21–22). Длина Мс I из Каминной – 78 мм (73,8-М 83,55–89,7 мм, n = 19); Мс IV – 86 мм (80-М 95,18– 106 мм, n = 29); Мт III – 76 мм (75-М 85,48–95 мм, n = 23). Ширина дистального отдела плечевой кости – 88 мм (87,3-М 102,82–121,5 мм, n = 27), то же лучевой кости – 61 мм (50,2-М 61,37–70,4 мм, n = 28 у медведей из пещеры Памятной). Измерения показывают, что большинство о статков медведей из Каминной принадлежало взрослым некрупным самкам, чьи про- меры находятся на уровне либо даже несколько ниже минимальных значений соответствующих промеров голоценовых медведей Хакасии.
Соболь ( Martes zibellina ). Среди хищных его остатки наиболее многочисленны (1,2 % от числа всех определимых ко стей) и присутствуют практически во всех слоях (см. табл. 1). Удельный вес остатков соболя наиболее высок в слоях 1, 2, 9 и 11. Ряд костей посткраниального скелета сохранился целиком. Длина 2 бедренных костей составляет 76,6 и 79 мм, 2 лучевых - 55,2 и 56,1 мм, локтевой - 56,3 мм. По этим и целому ряду других промеров соболь из Каминной укладывается в пределы изменчивости голоценовых соболей из пещер окрестностей Красноярска [Оводов, 2009]. В Каминной особенно хорошо представлены фрагменты нижней челюсти. Длина ряда Р 1 -М2 составляет 28-М 29,50-31 мм, n = 9 (25,1-М 28,4033 мм, n = 129 у соболей из Красноярска). Длина ряда М 1 -М2 - 12,1-М 12,93-13,3 мм, n = 10 (10,8-М 12,3314,9 мм, n = 130); высота под М 1 - 8-М 9,34-10,3 мм, n = 12 (6,8-М 8,28-10,5 мм, n = 131 [Там же]). Таким образом, по размерам тела голоценовый Martes zibellina Алтая в среднем несколько превосходил одновременных ему соболей из района Красноярска.
Барсук ( Meles anakuma ). Представлен ветвью нижней челюсти с обломанной восходящей ветвью (сл. 9) и большеберцовой костью с обломанным проксимальным концом (сл. 11).
Выдра ( L. lutra ). В 11 слое найдены дистальная половина и дистальный конец малой берцовой кости, правая и левая, по размерам принадлежащие к разным особям (ширина нижнего конца 11,6 и 14 мм).
Рысь ( L. lynx ). В слое 1 обнаружены кости скелета молодой особи, включающие: нижний отдел лопатки, часть диафиза плечевой кости, 2 лучевые кости (с отпавшим нижним эпифизом и проксимальная половина), 2 локтевые кости (с обломанным верхним концом и дистальная половина), часть тазовой кости. В 3 слое найдены две неполные ветви нижней челюсти: от старой особи, с сильно стертым клыком и неполным Р4 и от юной особи (М1 еще не прорезался), а также проксимальная треть диафиза большеберцовой кости молодой особи и пяточная кость взрослой особи с обломанным дистальным отделом. Единственная находка рыси в слое 11 - изолированный Р4 (длина/ширина - 18,9/8,8 мм).
Немногочисленные остатки свиньи ( Sus scrofa ) отмечены с перерывами с 1 по 11 слой (см. табл. 1). Дикая и одомашненная формы хорошо различаются по строению черепа. По посткраниальному скелету их различение возможно только на основе размеров костей. С большой долей уверенности к кабану можно отнести имеющие крупные размеры остатки из слоя 5, включающие фрагменты верхней и нижней челюсти, изолированные зубы. В эту же категорию попадают два обломка лопаток из слоя 1 с наименьшей шириной шейки 34,7 и 35,3 мм и пяточная кость длиной
91,7 мм. Пограничное по размерам значение имеет астрагал из слоя 1 с длиной 49,3 мм. По данным В.И. Цалкина [1956], максимальная длина астрагала домашней свиньи Древней Руси достигала 48 мм. Около половины костей свиньи из Каминной принадлежит к молодым особям, что дополнительно осложняет их отнесение к дикой или домашним формам. Домашняя свинья, очевидно, попала на Алтай лишь относительно недавно, с приходом русских поселенцев. В кочевом или полукочевом хозяйстве аборигенного населения этот вид домашних животных трудно себе представить. Скорее всего, все фрагментарные остатки, особенно из 5, 9 и 11 слоев, могут быть отнесены к дикой форме. Остатки Sus scrofa в сравнительно небольшом количестве присутствуют в голоценовых слоях Денисовой пещеры [Васильев, Гребнев, 1994; Васильев, Шуньков, 2010; Васильев и др., 2019] и ритуального грота Кучерла [Молодин, Васильев, Оводов, 2008].
Остатки кабарги ( Moschus moschiferus ) единичны. В слое 2 найден обломок нижней челюсти, в котором сохранились Р3-М 1 . В слое 11 - межчелюстная ко сть и изолированный резец. Ко сти кабарги отсутствуют в голоценовых отложениях Денисовой пещеры [Васильев, Гребнев, 1994; Васильев, Шунь-ков, 2010; Васильев и др., 2019], но отмечены во всех слоях ритуального грота Кучерла-1 в Центральном Алтае, где они составляют до 2 % от числа остатков зверей охотничье-промысловой группы [Молодин, Васильев, Оводов, 2008]. Обилие остатков кабарги служит хорошим индикатором горно-таежных ландшафтов.
Сибирский благородный олень ( Cervus elaphus sibiricus ). Наряду с косулей марал - наиболее массово представленный вид охотничье-промысловой териофауны. Число его остатков достигает 39 % от всех крупных млекопитающих, диких и домашних. Серии из десятков одноименных костей дают по ряду промеров исчерпывающее представление о морфометрических особенностях голоценового марала. К сожалению, не сохранилось ни одной целой крупной трубчатой кости. Лишь для одной из пястных костей, расколотых вдоль, удало сь измерить наибольшую длину. Для сравнения были привлечены материалы по современному парковому маралу, собранные автором ( С.В. ). в конце 1980-х гг. (табл. 3, 4). Если принять сумму средних значений сопоставимых промеров марала из Каминной за 100 %, то современный парковый марал в сравнении с ним составляет лишь 94,3 %. Столь же представительный материал по голоценовому маралу из грота Кучерла [Там же] показывает, что по размерам тела Cervus elaphus sibiricus Северо-Западного и Центрального Алтая не различались (100,2 % у марала Кучерлы). Очевидно, что для полноценного сравнения с плейстоценовыми формами Cervus elaphus следует привлекать в первую очередь серии костей голоценового благородного оленя,
Таблица 3. Размеры костей передней конечности Cervus elaphus sibiricus
Единичные остатки лося (A. alces) происходят из слоев 1–3. В слое 1 это фрагмент нижней челюсти с рядом М (длина 96,5 мм), запястная добавочная ко сть, проксимальный конец пястной кости (шири-на/поперечник – 60,5/41,5 мм), 2 дистальных конца первых фаланг, неполная третья фаланга. Слой 2:
Таблица 4. Размеры костей задней конечности Cervus elaphus sibiricus
Тур или бизон (Bos-Bison sp.). В 9 слое найдена нижняя половина расколотого вдоль симфизного отдела нижней челюсти общей длиной 90 мм. По размерам и морфологии она не отличается от челюсти крупного бизона, и, как и другие костные остатки из 9 слоя, имеет типичную голоценовую сохранность. Скорее всего, фрагмент челюсти принадлежит туру (Bos primigenius). По всем имеющимся на сегодня данным, на юге Западной Сибири бизон исчезает с завершением плейстоцена, и уже в раннем голоцене он замещается здесь другим крупным быком – туром [Пластеева и др., 2020]. Фрагментарные остатки очень крупного быка, не уступающего по размерам первобытному бизону, единично зафиксированы также в голоценовых слоях пещер Денисова и Чагырская [Васильев, Шуньков, 2010; Деревянко и др., 2018].
Ко сти сибирского горного козла ( Capra sibirica ) единично присутствуют с 1 по 11 слой (см. табл. 1). Ширина дистального конца лопатки – 46,8 мм, ширина / поперечник суставной поверхности – 39,0/31,4 мм. Ширина нижнего конца 3 экз. плечевой кости – 45, 42,8, 47,8 мм, ширина суставного блока – 42, 42, 47,7 мм, поперечник минимальный в желобе – 19,8, 19,4, 22,5 мм, высота медиального мыщелка – 26, 25,6, 29,6 мм. Ширина / поперечник 2 диафизов лучевой кости – 27,5/15,5 и 25,3/14,3 мм. Длина пяточной кости – 88 мм, ширина / поперечник дистального конца – 28,5/31 мм, то же пяточного бугра – 19,0/22,7 мм, ширина тела ко сти минимальная – 12,8 мм. Длина астрагала: латеральная – 38,6 мм, сагиттальная – 30 мм, медиальная – 36,2 мм, ширина дистального конца – 24,5 мм, медиальный поперечник – 21,2 мм. Ширина дистального конца пястной кости – 35,5 мм. Ширина / поперечник проксимального конца плюсневой кости – 27,5/26,8 мм. В голоцене Северо-Западного Алтая сибирский козерог был относительно малочисленным видом и окончательно исчез здесь под воздействием человека, очевидно, уже ближе к историческому времени. В Центральном Алтае, в районе грота Кучерла, Capra sibirica был представлен гораздо лучше, его остатки составляют почти 5 % от числа охотничье-промысловой териофауны [Молодин, Васильев, Оводов, 2008].
Неожиданно среди материала обнаружились остатки архара ( Ovis ammon ) типичной голоценовой сохранности. В слое 3 это почти целая коленная чашка (длина – 45, ширина – 35,5, высота – 28 мм). В слое 5 найден неполный первый шейный позвонок (ширина передней и задней суставных поверхностей – 81 и 72 мм соответственно), в слое 9 – третья фаланга. В настоящее время архар, как и сибирский горный козел, в окрестностях пещеры давно истреблен человеком; небольшая популяция сохранилась на Алтае лишь в самых труднодоступных районах на границе с Монголией.
Список литературы Териофауна из голоценовых отложений пещеры Каминная (Северо-Западный Алтай)
- Васильев С.К., Гребнев И.Е. Фауна млекопитающих голоцена Денисовой пещеры // Деревянко А.П., Молодин В.И., Денисова пещера. Часть I. - Новосибирск: ВО Наука, 1994. - С. 167-180.
- Васильев С.К., Гребнев И.Е. Морфология костей скелета голоценового бурого медведя (Ursus arctos L., 1758) Кузнецкого Алатау // Енисейская провинция. Альманах. Вып. 4. - Красноярск: КККМ, 2009. - С. 68-76.
- Васильев С.К., Деревянко А.П., Маркин С.В. Фауна крупных млекопитающих финала сартанского времени Северо-Западного Алтая (по материалам пещеры Каминной) // Археология, этнография и антропология Евразии. - Новосибирск: Изд-во ИАЭТ СОРАН, 2006. - № 2 (26). - С. 2-22. EDN: KJVOHX
- Васильев С.К., Козликин М.Б., Колясникова А.С., Шуньков М.В. Костные остатки из голоценовых отложений в южной галерее Денисовой пещеры (материалы 2017 и 2018 годов) // Проблемы археологии, этнографии, антропологии Сибири и сопредельных территорий. - Новосибирск: Изд-во ИАЭТ СО РАН, 2019. - Т. XXIV. -С. 53-58. EDN: NPLQJZ
- Васильев С.К., Шуньков М.В. Остатки млекопитающих из голоценовых отложений в восточной и южной галереях Денисовой пещеры // Проблемы археологии, этнографии, антропологии Сибири и сопредельных территорий. - Новосибирск: Изд-во ИАЭТ СО РАН, 2010. - Т. XVI. -С. 21-27. EDN: OXDXAB
