The morphology of permanent molars from the Paleolithic layers of Denisova cave

Author: Zubova A.V., Chikisheva T.A., Shunkov M.V.
Journal: Archaeology, Ethnology & Anthropology of Eurasia @journal-aeae-en
Section: Anthropology and paleogenetics
Article in issue: 1 т.45, 2017.
Free access
Short address: https://sciup.org/145145290
IDR: 145145290 | DOI: 10.17746/1563-0110.2017.45.1.121-134
Text of the review article The morphology of permanent molars from the Paleolithic layers of Denisova cave
The archaeological site of Denisova Cave is located in the south of the West Siberian region, in the Altai Mountains, close to the Russian Federation’s border with China, Mongolia, and Kazakhstan.
The remains of Pleistocene hominins from the Denisova Cave have been studied for more than 30 years. First dental remains were excavated here in 1984: a deciduous molar named Denisova 2 was found in the stratigraphic layer 21.1 (Shpakova, Derevianko, 2000). In the same year, one more tooth was found in layer 12. It was initially attributed as an upper permanent incisor (Turner, 1990; Shpakova, Derevianko, 2000); but later, a comparative study demonstrated that the tooth actually belonged to an animal from the Bovidae family (Viola et al., 2011: 209). In 2000, a well-preserved upper permanent molar was recovered from layer 11.1 (Denisova 4); and in 2010, fragments of the crown of an upper permanent molar (Denisova 8) were found in the bottom of stratigraphic layer 11.4, in the contact zone with layer 12.
According to geochronological data, the Denisova 4 and Denisova 8 permanent molars are dated to 50–40 ka BP; however the layer containing the Denisova 4 is younger than the layer where Denisova 8 was found (Sawyer et al., 2015).
Sequencing of mtDNA and the nuclear genome from the distal phalanx of the carpal minimus of a 6–7 year old girl (which was found in layer 11.2 (Denisova 3)) and the Denisova 4 molar has shown that both specimens represent a formerly unknown hominin species. The species was named “Denisovan” (Reich
et al., 2010), or Homo altaiensis (Derevianko, 2011) by the authors of the study. It was the first case in the history of physical anthropology when a new taxon was described by the results of genetic analysis, rather than by a morphological study.
The analysis of the mtDNA has shown that the hominin lineages ancestral to Denisovans, Neanderthals, and Homo sapiens diverged about a million years ago (Krause et al., 2010). The study of the nuclear DNA revealed a much later date for the separation of the line leading to H. sapiens from the hominin population ancestral to H. neanderthalensis и H. altaiensis , about 800 ka BP (Meyer et al., 2012). The divergence between Denisovan and Neanderthal lineages was initially thought to have occurred 640 ka BP (Reich et al., 2010); but according to the results of recent studies, it happened only some 430 ka BP (Meyer et al., 2016).
Denisovans have occupied the Altai Mountains for a long period of time. Judging by the rate of accumulation of mutations in the mtDNA of present-day humans, it can be hypothesized that the Denisova 2 and Denisova 8 specimens are some 65 thousand years more ancient than the Denisova 3 and Denisova 4 samples (Slon et al., 2015). It was found that Denisovan genome contained alleles associated with a dark skin-color, chestnut hair, and brown eyes (Meyer et al., 2012). According to the results of analysis of the mtDNA of all individuals found at Denisova Cave, the level of the intragroup genetic diversity in the Denisovan population was much lower than in both Neanderthals and modern humans (Meyer et al., 2012; Slon et al., 2015).
The place of origin of the Denisovans, as well as their position in the hominin taxonomy, have been hotly debated. On the basis of archaeological data, A.P. Derevianko put forward a hypothesis that the Denisovans are a subspecies of polymorphic H. sapiens, contemporaneous with other subspecies, e.g. H. sapiens neanderthalensis , H. sapiens africaniensis , and H. sapiens orientalensis (Derevianko, 2011). According to this hypothesis, the subspecies originated from local Asian populations of H. erectus (Ibid.). However, recent paleogenetic data on the Pleistocene hominins from Sima de los Huesos in Spain suggest that the mitochondrial genomes of this population and those of Denisovans were similar (Meyer et al., 2016). This result questions an Asian origin for the most ancient components in the genome of H. altaiensis .
This study presents the results of an extended morphological analysis of the Denisova 4 and Denisova 8 permanent molars. The results describe peculiarities of H. altaiensis dentition, and infer relationships of the Denisovan population with other taxa: H. erectus s.l. , H. heidelbergensis s.l. , H. neanderthalensis , and H. sapiens s.l.
Study protocol
The dental samples from Denisova Cave were previously described using standard ASUDAS protocol and the protocol of Neanderthal apomorphies, during the paleogenetic study of the Denisovan hominins (Krause et al., 2010; Sawyer et al., 2015).
We carried out an examination of the Denisova 4 and Denisova 8 upper molars employing a substantially extended set of traits. The set was generally based on the ASUDAS protocol (Turner, Nichol, Scott, 1991; Scott, Turner, 1997) and the conventional set of traits used in Russian dental anthropology (Zubov protocol), which includes a comprehensive description of the pattern of crown grooves (Zubov, 1968, 1974, 2006; Zubov, Khaldeyeva, 1989, 1993). The protocol of the Neanderthal complex markers (Bailey, 2002; Bailey, Skinner, Hublin, 2011) was also employed in our study, as well as a protocol previously developed for accounting of plesiomorphic traits in modern human populations (Zubova, 2013).
Each of the protocols has some advantages, and thus the combination of these protocols can help to extract as much information as possible from the dental samples. The ASUDAS protocol employs the most diversified scales for fixing dental traits that are particularly precise in terms of describing the dental patterns found in modern populations. The main advantage of the traits put forward by S. Bailey is their increased sensitivity to the presence of Neanderthal genes in a population. The Zubov protocol permits not only study of the details of the macrorelief of the crown, but also the pattern of its grooves, which represent an independent and hierarchically organized set of traits: odontogliphics.
In total, 60 dental traits were scored. In the molars studied, the following traits were observed: reduction of the hypocone and metacone, rhomboid shape of the upper molars, metaconulus, mesiostylid, enamel extension, Carabelli cusp, distal and mesial accessory cusps of the upper molars, epicrista, plagiocrista (crista oblique), entocrista, posterior fovea, and odontoglyphic traits.
Hypocone reduction. The trait describes the dynamic of the hypocone’s size relative to the protocone and metacone. The ASUDAS and Zubov protocol scales have some differences: the former includes 6 grades (0 – absence of the cusp, 5 – maximal development of the cusp) (Turner, Nichol, Scott, 1991: 18), while the latter has only 4 grades (4, 4–, 3+, 3), where grade 4 stands for the maximal development of the cusp, and grade 3 for the absence of the cusp (Zubov, 1968: 152).
Metacone reduction. This trait describes the dynamic of size of the metacone relative to the paracone. In ASUDAS, the trait is scored in the same way as the hypocone reduction (Turner, Nichol, Scott, 1991: 18), while the Zubov protocol employs a 5-level scale (1 to 5), where grade 1 means the absence of reduction, and grade 5 its maximal development (Zubov, 1968: 160).
Rhomboid shape of the upper molars. This trait describes upper permanent molars showing a nonreduced hypocone, strongly developed in the disto-lingual direction, and a reduced metacone, which forms an oblique disto-vestibular corner of the crown (Bailey, 2004).
Metaconulus . A small cusp in the depth of the enamel of the axial ridge of the metacone, in its central part (Zubov, Khaldeyeva, 1993: 68).
Carabelli cusp . A styloid cusp in the mesio-lingual part of the crown. Grades 0 to 7 in ASUDAS: 0 – absence, 7 – maximal development (Turner, Nichol, Scott, 1991: 19). Zubov protocol employs 5 grades from 0 to 5 (1968: 157). The grades of the two protocols correspond as follows:
|
ASUDAS |
Zubov |
|
0 |
0 |
|
1,2 |
1 |
|
3,4 |
2 |
|
5 |
3 |
|
6 |
4 |
|
7 |
5 |
Distal accessory cusp of the upper molars. A styloid cusp at the distal marginal ridge, at the interface between the metacone and hypocone. A scale, in which grade 1 correspond to the absence of the cusp and grade 6 to maximal development, is employed in both protocols (Zubov, 2006: 56; Turner, Nichol, Scott, 1991: 19).
Mesial accessory cusp of the upper molars. A cusp at the mesial margin of the crown. It is delimited by an additional groove that falls into the fissure separating the paracone and protocone. Phylogenetically, it differs from the cusps formed by the distal segments of the paracone and protocone (Zubov, Khaldeyeva, 1993: 67; Scott, Turner, 1997: 45). A scale for evaluating the degree of development of this trait is absent.
Epicrista . A ridge connecting protocone and metacone. The trait is present when the fissure dividing the cusps is fully covered.
Plagiocrista . A ridge connecting the metacone and protocone. This study employs a 4-level scale for this trait (Zubova, 2013: 114).
Entocrista . A marginal ridge connecting the protocone and hypocone (Zubov, Khaldeyeva, 1989: 62). It is almost never found in modern humans.
Posterior fovea. An elongated depression in the distal part of the upper and lower molars, which can vary in length. In modern humans, it is typically located at the occlusal surface of the metacone. Mesially, it is delimited by the distal segment of the metacone, and distally by the distal marginal ridge (Zubov, 2006: 61). In ancient specimens, it can reach the surface of the hypocone as well. In such cases, it is delimited by the same elements as in the metacone. Scales for evaluating the degree of development of this trait are absent.
Odontoglyphic elements . Like other dental traits, these elements are not equal in terms of taxonomic value. Combinations of the elements reflect the evolutionary status of the population (Zubov, 1974). The nomenclature of the odontoglyphic elements has been repeatedly changing during its development. As a result, different researchers employ rather different nomenclatures for these elements (Zubov, 1974, 2006; Zubov, Khaldeyeva, 1989; Hillson, 1996). In our study, we follow the methodology described in the last publications of A.A. Zubov (Zubov, Khaldeyeva, 1989; Zubov, 2006). In the next section, the classification and description of the odontoglyphic traits are outlined.
We employ three types of the grooves of the occlusal surface of the crown. The first and most ancient type comprises the intertubercular fissures of the first order. They separate major cusps of the crown, and are designated as I–IV for the upper molars, and I–VI for the lower molars. Fissure I separates the metacone and the paracone; fissure II separates the paracone and the protocone; fissure III separates the metacone and the protocone; and fissure IV separates the hypocone from both the metacone and the protocone.
Tubercular grooves of the second order delimit the axial ridges of each cusp, dividing a cusp into three segments. In some studies, these segments are designated as mesial, central, and distal (Carlsen, 1987; Bailey, Skinner, Hublin, 2011). Intersegmental grooves are tagged as 1 and 2 plus the two (or three for the lower) first characters of the name of the cusp (e.g. “pa” stands for the paracone, “prd” for the protoconid) (Zubov, 1974).
In the metacone and paracone, these grooves separate the axial segment from mesial one (groove 1), and the axial segment from the distal one (groove 2). The grooves are designated as 1me, 2me, 1pa, and 2pa, respectively. In the protocone and hypocone, groove 1 separates the axial and distal segments (1pr, 1hy), groove 2 the axial and mesial segments (2pr, 2hy).
Grooves of the third order are divided into two categories. The first includes accessory grooves dividing the mesial and distal segments of each cusp into two parts in the sagittal direction, and lying parallel to grooves 1 and 2. These are designated as 1’ and 2’, respectively. The second category includes accessory grooves of the axial ridges of the major cusps, designated as 3 and 4 (Zubov, Khaldeyeva, 1989; Zubov, 2006).
In the metacone, 1’me falls into fissure I, and divides the mesial segment in the vestibular direction, while 2’me falls into fissure III parallel to the posterior fovea and divides the distal segment in vestibular direction. In the paracone, 1’pa falls into fissure I and divides the distal segment in vestibular direction, 2’pa falls into fissure II and divides the mesial segment in vestibular direction, 3pa falls into fissure II and divides the axial segment in vestibular direction, and 4pa divides the axial segment in medio-distal direction. In the protocone, 1’pr falls into fissure IV and divides the distal segment in lingual direction, 2’pr falls into fissure II and divides the mesial segment in lingual direction, 2’’pr duplicates 2’pr in the mesial segment and falls into fissure II or 2’pr, 3pr falls into fissure II and the central fovea and divides the axial segment in lingual direction, 4pr bisects the axial segment in transverse direction. In the hypocone, 1’hy falls into fissure IV or the posterior fovea and divides the distal segment in lingual direction, 2’hy falls into fissure IV or the basin of the talon and divides the mesial segment in lingual direction.
The total number of tubercular grooves and their directions differ in various species of the genus Homo . The grooves of the third order are the most variable. Many of them are reduced, and very rare in modern humans. An increased irregularity of enamel is typical of early Homo , and teeth of these hominins can exhibit additional grooves of the third order, which are absent in later species of Homo . The positions of the points of contact between tubercular grooves and the inter-cusp fissures can also vary. Furthermore, the tendency to ridge formation is more prominent in extinct representatives of the genus Homo than in modern humans.
The traits studied can be divided into three groups. The first group includes evolutionary stable and taxonomically neutral variables related to the basal teeth morphology in the hominin lineage and its evolutionary continuity (e.g. four-cusped upper molars, five-cusped lower molars or the number of intertubercular grooves of the crown). Traits of this group are present in all Homo species.
The second group is composed of so-called generalized archaic markers. These are plesiomorphic traits displaying a negative temporal dynamic. In other words, they are found with the highest frequency in early
Homo,
and become increasingly rare in later
Homo
species. The second group includes such traits as derivatives of the cingulum; ridge-forming structures of molars; M1
The third group includes traits that are evolutionarily progressive among members of the genus Homo, and is composed of two blocks of traits. One of the blocks includes traits describing reduction of the dentition: hypocone and metacone reduction, decrease in the size of distal teeth in a row, small size or absence of the styloid cusps in the distal part of the lower molars, axial position of the hypoconulid, simplified odontoglyphic pattern, etc. The frequency of these traits steadily increases with time in the hominin lineage. Another block includes apomorphic patterns of various taxa.
When describing the upper molars from Denisova Cave, most attention was paid to the traits of the second and third groups, namely plesiomorphic traits and the markers of Neanderthal and modern dental patterns. The frequency traits were then compared with the patterns typical of H. erectus s.l. , H. heidelbergensis s.l. , H. neanderthalensis , and H. sapiens s.l.
Previously published raw data, as well as high-definition images and morphological descriptions of finds, were used as reference data (see Table).
Material
Our sample comprises two permanent molars. The Denisova 4 molar from the lithological layer 11.1 was previously described by B. Viola as an upper left third molar (Viola et al., 2011). But the pattern of attritional facets provides some evidence that the tooth might also be a second molar, if the individual lacked the third molar. In the study cited, the patterns of enamel macrorelief and dental metrics were published. The authors pointed out a similarity in dental metrics between the Denisovan specimen and early Homo (and even australopithecine) samples, but not Neanderthal dentition.
The Denisova 8 molar also belongs to the left side of the upper jaw. A.P. Buzhilova (2014) determined it to be an upper second or third permanent molar, while Viola pointed that this was most probably the third upper left molar (Sawyer et al., 2015). This tooth is worse preserved than the Denisova 4 molar: the roots were completely lost, the crown was reconstructed from fragments, and the mesial part of the crown at the interface between the paracone and protocone was destroyed. The macrorelief of the cusps of the trigon was almost completely worn off, as well as most intertubercular fissures. But there is almost no attrition at the hypocone and the distal part of the metacone; just one small contact-facet can be seen on the top of the main ridge of the hypocone, in its mesial part. On the interproximal surface of the distal part there is no contact-facet.
Morphological description
Left upper permanent second or third molar (М2/3) from the layer 11.1, Denisova 4 (Fig. 1). This belonged to a young adult male (Slon et al., 2015).
The tooth is very massive, with a long neck. The roots are strongly divergent in the mesial and distal norms, the crown exhibits rounded corners. The bucco-lingual
Materials used for comparison
|
Region |
Taxonomic status of the find * |
Site |
Specimen No. |
Source |
|
1 |
2 |
3 |
4 |
5 |
|
Africa |
Anatomically modern Homo sapiens |
Fish Hoek |
– |
Schwartz, Tattersall, 2003 |
|
Homo helmei |
Florisbad |
– |
Ibid. |
|
|
Homo habilis |
Hadar |
A.L. 666 |
" |
|
|
Homo heidelbergensis |
Kabwe |
– |
" |
|
|
Homo erectus |
Koobi Fora |
KNM-ER 1813, KNM-ER 3733 |
" |
|
|
Homo ergaster |
Nariokotome |
KNM-WT 15000 |
Khaldeyeva, Zubov, Kharlamova, 2012 |
|
|
Homo habilis |
Olduvai Gorge |
OH 6, OH 13, OH 16, OH 24 |
Schwartz, Tattersall, 2003, vol. II |
|
|
Homo erectus ? Athlanthropus mauritanicus (?) |
Thomas Quarry (Rabat) |
– |
Ibid. |
|
|
Homo erectus |
Konso |
KGA 4-14, KGA 11-350 |
Suwa et al., 2007 |
|
|
Early Homo sapiens |
Dar es-Soltan |
DS II – H9, DS II – NN, DS II – H5, DS II – H10 |
Hublin et al., 2012 |
|
|
" |
Smugglers’ Cave |
Ctb H7, Ctb Ib 19, Ctb T4, Ctb T3b |
Ibid. |
|
|
West Asia |
Early Homo sapiens |
Jebel Qafzeh |
Qafzeh 4, Qafzeh 5, Qafzeh 6, Qafzeh 9, Qafzeh 11 |
Schwartz, Tattersall, 2003, vol. II |
|
Homo neanderthalensis |
Skhul |
Skhul IV, Skhul V |
Ibid. |
|
|
" |
Tabun |
Tabun I, Tabun T I, Tabun T II |
" |
|
|
" |
Kebara |
KNM 24, KNM 21 |
Tillier et al., 2003 |
|
|
Early Homo sapiens |
Qesem |
– |
Hershkovitz et al., 2011 |
|
|
East and Central |
Archaic Homo sapiens |
Jinniushan |
– |
Schwartz, Tattersall, 2003, vol. II |
|
Asia |
Homo sapiens |
Liujiang |
– |
Ibid. |
|
Homo erectus |
Sangiran |
Sangiran 4, 7, 17, 27, NG 91- G10 No1, NG 0802.1, NG 0802.3, NG 92.3, Njg 2005.05, Bpg 2001.04, PDS0712, NG0802 |
Ibid.; Zanolli, 2013; Kaifu et al., 2007; Zaim et al., 2011 |
|
|
Homo sapiens |
Wadjak |
Wajak 1, Wajak 2 |
Schwartz, Tattersall, 2003, vol. II |
|
|
Homo erectus |
Zhoukoudian, Lower Cave |
ZKD 169.25, ZKD PA 327, ZKD skull XI |
Ibid. |
|
|
Homo sapiens |
Zhoukoudian, Upper Cave |
PA 101, PA 102, PA 103 |
Ibid.; Turner, Manabe, Hawkey, 2000 |
|
|
Early Homo |
Xujiayao |
PA 1480, PA 1481, PA 1500 |
Xing et al., 2015 |
|
|
Homo sapiens |
Daoxian |
DX 1, 4, 5, 6, 8, 12, 14, 16, 17, 20, 21, 24, 28, 31,33, 35, 36, 39, 41, 47 |
Liu et al., 2015 |
|
|
Homo erectus |
Liang Bua |
LB1 |
Kaifu et al., 2015 |
|
|
North Asia |
Homo sapiens |
Malta |
1, 2 |
Zubov, Gokhman, 2003; Zubova, Chikisheva, 2015 |
|
Homo neanderthalensis |
Chagyrskaya |
10, 51.1, 57 |
Unpublished data of A.V. Zubova |
|
|
Western Central Asia |
Homo neanderthalensis |
Obi-Rakhmat |
– |
Glantz et al., 2008; unpublished data of A.V. Zubova |
|
" |
Teshik-Tash |
– |
Unpublished data of A.V. Zubova |
Table (end)
|
1 |
2 |
3 |
4 |
5 |
|
Europe |
Homo sapiens |
Abri Pataud |
Pataud 1 |
Schwartz, Tattersall, 2003, vol. II |
|
Homo heidelbergensis |
Arago |
Arago 21, 14, 31 |
Ibid. |
|
|
Homo antecessor |
Atapuerca: Gran Dolina |
ATD 6-69 |
" |
|
|
Homo heidelbergensis / antecessor |
Atapuerca: Sima de los Huesos |
AT-16, AT, 3177, AT-138, AT-406, AT-139, AT-26, AT-959, AT-20, AT-2076, AT-812, AT-944, AT-196, AT-2071, AT-4317, AT-3424, AT-587, AT-46, AT-4326, AT-960, AT-824, AT-2179, AT-407, AT-4319, AT-4336, AT-12, AT-2175, AT-815, AT-821, AT-15, AT-170, AT-602, AT-816, AT-274, AT-3181, AT-171, AT-826, AT-601, AT-945, AT-1471, AT-2393, AT-3183, AT-194, AT-5082, AT-2150, AT-140 |
Martinón-Torres et al., 2012 |
|
|
Homo erectus |
Dmanisi |
D 2882, D 2700 |
Schwartz, Tattersall, 2003; Martinón-Torres et al., 2008 |
|
|
Homo sapiens |
Engis |
Engis 2 |
Schwartz, Tattersall, 2003, vol. II |
|
|
" |
Grimaldi |
Barma grande 2 |
Ibid. |
|
|
" |
Isturitz |
Ist 71 |
" |
|
|
Homo neanderthalensis |
Krapina |
45, 46, 47, 48, D 119, D 120, D 170, D 180, D 136, D 164, D 178, D 188 |
Radovcic et al., 1988 |
|
|
" |
La Quina |
H 5, H 18 |
Schwartz, Tattersall, 2003, vol. II |
|
|
" |
Le Moustier |
– |
Ibid. |
|
|
Homo sapiens |
Mladec |
1, 2 |
" |
|
|
Homo neanderthalensis |
Pech-de-l’Azé |
– |
" |
|
|
" |
Saccopastore |
2 |
" |
|
|
" |
Sakajia |
– |
" |
|
|
" |
Scladina |
– |
" |
|
|
" |
Spy |
1, 2 |
" |
|
|
" |
Subalyuk |
2 |
" |
|
|
Homo sapiens |
Akhshtyrskaya |
– |
Unpublished data of A.V. Zubova |
|
|
" |
Rozhok-1 |
– |
Same |
|
|
" |
Caldeirao |
1 |
Trinkaus, Bailey, Zilhao, 2001 |
|
|
" |
Sungir |
2, 3 |
Zubov, 2000 |
|
|
" |
Kostenki |
14, 15, 17, 18 |
Khaldeyeva, 2006; unpublished data of A.V. Zubova |
|
|
" |
Visogliano |
6 |
Abbazzi et al., 2000 |
|
|
" |
Galeria da Cisterna |
– |
Trinkaus et al., 2011 |
|
|
Early Homo |
Pe§tera cu Oase |
2 |
Trinkaus, 2010 |
*Since the taxonomic status of some finds is debatable, information in this column is given according to the opinion of the authors referenced in this article.
diameter of the metacone is small relative to the paracone, while the mesio-buccal corner demonstrates only moderate obliquity. As a result, the crown is narrowed in its distal portion. But since the hypocone is not biased lingually, the crown does not display the rhomboid shape typical of the Neanderthal upper molars (Fig. 2). The apexes of the major cusps are inclined towards the center of the crown. The intertubercular fissures are very deep. Fissures I and III are visible only on the occlusal surface, while fissures II and IV expand to the vertical surfaces of the tooth: fissure IV, which divides the hypocone and protocone, reaches the lingual surface, and fissure II, which separates the protocone and paracone, is present on the mesial surface (Fig. 3).
The paracone is divided into three segments by deep grooves, which dissect the margin of the crown and continue in the upper portion of the buccal surface. The mesial ridge of the paracone is wider than its distal ridge, which disappears in the middle of the axial ridge. The groove delimiting the mesial ridge joins the intertubercular fissure, which separates the metacone and paracone. The paracone exhibits deep grooves 1ра and 2ра, the first of which falls into fissure I, and the second into the central fovea. The terminal segments of the grooves lie on the border of the occlusal plane, and change their direction towards the apex of the axial ridge (Fig. 3). So the abovementioned grooves delimit elements of the marginal ridge of the vestibular surface (eocrista). The latter are found in the central branch of the ridge, and are not present in modern humans. At the interface between the paracone and metacone, the ridge is intercepted by fissure I and a short parallel groove 1’pa.
The metacone is not reduced. Its mesio-distal diameter is not less than that of the paracone. On the surface of the metacone, an axial, a mesial, and a distal segment can be distinguished. The axial segment is very massive, and its distal portion is divided into separate fragments by accessory transversal grooves. The distal and mesial segments of the metacone are clearly visible only at the vestibular margin of the crown.
The plagiocrista is almost completely interrupted by the central groove; only a thin enamel bridge in the distal portion, lying parallel to fissure IV, is left.
The terminal segments of the first and second grooves of the metacone continue on the vestibular surface. They form separate apexes of the mesial and distal portions of the cusp. Groove 1me falls into fissure I just below 1pa, and 2me merges with the basin of the talon. Two accessory enamel ridges branch out of the axial ridge of the metacone in the distal direction. These are oriented towards the accessory cusps of the distal margin of the crown, but do not reach it, being intercepted by the basin of the talon. The ridges are separated by an accessory groove of the third order. The groove delimits elements of the metaconulus in the axial ridge, and does not match precisely with conventional elements of the odontogliphic pattern of the molars of modern humans (Zubov, 2006; Zubov, Khaldeyeva, 1989). Following the nomenclature of other cusps, it is referred to as 4me. In the vicinity of the central fovea, the axial segment of the metacone is divided in the axial plane by one more accessory groove of the third order (3me), which originates approximately in the middle third of the ridge and falls into fissure III.
The hypocone is large, sub-oval in shape, and elongated in the vestibular-lingual direction. A massive central ridge is prominent on its surface. The mesial segment of the cusp is very thin, and the groove that delimits it is almost merged with fissure IV. The distal segment is somewhat better pronounced. On the occlusal surface, the hypocone is separated from the metacone and protocone by a wide and elongated basin of the talon, which merges with the elements of the posterior fovea at the metacone. In its distal portion, the hypocone is separated from the metacone by a rounded accessory cusp in the marginal ridge (С5 grade 1 ASUDAS). Both major grooves of the hypocone (1hy and 2hy) fall into the basin of the talon. These are not particularly long, but rather deep. They are duplicated in the mesial and distal segments by parallel grooves of the third order, which are very rarely found in modern humans. Similarly to corresponding grooves of other cusps, they can be referred to as 1’hy and 2’hy. An element of the marginal ridge is prominent in the apex of the cusp, as is the case in the metacone and paracone.
The protocone is massive: it is the largest cusp of the crown. Such a large size is related to an increase in size of its basal portion owing to the Carabelli cusp, which contacts with the mesio-lingual groove separating the hypocone and protocone. The Carabelli cusp occupies the whole base of the protocone in its cervical portion (Fig. 1). In the middle third of the height of this cusp, there are four apexes formed by cingular ridges (Fig. 2). Thus, the genetic potential of the upper molar growth is more fully realized in this specimen than in modern human teeth. Multiple apexes in such cases are a manifestation of rudimentary derivatives of the cingulum, which are referred to by P. Hershkovitz (1971) as entostyles.
Finding a correspondence between the morphological pattern described above on the one hand, and the grades of standard dental protocols on the other, is a complicated task, since none of the protocols has grades to describe a cusp with multiple apexes. In ASUDAS, the pattern, in which the Carabelli cusp contacts with the intertubercular groove, is referred to as grade 5; in Zubov protocol it is grade 4.
The protocone, like other major cusps, is segmented into three portions. The central segment is the largest, followed by the mesial, and the distal segment is the smallest. The second groove of the protocone (2pr)
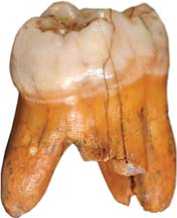
Fig. 1. Denisova 4: disto-vestibular view.

Fig. 2. Denisova 4: occlusal view.
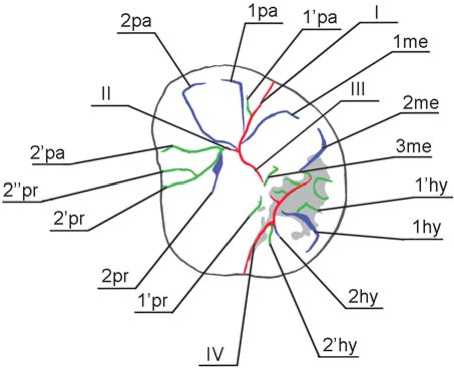
Fig. 3. Denisova 4: odontoglyphic pattern.
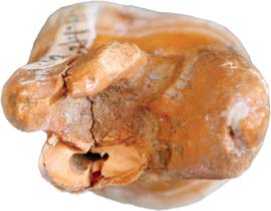
Fig. 4. Denisova 4: root system.
merges with 2’pr and forms an isolated triradius. One more groove of the third order, 2’’pr, falls into 2’pr in its terminal segment. The 1pr groove is reduced, and its distal segment is outlined by 1’pr. The latter appears as a fovea isolated from fissures III and IV by a narrow enamel bridge. The bridge connects the distal segment of the protocone with the distal portion of the axial ridge of the metacone, thus forming a continuous element of the plagiocrista (Fig. 3).
The protocone and paracone are separated by an accessory mesial cusp, which is formed by accessory grooves 2’ра and 2’pr both falling into fissure II. In modern humans, this cusp is usually round in shape, and is formed by the terminal triradius of fissure II strictly at the mesial marginal ridge. In our case, the cusp appears as a segment lying parallel to fissure II and almost reaching the central fovea. The triradius is biased towards the central third of the intertubercular fissure, which is thereby substantially shortened. “Sprigs” of the triradius are similar to the major tubercular grooves. The epicrista is interrupted. The anterior fovea is absent, as is the enamel extension on the vestibular side of the tooth.
Initially, the tooth had three roots: lingual, mesio-buccal, and disto-buccal (Fig. 4). The lingual root, the longest and the most massive, was destroyed during the paleogenetic investigation. It is oval in section, flattened in the bucco-lingual direction, and branches off the buccal roots at a very large angle. The lingual root separates from the distal root at a level of 3.6 mm from the cementoenamel junction, and from the mesial root at the level of 4.1 mm.
The roots of the buccal side separate only in its lower third, 8.2 mm from the cementoenamel junction. Above this level they are connected by a cemental lamina. The mesial root is curved in the middle third. It is flattened in the mesio-distal direction, and its vestibular portion is more massive than the lingual one. The distal root is the least massive and almost flat in section.
Left upper permanent third molar (М3) from the base of the layer 11.4 at the interface with the layer 12, Denisova 8 (Fig. 5). This is an upper left molar of an adult male (Slon et al., 2015), who was slightly older than the Denisova 4 individual. After restoration of the crown, it became possible to assess its contours. It is of oval shape, without an obliquity in the mesio-vestibular portion, but with an expansion in the vestibular portion as compared to the lingual portion (Fig. 6).
The protocone looks fairly massive despite postmortem destruction. The axial ridge of the cusp is very wide, and, together with the axial ridge of the mesial portion of the metacone, it forms a wide ridge, plagiocrista. It was impossible to determine reliably if the plagiocrista was continuous or discrete. Groove 1pr is worn off, while groove 2pr was probably deeper than 1pr. This conclusion is based on the traceability of 2pr despite substantial attrition of the mesial portion of the crown. It segmented the marginal ridge of the protocone and extended to the vestibular surface of the crown (Fig. 7).
The metacone is very massive, and divided into two parts; it is much larger than the paracone and hypocone. One part is composed of the axial portion of the cusp with a reduced mesial segment, the second part comprises the distal segment and distal marginal ridge. Groove 1me is worn off, while 2me, which separates the two portions of the cusp, is of substantial length and depth and is similar to the intertubercular fissures. In the disto-vestibular
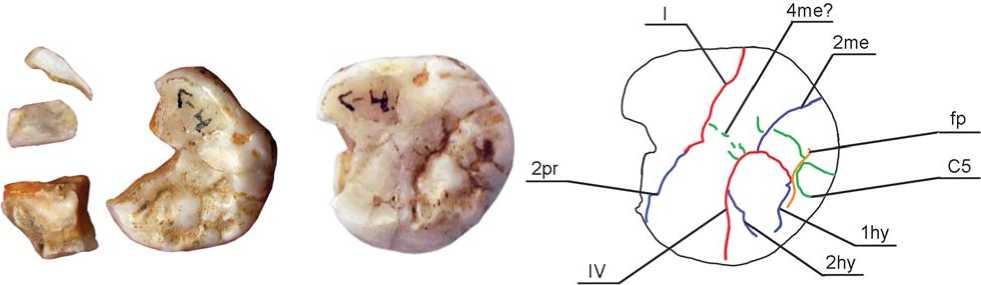
Fig. 5. Denisova 8 molar before the restoration.
Fig. 6. Denisova 8 molar after the restoration.
Fig. 7. Denisova 8: odontoglyphic pattern.
portion of the crown, 2me divides the marginal ridge, and extends well into the external surface of the tooth wall, reaching its lower third. In the central portion, 2me falls into fissure III. The axial ridge of the metacone is as massive in Denisova 8 as in Denisova 4. From the distal side, it is divided by accessory grooves 4me and 4’me, which delineate elements of the metaconulus in its structure (Fig. 7).
A massive ridge is prominent in the cusp that is formed by the distal part of the metacone. This ridge contacts the axial ridge of the hypocone, and forms a structure that is parallel to the plagiogrista but is interrupted by the sagittal groove dividing the metacone and hypocone. The ridge is divided into two parts by an accessory groove falling into 2me on one side, and into the posterior fovea on the other side. The fovea delimits the ridge distally, and separates it from the distal marginal ridge. On the vestibular surface of the metacone, at the interface with the paracone, there is a well-defined mesiostyle. The enamel extension is absent on the vestibular side of the tooth.
The paracone is substantially smaller than the metacone. The mesio-distal diameter of the former visually corresponds to the mesial portion of the latter. Owing to strong attrition and postmortem destruction, it is difficult to describe the paracone. We can only hypothesize the presence of an accessory groove 4ра, which dissected the axial ridge parallel to fissure II.
The hypocone is substantially reduced, and it does not form the disto-lingual corner of the crown. The cusp is smaller than the metacone and protocone and not as prominent as in Denisova 4. According to A. Dahlberg’s scale, which is used in the Zubov protocol, the hypocone can be assigned to grade 4, and according to ASUDAS, to grade 5 (Turner, Nichol, Scott, 1991). The cusp is segmental in shape, with the apex lying at the intersection between fissures III and IV, and it is strongly morphologically differentiated. Three ridges can be observed on its surface: mesial, central, and distal marginal. The distal segment of the major portion of the cusp is reduced. The mesial ridge is very narrow and short, and appears as a thin enamel crest branching out of the margin of the lingual surface in the vestibular direction. It is separated from the axial ridge by the 2hy groove, which merges with intertubercular fissure IV near the lingual margin of the crown (Fig. 7). The axial ridge of the hypocone is outstandingly massive. It lies in the vestibular-lingual direction, parallel to the crista oblique (plagiocrista). It is delimited in distal portion by 1hy, which is almost merged with the posterior fovea separating the axial ridge of the hypocone from its distal marginal ridge. The fovea is very narrow, and appears as a deep groove interrupted by an accessory distal cusp (grades 3–4 ASUDAS). The fissure separating the hypocone and protocone terminates on the surface of the marginal ridge, instead of extending to the lingual surface as is usually found. A thin continuous bridge between the hypocone and protocone remains at the intersection between the occlusal and lingual surfaces of the tooth. It can be interpreted as a rudiment of the archaic ridge called “entocrista”, which is found in primate dentition, but in modern humans is only observed in the deciduous second molar buds (Zubov, Khaldeyeva, 1989).
The root system of the tooth is destroyed; but the morphology of the remaining fragments points towards the presence of three roots: lingual, distal, and mesial.
On the taxonomic status of the dental pattern of the Denisovan upper molars
The permanent molars from Denisova Cave have a number of common traits, which suggests that both specimens belong to the same taxon. The most specific features of their dental pattern are: large size and rounded corners of the crowns; absence of notable reduction of the distal teeth in the row of molars; exceptionally strong development of the grooves of the first and second orders, which usually extend to the walls of the crown or form accessory cusps; and presence of accessory grooves of the third order, which are not found in modern humans. In both teeth, there are cingular structures represented by numerous entostyles in the Carabelli complex, accessory marginal cusps, and mesiostyle of the upper molars. This series of features also includes a pronounced trend towards formation of ridges on the occlusal surfaces of major cusps. This trend is evidenced in the formation of a wide and complex plagiocrista; a segmentation of the metacone accompanied by the emergence of a ridge parallel to the plagiocrista; a segmentation of major cusps; and in the persistence of the entocrista and elements of eocrista.
According to the results of the comparative analysis, the combination of traits observed in Denisovan molars does not match the dental patterns of any European hominin taxa. Most traits typical of Denisovan pattern are highly archaic markers. Despite this, the Denisovan dental complex lacks Neanderthal features (Khaldeyeva, Kharlamova, Zubov, 2010; Bailey, 2002) that might suggest an affinity of the population studied to H. heidelbergensis. The latter taxon is characterized by the increased frequency of archaic markers, accompanied by the presence of Neanderthal traits (Martinón-Torres et al., 2012). Markers of H. sapiens lineage, such as strong reduction of the hypocone of the second upper molar and reduction of the grooves of the third order, are not present in the Denisovan dental pattern. A substantial reduction of the hypocone of the upper third molar is the only relatively advanced evolutionary feature observed in Denisovan teeth. At the moment, there are not enough data to discuss apomorphic features of this species.
The morphological features of the molars from Denisova Cave are generally consistent with the pattern typical of the Archantropus evolutionary stage. They are mostly similar to the dental complexes found in H. erectus of Southeast Asia; in particular, in Sangiran specimens. Sangiran complexes are extremely variable, but there are megadontic specimens among them, comparable in size to Denisovan teeth (Lovejoy, 1970: Tab. 2; Orban-Segebarth, Procureur, 1983: Tab. 2; Tyler, 2001). Almost the full set of archaic markers typical of Denisovans is observed in the Sangiran specimens. First, the frequency of cingular derivatives (ectostyle and ectostylid extensions on the vestibular and lingual surfaces of the upper and lower molars and accessory cusps) is increased in the Sangiran specimens: NG 8503 (Kaifu, Aziz, Baba, 2005), Sangiran 5, 6, 7, 9 (Schwartz, Tattersall, 2003: Vol. II), Sangiran 8 (Kaifu, Aziz, Baba, 2005), Sangiran 22, 27, 33 (Ibid.), and Bpg 2001.4 (Zaim et al., 2011). The trend towards the formation of ridges is in many cases as pronounced in Sangiran hominins as it is in Denisovans. However, it is unclear from published data if the entocrista and eocrista persisted in the specimens from Java, and if they displayed some archaic odontoglyphic variants. For instance, a posterior fovea appearing as a T-shaped fissure was observed in specimen Bpg 2001.4 (Ibid.). Trapezoid contours with rounded corners, similar to those of Denisovans, were described in the upper molars of Sangiran 4, 27, and Bpg 2001.04 specimens (Schwartz, Tattersall, 2003: Vol. II; Zaim et al., 2011).
Another location of dental specimens morphologically similar to the Denisovan teeth is Xujiayao, a final Middle– early Upper Pleistocene site in northern China (Xing et al., 2015). The upper permanent molars from Xujiayao display the following set of features: very large size, subsquare shape of the crown, accessory mesial and distal marginal cusps, complex shape of the Carabelli cusp, differentiated odontoglyphic pattern with extremely pronounced major tubercular grooves, and a tendency towards fragmentation of the metacone and hypocone (Ibid.: Fig. 2). An extremely strong divergence between vestibular and lingual roots, similar to Denisova 4, was observed in the РА 1481 and РА 1500 upper molars (Ibid.: Fig. 3). The dental specimens from Xujiayao display a mosaic morphology, which is substantially different from the samples of early modern humans from China, but retain an archaic component that makes them similar to the dentition of Middle Pleistocene hominins from East Asia: Sangiran, Zhoukoudian, Longtandong, and Chaoxian. From Neanderthal complex markers, only nonspecific traits, broadly found in the samples of the Middle Pleistocene hominins, were observed in the Xujiayao teeth (Ibid.: 237–238).
The taxonomic status of the Xujiayao samples relative to H. sapiens and H. neanderthalensis has not yet been determined. But the strong similarity in morphology of the upper molars from Xujiayao and Denisova Cave might suggest that the two populations could have belonged to the same taxon, exhibiting a long persistence of erectoid traits. If future research confirms this similarity, this will become a strong argument to support the hypothesis that Denisovans were widespread in East Asia (Reich et al., 2010; Derevianko, 2011). It is of note, though, that the complex of archaic features common to H. erectus from Sangiran and Denisovans is substantially reduced, or absent, in other Chinese finds (Turner, Manabe, Hawkey, 2000; Wu, Poirier, 1995; Schwartz, Tattersall, 2003: Vol. II; Xing, Zhou, Liu, 2009; Liu et al., 2010).
The skeletal remains of H. floresiensis (Brown et al., 2004), a species that emerged as a result of long island isolation, confirm the possibility of long-term conservation of erectoid morphology. The dental pattern typical of Denisovans and Xujiayao hominins suggests the presence of one more locus of evolutionary conservation in East Asia.
The similarity between dental complexes of the Upper Paleolithic population from Altai, the Middle Pleistocene hominins from China, and the Lower Paleolithic population from Southeast Asia does not contradict the results of paleogenetic studies. The estimated time of divergence between the ancestors of Denisovans and the common ancestor of H. sapiens and H. neanderthalensis coincides with the latest dates obtained for H. erectus finds from Java (Pope, Cronin, 1984). In Altai, the Karama site has a similar age (Bolikhovskaya, Derevianko, Shunkov, 2006). Thus, a migration becomes a feasible explanation for the similarity between the dental patterns of Denisovans and those of H. erectus from Java. Importantly, the complex of archaic morphological features is more pronounced in later Javanese H. erectus than in earlier specimens (Kaifu et al., 2005).
The greatest genetic impact of Denisovans is found in modern populations from Southeast Asia and Melanesia (Reich et al., 2010), Papua-New Guinea, Polynesia, and Fiji (Reich et al., 2011). The results of these studies have shown that admixture of Denisovan and basal modern human genomes could have occurred in neither the northwest nor the west of the Asian continent. The admixture between these two species most probably took place in Southeast Asia (Ibid.: 523). The paleogenetic data also suggest that Denisovan genes were widespread in this region before the advent of modern humans.
Our results have brought us to the following major conclusions. First, the conservation of archaic components without any replacement by more progressive features was the main evolutionary trend in the emergence of H. altaiensis. The most prominent feature of dental morphology of this species is the set of erectoid traits found in both molars studied. The set is fully present in both Denisovan individuals, despite the high level of genetic divergence between them. Genetic diversity was generally very low in the Denisovan population (Meyer et al., 2012; Slon et al., 2015), which sharply contrasts it to the maximally broad adaptive radiation and genetic diversity typical of modern humans. In this respect, Denisovans were more similar to Neanderthals who, as compared to modern humans, were a more specialized species with a lower level of genetic diversity (Reich et al., 2010: 1055).
Second, the peculiar morphology similar to the Denisovan molars is found only in Asian hominins, but not in any European specimens. Thus, the origin of H. altaiensis is most probably related to Asian H. erectus , which is supported by archaeological data (Derevianko, 2011).
Judging by the prevalence of erectoid features in the morphology of the Denisovan molars, we can hypothesize that the part of Denisovan genome related to an unknown hominin species (Krause et al., 2010) might have belonged to H. erectus. This is just a very tentative suggestion, since most of the archaic traits in the Sangiran hominins dentition are not apomorphic, but rather inherited from more ancient species of the genus, H. habilis and H. rudolfensis. However, from the point of view of dental morphology data, the presence of some genetic heritage of Asian H. erectus in Denisovans appears fairly well-based.
Conclusions
The results of the present study confirm the high importance of dental traits in detecting interspecific differences in the genus Homo . The results have also confirmed the presence in Altai of a specific hominin population referred to as H. altaiensis, and different from H. sapiens and H. neanderthalensis not only genetically but morphologically as well. Peculiar features of dental morphology of this population are megadontia; and a long-term conservation of the dental markers typical of the Middle Pleistocene hominins of Northern China, and of H. erectus from Sangiran.
Thus, H. altaiensis exhibits a very conservative mode of morphological evolution.
The analysis of the Denisovan upper molar morphology has confirmed the equal validity of genetic and morphological criteria for differentiating hominin species, which has been a matter of hot debate in paleoanthropology. Moreover, this morphological analysis has enabled us to put forward a well-based hypothesis according to which the unidentified portion of the Denisovan genome belongs to H. erectus s.l . Thus, our results emphasize the fact that classical dental studies still retain an independent value not lessened by the advent of molecular genetics methods.
