The origin of the Okunev population, southern Siberia: the evidence of physical anthropology and genetics

Author: Kozintsev A.G.
Journal: Archaeology, Ethnology & Anthropology of Eurasia @journal-aeae-en
Section: Anthropology and paleogenetics
Article in issue: 4 т.48, 2020.
Free access
To test the competing hypotheses as to the origin of the Okunev culture, four male cranial series from Okunev cemeteries in the Minusinsk Basin were compared with 23 other pre-Andronovo series from southern Siberia, and 45 Early and Middle Bronze Age groups from Eastern Europe (24 Yamnaya and 21 Catacomb), using multivariate statistical analysis. While the Afanasyevo admixture in the Okunev population is possible, the hypothesis that the Okunev culture of the Minusinsk Basin originated from the second migration from the Eastern European steppes to southern Siberia in the Early Bronze Age is not supported. It could, however, be applied to people associated with the Okunev-type (Chaa- Khol) culture in Tuva, although these may as well have descended from the Afanasyevans. As concerns the Minusinsk Basin and other regions of southern Siberia except Tuva, the fi ndings agree with the idea of a marked evolutionary conservatism peculiar to the autochthonous populations of that territory, as evidenced by the fact that each of the three Early Bronze Age population clusters—on the Yenisei, in the Altai, and in Baraba—has its own Neolithic ancestors in the same area (this does not concern the Chaa-Khol, the Yelunino, and apparently the Samus populations). The immediate ancestors of the Okunev people can be identifi ed with the Neolithic population of the Krasnoyarsk-Kansk area, and more distant ones with the Upper Paleolithic southern Siberian common ancestors of the Okunev people and the Native Americans. These ancestors are evidenced by both cranial data (indirectly) and genetic data (directly). The la tter suggest that among these common ancestors were the Malta boy and the Afontova Gora II male. The Okunev population, then, is a relic, offeri ng us a unique opportunity to see what the Upper Paleolithic ancestors of the Native Americans may have looked like in their southern Siberian homeland.
Southern Siberia, Okunev culture, Yamnaya culture, Catacomb culture, Afanasyevo culture, Native Americans
Short address: https://sciup.org/145146035
IDR: 145146035 | DOI: 10.17746/1563-0110.2020.48.4.135-145
Text of the article The origin of the Okunev population, southern Siberia: the evidence of physical anthropology and genetics
The origin of the Okunev culture is a highly contentious matter. According to the traditional view, this culture had local Neolithic roots and was “inherently Siberian” (Maksimenkov, 1975: 36–37; Vadetskaya, Leontyev, Maksimenkov, 1980: 26; Sokolova, 2009). As a counter to that, a hypothesis that Okunev origins had been related to a migration of one of the Yamnaya-Catacomb groups from European Russia to southern Siberia was proposed (Lazaretov, 1997; Lazaretov et al., 2012)*. According to absolute dates, in the view of A.V. Polyakov (2017), the immigrants displaced or exterminated their predecessors (Afanasyevans) in less than 100 years. Cultural markers
of the migration, such as burials in catacombs, graves with ledges, placement of bodies on the right side, etc., are found only at the early, Uibat, stage of the Okunev culture, whereas later, at the Chernovaya stage, they disappear (Polyakov, 2020a, b).
The key role in the discussion is played by cranial data, but they are ambiguous. A.V. Gromov, who has authored the most detailed study of Okunev craniology, believed that “Okunev population was a mixture of groups differing in origin” (1997: 308). One of these he associated with the Neolithic population of the Krasnoyarsk-Kansk forest-steppe, which is the closest to Okunev group as a whole; the other with the Yamnaya and Yamnaya-Catacomb groups of Kalmykia. “A certain resemblance between Okunev crania and those of the Yamnaya and Catacomb people of Kalmykia does not provide direct evidence of a genetic affinity between them. However, to all appearances, precisely that physical type was peculiar to a population that was the source of the Caucasoid component in Okunev origins. Obviously, this Caucasoid group was outnumbered by the autochthonous component and gradually dissolved in it, having, however, left its trace, as seen by the Caucasoid tendency of both the Okunev population in toto and its separate groups as compared to contemporaneous autochthonous groups” (Ibid.: 315–316). Trying to elaborate his hypothesis, Gromov paid special attention to brachycranic Yamnaya and Yamnaya-Catacomb groups of Kalmykia, because the earliest of the then available Okunev groups, Tas-Khazaa, deviates from others precisely in that direction. Some heterogeneity is found at the within-group level as well: specifically, three female crania from Chernovaya VIII, and Uibat III and V stand out from others by their Mongoloid appearance.
These conclusions are generally rather vague. Those concerning the between-group level are formulated very cautiously and with an eye on the claims made by archaeologists. At the within-group level, the presence of three Mongoloid females does not support the idea of migration. On the contrary, those having an aberrant appearance should be the few Caucasoid males—the presumed immigrants, which is not the case. The results of the multivariate analysis provide, at best, weak indications of heterogeneity. Also, Gromov’s conclusions are quite discordant with the migrationist theories of archaeologists. Attempting to find a compromise, one arrives at a bizarre scenario: immigrants, who had been numerous enough to banish or destroy the Afanasyevans, eventually dissolved in the autochthonous population, which, therefore, should have been even more numerous. Where and how could that have happened? G.A. Maksimenkov’s idea (1975: 36–37) about the “Bronze Age Reconquista”—the return of Okunevans to their former habitat—is much more understandable, as it requires only two components rather than three.
Nonetheless, in the words of I.P. Lazaretov and A.V. Polyakov (2018: 60), “at present, few people doubt that the Okunev phenomenon resulted from intense migratory processes. A direct indication thereof is provided by physical anthropology. The Caucasoid component in the Okunev population differs from others by marked brachycrany and an unusual occipito-parietal deformation. The same features are found in the Late Yamnaya and Yamnaya-Catacomb population of the northwestern Caspian area” (the claim is supported by references to A.V. Gromov and A.A. Kazarnitsky). Now, stating that all this is what “few people doubt” is definitely misleading. For one, this point of view is disputed by a leading expert in the population history of southern Siberia— T.A. Chikisheva. Noting that Gromov was unable to reveal the tentative Caucasoid component in the Okunev population, she writes: “The Altai-Sayan highlands, at least from the Neolithic onward, can be regarded as the distribution area (or part of it) of an evolutionarily conservative substrate representing the Southern Eurasian formation. It is logical to associate the physical type characterized by a wide face and brachycrany, common among the Late Bronze Age people of southern Siberia, with that formation. It can be suggested that the populations of the Sayan piedmont and of the mountain-steppe basins originated from that substrate (people associated with the Neolithic traditions and the Okunev tribes)” (Chikisheva, 2012: 88, 123). She continues: “The Southern Eurasian formation was a substrate for all the autochthonous populations of the Altai-Sayan region known to date… In this context it has become evident that the impact of migratory impulses on the origin of physical types of the Altai-Sayan population was somewhat overstated” (Ibid.: 180).
Our findings are similar. First, the craniometric analysis has demonstrated that among the southern Siberian Bronze Age groups precisely the Okunev group, unlike others such as Afanasyevo, Andronovo, Karasuk, and Tagar, can be considered ancestral to all or most modern populations of southern and western Siberia. This supports the hypothesis about the stability of the autochthonous substrate represented by the Okunev people and their relatives (Kozintsev, 1976). In his dissertation, Gromov attempted to downplay this conclusion, referring to what he viewed as the Mongoloid admixture in Okunevans, which opposes them to other southern Siberian Bronze Age groups (2002: 16–17). Later, however, it was shown that Okunevans cannot be regarded as Caucasoids with a Mongoloid admixture, and Gromov appears to have agreed with this. Indeed, the integration of data on two independent trait systems (craniometry and cranial nonmetrics) has allowed us to conclude that the role of admixture in western and southern Siberia was relatively minor as compared to a considerable evolutionary conservatism of the autochthonous component. Specifically, the trait combination displayed by Okunevans and the Sopka-2 people was, shown to be markedly plesiomorphic (Kozintsev, Gromov, Moiseyev, 2003; Kozintsev, 2004)*.
In addition, an amazing fact was discovered: the combination of metric and nonmetric cranial traits links Okunevans to Native Americans. This discovery, initially outlined as a summary of a conference paper (Kozintsev, Gromov, Moiseyev, 1995), evoked such skepticism among archaeologists that the editors of both Okunev Collections did not venture to invite us to elaborate on our findings. Such an elaboration, based on a more advanced multivariate approach, was presented in an article published in the USA (Kozintsev, Gromov, Moiseyev, 1999), and was later repeated in Russian using new cranial samples (Kozintsev, Gromov, Moiseyev, 2003; Kozintsev, 2004; Vasilyev et al., 2015: 323–325). Obviously, Okunevans played no part in the peopling of the New World, but they and the Native Americans may have had common Upper Paleolithic ancestors in Siberia.
Okunevans display not only biological but also cultural similarities to certain groups of Native Americans. The parallels between Okunev art and that of Na-Dene Indians, noted by A.N. Lipsky (1969), can be supplemented by a rare type of cranial deformation (the obelionic flattening) evidently caused by cradle-boarding practices. Its similarity to the deformation type seen in Yamnaya and Catacomb crania from Kalmykia was studied in detail (Gromov, 1998), but it has never been noticed that an identical type is found in the New World, specifically in crania of the Pueblo Indians of southwestern USA (Nelson, Madimenos, 2010).
Our conclusions has been fully supported by three independently working teams of geneticists—Danish, headed by E. Willerslev (Allentoft et al., 2015); French, headed by K. Keiser (Hollard et al., 2018); and American, headed by D. Reich (Kim et al., 2018). The effect that this rediscovery, which we had awaited for twenty years, produced in the West was described by O.P. Balanovsky: “Overall, the totality of results described, especially the peculiar status of the Okunev group, is quite consonant with earlier findings by physical anthropologists. This is not only my opinion: in his talk at the 2015 Jena Conference Linguistics, Archaeology, and Genetics, Morten Allentoft quoted a reviewer of his article in Nature. The meaning of the passage was that many conclusions about the genetic relationships outlined in that article had been preceded by those found in Russian publications on physical anthropology—and who could imagine that Russian anthropologists were so shrewd? This appears to be a clear indication that geneticists should carry out such studies in collaboration with colleagues representing older and more experienced disciplines” (Balanovsky, 2015: 312). Maybe, but the reaction of our Russian colleagues— physical anthropologists and archaeologists—is stunned silence, as before.
Okunev genomes are specially examined in a master thesis by Allentoft’s student, the Danish geneticist C.G. Zacho, based on the analysis of DNA in samples taken from 18 Okunev individuals (Zacho, 2016). This study needs to be dealt with in some detail here, the more so because it is not mentioned in the recent Russian summary (Polyakov, 2019). First of all, our conclusion about the affinities between Okunevans and modern Siberian groups has been fully supported: “Okunevo is the ancient group currently known with the closest genomic affinity to present day Siberian populations” (Zacho, 2016: 40). The distinctness of Okunevans on the Siberian background, manifested in their ties with Native Americans, is upheld as well: “The observed combination of ancestry proportions appeared unique. The only individuals that had the same components present, albeit in very different proportions, were the Paleoindians” (Ibid.: 38). Our hypothesis was based on the assumption about common ancestors of Okunevans and Native Americans in Upper Paleolithic Siberia. This assumption has now become a fact. Specifically, genetic ties with Okunevans were detected in a boy who had lived at the Upper Paleolithic site Malta near Irkutsk some 24 ka BP, and in a male from the Upper Paleolithic site Afontova Gora II in Krasnoyarsk, dating to 17 ka BP. Both of them, like Okunevans, reveal affinities with Native Americans (Raghavan et al., 2014; Allentoft et al., 2015).
The idea that the Okunev skeletal sample is a heterogeneous mixture (incidentally, finding little support even in cranial studies) is disproved by genetic analysis. “Both the nuclear PCA and ADMIXTURE analyses indicated a very homogenous gene pool in the Okunevo Culture, in correspondence with the previous genetic study of the Okunevo by Allentoft et al. (2015)” (Zacho, 2016: 38). However, the presence of several genetic components in the Okunev gene pool, indicating past admixture, as in the vast majority of known human groups, is apparent (see (Ibid.: App. 6)): apart from the “Native American” autosomal component proper, whose share is estimated at 4.8 %, there is a Western Eurasian component, as Zacho calls it (61.8 %), and a Siberian component (32.6 %). The proportion of both the latter components is high in Native Americans, in the Malta boy (in his genome, the former component predominates), and in the Ust-Ishim male, dating to ~45 ka BP (in whose genetic makeup both components are nearly equally represented) (Fu et al., 2014). The smallest component of the Okunev gene pool (0.8 %) is typical of Southeast Asians, making one recall the Far Eastern complex that L.A. Sokolova identifies in the Okunev culture (2009: 24).
Therefore, although the component termed Western Eurasian is predominant in the Okunev gene pool, there is no need whatsoever to believe that it was introduced by a Bronze Age migration from Eastern Europe. The sharp difference between the Okunev gene pool and that of the Yamnaya-Afanasyevo population, on the one hand, and the genetic affinities between Okunevans and the Upper Paleolithic Siberians (see above), on the other, suggests that the admixture of various components may have begun many millennia before the formation of the Okunev culture*. The genetic homogeneity of the Okunev sample points in the same direction. “It seems most likely that the Western Eurasian component is from a source that shared ancestry with the Malta individual, which had a substantial West Eurasian ancestry, and that the East Asian ancestry arrived from another source” (Zacho, 2016: 39). Such a source, in Zacho’s view, was the population to which the Ust-Ishim individual belonged.
In later studies, these findings were interpreted in a different way. Recent genetic discoveries call into question the unilinear west vs. east dichotomy (Caucasoid vs. Mongoloid in traditional terms). The actual pattern of differentiation in Eurasia and America proved much more complex. According to a new interpretation, the autosomal gene pool of Okunevans as well as the Botai people, the Yamnaya people of northeastern Kazakhstan, and certain groups of the Baikal area is a mixture of two components— the larger Ancient North Eurasian (ANE) and the smaller Ancient East Asian (AEA) (Damgaard et al., 2018).
The former component, which was recently discovered, is present in the Malta boy, who is genetically close to the Afontova Gora II male (Raghavan et al., 2014), whereas among modern groups, those closest to the Upper Paleolithic individuals are Native Americans, Chukchi, Koryaks, Kets, and Selkups (Flegontov et al., 2016). The proportion of ANE in Native Americans amounts to 30–40 %. Kets could have inherited it from Okunevans in their Altai-Sayan homeland (Ibid.). Cranially, the Okunev (or “Americanoid”) tendency is the most distinct in Khakassians of the Sagay clan, who live in the same territory where Okunevans had lived before them (Kozintsev, 2004). Likewise high (about 50 %) is the proportion of ANE in Caucasoids, including the Yamnaya people, who inherited it from their ancestors—the so-called Eastern Hunter-Gatherers. These are represented by two bone samples dating to the mid-sixth millennium BC—one from the Mesolithic cemetery on the Yuzhny Oleny Island in Karelia, the other from a sub-Neolithic site Lebyazhinka IV in the Middle Volga basin, associated with the Elshanka culture (Haak et al., 2015).
As the geography and chronology of the ANE component show, it is misleading to describe it as Western
Eurasian and associate it solely with ancient Caucasoids. To all appearances, it emerged before the Caucasoid-Mongoloid split. It was absent in Central and Western Europe before the Yamnaya expansion (Flegontov et al., 2016). The observed pattern likely suggests that the remote ancestors of the Yamnaya people had migrated from the east, whereas the Yamnaya-Afanasyevo migrations to the east occurred later. The second largest component of the Okunev gene pool—AEA (it can be described as Mongoloid in traditional terms)—was associated with Early Neolithic (Kitoy) population of the Baikal area.
As to the male genetic legacy of Okunevans, in 14 cases out of 16 (87.5 %) the Y-chromosome subclades belong to the eastern haplogroups Q1 and NO1. The former haplogroup, like the autosomal part of the gene pool, links Okunevans with Native Americans. In two instances (12.5 %) subclades of the western haplogroup R1b were found, possibly indicating Yamnaya and/or Afanasyevo affinities, but present also in their likely ancestor—the Elshanka individual from Lebyazhinka IV (Haak et al., 2015; Damgaard et al., 2018; Hollard et al., 2018). The variant detected in the Malta boy is close to the basal type of the R haplogroup (Raghavan et al., 2014). If the Yamnaya-Afanasyevo admixture (those populations are indistinguishable both genetically and cranially) is present in Okunevans, it can be estimated at ~16 %. This signal is not traceable on the X-chromosome, suggesting that the presumed admixture was male-derived (Damgaard et al., 2018). It could have been received from the Afanasyevans, whose cultural effect on the Okunev culture is beyond doubt (Ibid., Suppl.: 21). We had long ago described Okunevans as “Americanoids” with some Caucasoid admixture (Kozintsev, Gromov, Moiseyev, 1995: 77).
Neither craniometry nor genetics, then, gives any reason to think that the Okunev population emerged owing to a second migration from the Eastern European steppes in the Early Bronze Age. If the Afanasyevo admixture is indeed present, this hypothesis is redundant. However, there is a fact that the migrationists for some reason ignore: a small cranial series from the Chaa-Khol (i.e., Okunev-like) burials at the Aimyrlyg cemetery in Tuva, being strikingly different from the Okunev series of the Minusinsk Basin, is morphologically identical to certain Yamnaya and Catacomb series from Ukraine. We pointed to this fact in several publications (Kozintsev, 2008, 2009), including one specially addressing this issue (Kozintsev, Selezneva, 2015). But archaeologists are apparently as skeptical about these conclusions as about those regarding the affinities between the Minusinsk Okunevans and the Native Americans. All this prompts us to revisit the Okunev problem, the more so because the comparative database has been enlarged manifold over the recent years.
Material and methods
Measurements of four male Okunev series were taken from Gromov’s publication (1997). Neither the composition nor the names of these groups conform to the modern classification. In particular, the name “Uibat group” is used in the geographic sense, since this group includes all Okunev crania from the Uibat River valley, not only those dating to the early (Uibat) stage. In Gromov’s dissertation (2002), the name “Uibat group” is used in the chronological sense, with regard to crania formerly included in the Tas-Khazaa group. However, Lazaretov (2019) recently separated the Tas-Khazaa stage from the Uibat stage, which he now believes to have been even earlier and which, so far, is not represented by craniometric data*. To avoid confusion, I will use the groups and the names that were used in Gromov’s publication (1997).
The comparative database includes craniometric data on male series representing populations culturally related to Okunevans—Karakol, Chaa-Khol, Yelunino, Samus, Ust-Tartas, Odino, Krotovo; the Neolithic group from the Krasnoyarsk-Kansk forest-steppe, the Upper Ob (Ust-Isha and Itkul), and the Baraba forest-steppe, as well as measurements of 24 Yamnaya, 21 Catacomb, and 9 Afanasyevo series. Data on the Baraba groups were taken from T.A. Chikisheva’s book (2012: 36–43, 69–72, 98–101) and her publication co-authored with D.V. Pozdnyakov (Chikisheva, Pozdnyakov, 2019). Sources of information about most other groups are indicated in my previous publication (Kozintsev, 2009). I used corrected data on Afanasyevo series (Solodovnikov, 2009). Measurements of crania from Yamnaya and Catacomb burials in the Stavropol area were taken from the publication by G.P. Romanova (1991); those relating to the Azov-Caspian steppes, from A.A. Kazarnitsky’s monograph (2012: 38, 42–43, 47, 49–50, 58, 69, 77, 81, 91, 103); those relating to the Volgograd Region, from the article by M.A. Balabanova (2016); those relating to the Volga-Ural region, from the book by A.A. Khokhlov (2017: 241–242, 246–253, 267–268); and those relating to Ukraine, from the book by S.I. Kruts (2017: 64–66).
Data on 14 principal craniometric traits were elaborated using the multiple discriminant (canonical) analysis, and the Mahalanobis D2 distances corrected for sample size were computed. The distance matrix was subjected to nonmetric multidimensional scaling and cluster analysis. The minimum spanning tree was
*There are only unpublished nonmetric data, which sharply oppose crania of the Uibat stage not merely from other Okunev groups but also from the Yamnaya and Catacomb series. In the light of these data, the idea of Late Yamnaya and Catacomb migration to Siberia appears implausible (I thank Andrey Gromov for this information).
computed, showing the shortest path between the points in the multivariate space. The software included B.A. Kozintsev’s statistical package and Ø. Hammer’s PAST package (Hammer, 2012)*.
Results
On the plane generated by two axes of nonmetric multidimensional scaling, two large clusters, tentatively called eastern and western, are visible (Fig. 1). The eastern cluster is subdivided into three subclusters: (1) Yeniseian, including the Okunev groups and the Neolithic group from the Krasnoyarsk-Kansk forest-steppe; (2) Altaian, including the Neolithic series from the Upper Ob (Ust-Isha and Itkul), and Karakol; (3) Barabian, including the Neolithic group from the Baraba forest-steppe and seven Bronze Age (mostly pre-Andronovo) series from the same region. The structure of the Yeniseian and Barabian subclusters is rather indistinct. Within the Yeniseian cluster, the Neolithic group is not opposed to Okunev groups, but joins one of them—Verkh-Askiz. Within the Barabian cluster, the Neolithic group is opposed to others, but these are arranged without visible correspondence to cultures or stages. The western cluster consists of two subclusters, one including only two groups—Chaa-Khol and Yelunino, the other comprising 54 Yamnaya, Afanasyevo, and Catacomb groups arranged in a random order, indicating close relationship between those three populations.
Edges of the minimum spanning tree, making up a bridge between the eastern and the western cluster, connect the Odino group from Tartas-1 with Samus, and the latter with the Catacomb series from the Stavropol Region. The connection is due to the intermediacy of Samus. Its characteristics, however, are very inaccurate, because it consists of the few male crania, to which female ones have been added after transforming their parameters into male counterparts, using the coefficients of sexual dimorphism. The unreliability of this method is aggravated by the fact that males and females in such cases can represent two different populations—immigrant and native, respectively. Another group, which may be regarded as potentially intermediate, is Yelunino, which falls into the western cluster; in addition, female crania from that group look markedly more Mongoloid than male ones (Solodovnikov, Tur, 2003).
As to Okunev groups, two of them—Uibat (in the geographic sense) and Tas-Khazaa—are very close, and both are somewhat shifted toward the western cluster (this especially concerns the latter group). The same applies
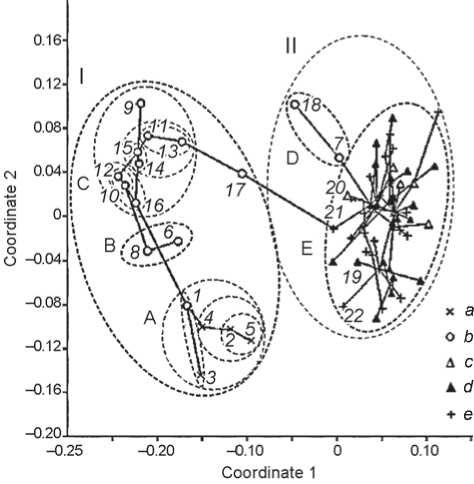
Fig. 1. The position of male cranial series in the space generated by two axes of the nonmetric multidimensional scaling of the corrected Mahalanobis D2 distance matrix.
Straight lines are edges of the minimum spanning tree showing the shortest path between points in the original multivariate space. Dashed contours delineate clusters (I – eastern, II – western) and subclusters (А – Yeniseian, B – Altaian, C – Barabian, D – Chaa-Khol-Yelunino, E – Yamnaya-Catacomb-Afanasyevo).
a – Okunev series; b – other Siberian series except Afanasyevo; c – Afanasyevo; d – Yamnaya; e – Catacomb. 1 – Neolithic group from the Krasnoyarsk-Kansk forest-steppe; 2–5 – Okunev groups: 2 – Uibat (in the geographic sense), 3 – Verkh-Askiz I, 4 – Chernovaya IV, VI, and VIII, 5 – Tas-Khazaa; 6 – Karakol; 7 – Chaa-Khol; 8 – Neolithic of the Upper Ob basin (Ust-Isha and Itkul); 9 – Neolithic of the Baraba forest-steppe; 10 , 11 – Ust-Tartas: 10 – Sopka-2/3, 11 – Sopka-2/3A; 12–14 – Odino: 12 – Sopka-2/4A, 13 – Tartas-1, 14 – Preobrazhenka-6; 15 , 16 – Krotovo: 15 – Sopka-2/4B, C (classic), 16 – Sopka-2/5 (Late Krotovo – Cherno-Ozerye); 17 – Samus; 18 – Yelunino; 19–22 – Yamnaya and Catacomb groups least removed from Okunev and Chaa-Khol: 19 – Yamnaya of the Stavropol Region, 20 – Yamnaya of the Ingulets area, 21 – Catacomb of the Stavropol Region, 22 – Late Catacomb of the Kherson Region.
to the entire Yeniseian subcluster, toward which the Yamnaya-Afanasyevo-Catacomb subcluster, too, shows a slight inclination.
Let us address the ties of separate Okunev groups and of those culturally most related to Okunev. Each of Fig. 2–7 shows ten groups closest to the respective group, ranked in the increasing order of D2values. We will speak of closeness, resemblance, or similarity if D2is less than 5. All distances between Okunev groups meet this condition.
Uibat (in the geographic sense) (Fig. 2). Apart from Okunev groups, it resembles Neolithic groups from the Krasnoyarsk-Kansk forest-steppe and from the Upper Ob (Ust-Isha and Itkul). Three of the five remaining series belong to the eastern cluster, and two to the western cluster.
Verkh-Askiz (Fig. 3). Its closest parallel is the Neolithic series from the Krasnoyarsk-Kansk forest-steppe. None of the other groups, except those associated with Okunev culture, are similar to it. Five of the remaining six groups fall into the eastern cluster, and one into the western cluster.
Chernovaya (Fig. 4). Apart from Okunev series, it resembles Neolithic groups from the Krasnoyarsk-Kansk forest-steppe and from the Upper Ob. Four of the remaining five groups belong to the eastern cluster, and one to the western cluster.
Tas-Khazaa (Fig. 5). Apart from the Okunev groups, it is close only to the Neolithic group from the Krasnoyarsk-Kansk forest-steppe. Among the remaining six series, two belong to the eastern cluster, and four (two Yamnaya and two Catacomb) to the western cluster.
Karakol (Fig. 6). It is similar only to the Neolithic group from the Upper Ob. Three of the remaining nine series are Okunev, and six others are members of the eastern cluster too.
Chaa-Khol (Fig. 7). What we observe here is radically different from anything that we saw before. All ten most similar groups belong to the western cluster, and all are extremely close to Chaa-Khol. The list can be extended, and the use of the reduced trait battery shows that parallels include Western European groups representing the Funnel Beaker population and those associated with the Globular Amphora culture (Kozintsev, Selezneva, 2015). At the same time, there are no indications of especially strong ties between Chaa-Khol and Afanasyevo: the closest groups represent the Yamnaya and Early Catacomb culture of Ukraine, and generally eight of the ten groups most similar to Chaa-Khol belong to Yamnaya and Catacomb populations (17.8 %), whereas one is Afanasyevo (11.1 %). There are even fewer grounds to speak of affinities between Chaa-Khol and the populations of southwestern Central Asia, contrary to what earlier authors believed.
Does any of the Okunev series display real similarity to Yamnaya, Catacomb, or Afanasyevo groups? The question emerges primarily with regard to the Tas-Khazaa group—the earliest available (see Fig. 1, 5). Apart from its similarity to three other Okunev groups, it is close to one more member of the eastern cluster—the Neolithic series from the Krasnoyarsk-Kansk area. If all the first ten groups are considered, then, apart from Okunev groups, there are three of the 12 “eastern” series (every fourth) against four “western” out of 54 members of the Yamnaya-Catacomb-Afanasyevo subcluster (7.4 %), and taking into consideration only the 45 Yamnaya and Catacomb series, 8.9 %, i.e., every tenth at best. The difference is admittedly insignificant, but its direction is opposite to what one might expect according to the hypothesis that Early Okunev males resemble the
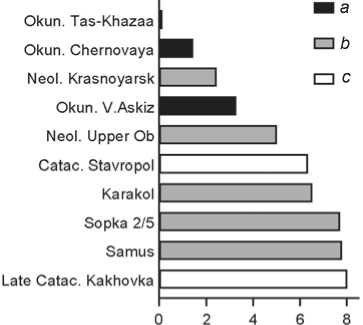
Fig. 2 . Groups closest to Uibat (in the geographic sense) (the actual corrected distance from Tas-Khazaa is negative, i.e. the uncorrected distance is less than its error).
a – Okunev groups; b – other Siberian groups; c – Catacomb groups.
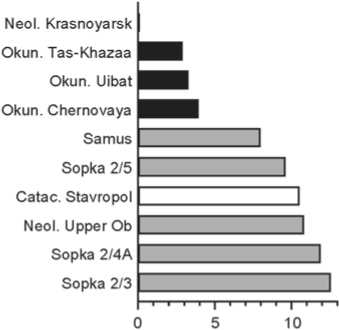
Fig. 3 . Groups closest to Verkh-Askiz (the actual corrected distance from the Neolithic group of the Krasnoyarsk-Kansk forest-steppe is negative).
See Fig. 2 for conventions.
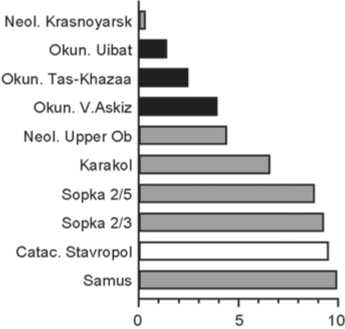
Fig. 4 . Groups closest to Chernovaya.
See Fig. 2 for conventions.
Okun. Uibat
Okun. Chernovaya
Okun. V.Askiz
Neol. Krasnoyarsk
Catac. Stavropol
Late Catac. Kherson
Neol. lipper Ob
Kara koi
Yam. Sravropol
Yam. Kriv. Luka

0 2 4 6 8
Fig. 5 . Groups closest to Tas-Khazaa. See Fig. 2 for conventions.
a – Okunev groups; b – other Siberian groups; c – Yamnaya and Catacomb groups.
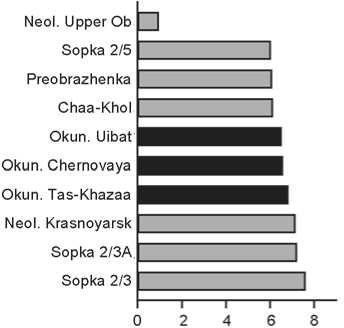
Fig. 6 . Groups closest to Karakol. See Fig. 2 for conventions.

Fig. 7 . Groups closest to Chaa-Khol (Aimyrlyg) (the actual corrected distance from the Yamnaya group of the Ingulets area is negative).
Yamnaya and Catacomb people rather than the Neolithic people of Siberia and their descendants—and this despite the fact that Tas-Khazaa is the earliest and the most “western looking” of the available Okunev groups.
The situation with the Uibat group (in the geographic sense), which also displays a slight “western” tendency, is even clearer (see Fig. 1, 2). Apart from the Okunev groups, it resembles two more “eastern” series, and if more distant ones from the first ten are considered, five “eastern” ones (41.7 %), but only two of the Yamnaya-Catacomb-Afanasyevo cluster (3.7 %). According to Fisher’s exact test, the likelihood that the difference is incidental equals 0.0015, so it can be stated with certainty that Okunevans of the Uibat River valley were cranially closer to Siberian natives than to actual or presumed migrants from the Eastern European steppes. There is no need to discuss two remaining Okunev series—their autochthonous origin is evident and does not require statistical proof. Given these results, based on the entire combination of traits, references to isolated traits such as brachycrany are unconvincing.
Turning to the western cluster, a somewhat specific position of the Catacomb group from the Stavropol Region should be noted, since it seems to display some “eastern” and, respectively, “Okunev” tendency (see Fig. 1–5). This tendency, however, is slight, and there is no resemblance to Okunev groups in the sense outlined above.
Discussion
The findings of this study lend no support to the belief that admixture played a critical role in the origin of the Okunev population. Instead, they agree with the idea that the autochthonous component was predominant and very ancient in that region (Chikisheva, 2012: 88, 123, 180). The key factor affecting the differentiation of native populations falling into the eastern cluster was geographic. Within each of the three eastern subclusters— Yeniseian, Altaian, and Barabian—the Early Bronze Age populations appear to have been directly descended from their local Neolithic predecessors: Okunevans from the Krasnoyarsk-Kansk people; those associated with the Karakol tradition, from people of Ust-Isha and Itkul; and representatives of all cultures and stages at Sopka and their relatives in Baraba, too, had Neolithic ancestors in the same region. Group differentiation must have been caused mostly by random processes, and the effect of migrations was minimal. The search for “racial components” allegedly introduced from without (see, e.g., (Solodovnikov, 2007)) has proved futile in nearly all instances. There are two exceptions— Samus, known from very inaccurate data (see above), and Yelunino.
Okunevans appear to be full-fledged Siberian autochthons, supporting Gromov’s principal conclusion. The Uibat (in the geographic sense) and especially the Tas-Khazaa group display a slight “western” tendency, and the same applies to the whole Yeniseian subcluster as compared to the Altaian and Barabian subclusters. This could well be due to the Yamnaya-Afanasyevo admixture (Damgaard et al., 2018). There are reasons, however, to ascribe this tendency, not so much to the comparatively late (Early Bronze Age) migration from the Eastern European steppes as to much earlier events of the population history.
Indeed, given the affinities of Okunevans with Native Americans (see above), the observed facts are seen in an entirely different light. A number of Native American groups display “quasi-Caucasoid” facial features (Vasilyev et al., 2015: 315–319). Although cranial characteristics of Upper Paleolithic Siberians are unknown, genetic evidence suggests that what we observe in this case is a very ancient legacy. To all appearances, the ANE component, which was abundant in Upper Paleolithic populations of southern Siberia, spread in both directions—eastwards, toward the New World, and westwards, toward Europe. It reached America as early as the Upper Paleolithic, and Eastern Europe no later than the Mesolithic.
The “western” cranial tendency is present in the easternmost of the three eastern subclusters (Yeniseian), whose members, in addition, show an eastern, in fact an “American”, shift in their genetic makeup. It can be concluded that the Afanasyevo admixture, even if present, was not the major factor behind the observed pattern, and it is even less likely that the reason was a second pre-Andronovo migration from the Eastern European steppes, although cultural influences from that territory are quite possible.
A marked craniometric similarity between Okunevans and the Neolithic people of the Krasnoyarsk-Kansk forest-steppe is highly relevant to that issue. Everyone who invokes the migratory factor while discussing Okunev origins must adhere to logic and assume the same with regard to this Neolithic group, and such an assumption is arguably wrong. As concerns pre-Andronovo migrations to southern Siberia in the Early Bronze Age, only one of them is beyond doubt—one that gave rise to Afanasyevo. On that scale, Afanasyevans are virtually indistinguishable from Yamnaya or Catacomb people. If, on the other hand, we postulate a second migration, then its most likely representatives would be the Chaa-Khol people of Tuva. However, they could as well be descendants of Afanasyevans. Being very similar to people of Yamnaya, Catacomb, and Afanasyevo cultures, the Chaa-Khol people still cluster not with them, but with the Yelunino people, whose eastern tendency is accentuated by the fact that Yelunino females are markedly more Mongoloid than males.
Conclusions
-
1. The Okunevans of the Minusinsk Basin should be regarded as southern Siberian autochthons—descendants of the Neolithic and evidently Upper Paleolithic population of that region. Afanasyevo admixture is quite probable, but the hypothesis that the Yamnaya-Catacomb migration had played a considerable role in the origin of the Okunev population is not supported.
-
2. Cranially, the idea of migration is contradicted by a close similarity between Okunevans and the Neolithic population of the Krasnoyarsk-Kansk forest-steppe and by their specifically “Americanoid” tendency.
-
3. Genetically, this idea is disproved by the “Americanoid” characteristics of the Okunev gene pool and by the affinities between Okunevans and the Upper Paleolithic people of southern Siberia—the ancestors of Native Americans.
-
4. If a second pre-Andronovo migration from the Eastern European steppes to southern Siberia took place, then its most likely representatives are people associated with the Okunev-type (Chaa-Khol) culture of Tuva and the Yelunino people.
